 2018, Vol. 38
2018, Vol. 38文章信息
- 杨振冰, 刘园园, 何蕊廷, 杨康, 阿依巧丽, 谢嗣光, 康元昊, 曾波.
- YANG Zhenbing, LIU Yuanyuan, HE Ruiting, YANG Kang, AYI Qiaoli, XIE Siguang, KANG Yuanhao, ZENG Bo.
- 三峡库区不同水文类型支流大型底栖动物对蓄水的响应
- Responses of benthic macroinvertebrates in tributaries of different hydrological regimes in the Three Gorges Reservoir region to reservoir impoundment
- 生态学报. 2018, 38(20): 7231-7241
- Acta Ecologica Sinica. 2018, 38(20): 7231-7241
- http://dx.doi.org/10.5846/stxb201709281753
-
文章历史
- 收稿日期: 2017-09-29
- 网络出版日期: 2018-07-09






河流生态系统中具有多种生物类群, 大型底栖动物(以下简称底栖动物)是河流水生生态系统中的重要生物类群之一, 是河流水生生态系统的重要组成部分, 主要指体长大于500 μm, 生活史的全部或大部分时间生活在水体底部的水生无脊椎动物群[1-2]。底栖动物在水生生态系统的物质循环和能量流动中具有重要作用, 在河流生态系统功能正常发挥和河流健康维持中具有不可替代的地位[3]。不同底栖动物对栖息环境有不同的喜好和要求, 如蜻蜓目中大蜓属和色蟌属喜生活于流水生境中, 在水流速度缓慢的静水生境中却难觅踪影, 大部分蚬类生活在浅水区域而豌豆蚬属适应在深水区域生活。由于生境会直接影响在其中生活的底栖动物, 因此河流中底栖动物的分布和数量可很好地反映河流水生生境现状和河流生境的变化[4-5]。
三峡水库是我国在长江上修建的特大型水库, 水库建成后库区内河流水位抬升、水深增大、流速降低, 库区内河流水文环境发生了重大变化[6]。水库修建不仅对库区内长江干流水文环境产生了巨大影响, 同时对库区内支流的水文环境也产生了很大影响。三峡水库实行冬蓄夏排的调度模式, 夏季维持海拔145米的防洪控制低水位, 水库蓄水影响至重庆涪陵;冬季维持海拔175米的最高蓄水水位, 水库蓄水影响至重庆江津。受三峡水库水位调度模式(图 1)的控制, 三峡库区江津至涪陵段的长江一级支流为周期性受三峡水库蓄水影响的支流, 这些支流的下游河段仅在三峡水库高水位期受水库蓄水影响, 在三峡水库水位位于145米低水位期时不受水库蓄水影响;与此不同, 三峡库区涪陵至大坝段的长江一级支流为长期受三峡水库蓄水影响的支流, 这些支流的下游不仅在三峡水库高水位期受水库蓄水影响, 在水库低水位期同样要受水库蓄水影响。这两种类型支流的下游河段每年受三峡水库影响的程度和受影响的季节不同, 但两种类型支流的上游河段全年均不会被三峡水库蓄水淹没因而不受三峡水库影响。已有不少研究表明, 河流筑坝建库会改变河流的水深、流速、泥沙淤积、河床底质等特征, 对大型底栖动物及水生生态系统造成影响[7-10]。三峡水库成库运行至今已近十年, 目前仍不清楚经过近十年的运行后, 水库修建导致的河流水文条件和河流生境变化是否已对库区河流底栖动物构成和分布产生了影响以及产生了何种影响, 此影响在不同季节和不同水文类型河流间是否表现一致。为能客观真实地反映三峡水库修建对库区支流底栖动物的影响, 最好是在同一时期对受三峡水库影响和不受三峡水库影响的河流的底栖动物状况进行比较研究, 并且被调查河流的底栖动物状况在三峡水库修建前要尽可能相同或相似。根据此原则, 对于库区内长江干流而言, 无论是对库区长江干流底栖动物在建库前和建库后进行比较研究, 还是对不受三峡水库影响的长江干流和受三峡水库影响的长江干流的底栖动物进行比较研究, 均难以清楚说明三峡水库修建对河流底栖动物的影响。因为, 采用前一方法进行比较研究, 建库前后长江干流底栖动物状况即使存在差异, 但此差异既可能是因为水库修建所致, 也可能是因为年份时间不同所致;采用后一种方法进行研究, 不受三峡水库影响的长江干流和受三峡水库影响的长江干流的底栖动物状况即使存在差异, 但此差异也因可能是受三峡水库影响的长江干流段与不受三峡水库影响的长江干流段空间距离相隔太远二者底栖动物基础本底情况不同所导致而难以确定是因为三峡水库修建所致。鉴于此, 在本研究中, 我们在三峡库区选择了水文类型不同但空间位置相距很近, 河流长度相近、流域地理条件和社会经济状况类似的长江一级支流, 对这些支流的底栖动物进行研究。通过对这些不同水文类型支流回水区与非回水区(受三峡水库蓄水影响的河段为回水区, 位于支流下游;不受三峡水库蓄水影响的河段为非回水区, 位于支流上游)的底栖动物数量、密度、生物量和物种多样性以及底栖动物与环境因子关系的研究, 拟探明三峡水库修建对库区河流底栖动物的影响, 为三峡水库修建的生态影响和生态效应研究提供理论依据。

|
| 图 1 三峡水库水位年度变化 Fig. 1 Annual variation of water level in the Three Gorges Reservoir |
本研究在三峡库区选取了水文类型不同但空间位置相距很近、河流长度相近、流域地理条件和社会经济状况类似的四条支流(御临河、梨香江、渠溪河、龙河)开展研究, 其中御临河和梨香江为周期性受三峡水库蓄水影响的支流, 御临河位于长江左岸, 梨香江位于长江右岸;渠溪河和龙河为长期受三峡水库蓄水影响的支流, 渠溪河位于长江左岸, 龙河位于长江右岸。根据河流长度并结合生境特点在四条河流上共布设23个调查样点, 分别是御临河6个样点, 梨香江6个样点, 渠溪河5个样点, 龙河6个样点, 调查样点分布于各支流的回水区和非回水区(图 2)。周期性受蓄水影响支流的回水区调查样点包括御临河的Y4、Y5、Y6和梨香江的LX4、LX5、LX6, 这6个样点在三峡水库高水位蓄水时受蓄水影响, 在三峡水库维持海拔145米的防洪控制低水位期不受影响;非回水区调查样点包括御临河的Y1、Y2、Y3和梨香江的LX1、LX2、LX3, 这六个样点不受三峡水库蓄水影响。长期受蓄水影响支流的回水区调查样点包括渠溪河的Q4、Q5和龙河的L4、L5、L6, 这五个样点全年均受三峡水库蓄水影响;非回水区调查样点包括渠溪河的Q1、Q2、Q3和龙河的L1、L2、L3, 这六个样点不受三峡水库蓄水影响。于2015年7月(即三峡水库低水位期)和2016年1月(即三峡水库高水位期)对上述4条支流各样点开展野外调查、测定和底栖动物研究工作。

|
| 图 2 三峡库区支流御临河、梨香江、渠溪河、龙河大型底栖动物调查样点分布 Fig. 2 Location of sampling sites for benthic macroinvertebrates study in tributaries of Three Gorges Reservoir region, Yulin River, Lixiang River, Quxi River, and Longhe River |
采样过程中针对不同水文条件使用不同工具进行采样, 在水深较浅的非回水区, 底栖动物采集选用面积0.3 m×0.3 m, 网孔径为500μm的改良索伯网, 将索伯网放入河床(网口朝向上游来水方向), 用毛刷将框内石块上的底栖动物刷入网内(框内采样深度约为10 cm);在水深较深的回水区, 选用面积为1/16 m2采泥器进行采样。每个调查样点采集4个重复样, 尽量包括多种微生境, 样品混合后用60目(孔径0.25 mm)分样筛对大型底栖动物进行筛选, 用镊子将底栖动物挑出并装入100 mL标本瓶中, 用5%—8%甲醛固定, 贴标签后带回实验室在体视显微镜下进行鉴定[11-15]和计数, 并使用万分之一天平称量底栖动物生物量(称量前用吸水纸吸干标本表面液体)。
1.2.2 环境因子测定在进行底栖动物采样的同时对各采样点水体的水温、溶解氧、pH、水深、电导率、流速、透明度、总氮、硝态氮、铵态氮、总磷和可溶性磷酸盐进行同步测定。水温、溶解氧、pH、水深和电导率的测定用美国哈希Hydrolab DS5X多功能水质监测仪进行, 流速用LS45A型旋杯式流速仪测定, 水体透明度使用塞氏盘测定, 总氮、硝态氮、铵态氮、总磷和可溶性磷酸盐的测定参考《地表水环境质量标准(GB3838—2002)》[16]。
1.3 数据分析与处理选用Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数描述底栖动物多样性, 各公式如下:
Shannon-Wiener多样性指数[17]:

|
Margalef丰富度指数[18]:
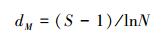
|
Pielou均匀度指数[19]:
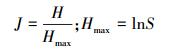
|
式中, S为物种数目, N为样方中观察的个体总数, ni为第i个物种的个数。
使用SPSS 22.0对底栖动物的密度、生物量和各多样性指标进行差异显著性检验, 检验采用单因素方差分析(One-way ANOVA), 方差不齐经转换后仍不齐则用非参数检验(Kruskal-Wallis H检验), 采用Origin9.0作图, 使用Canoco 4.5对底栖动物与环境因子进行相关性分析。
2 结果 2.1 底栖动物物种数量本研究在三峡库区四条支流的两次调查采样过程中共采集到大型底栖动物1778头, 计101种(属), 隶属于4门7纲17目44科, 其中节肢动物种类最多, 达到57种。2015年7月共采集到底栖动物655头59种, 其中在受蓄水影响河段采集到底栖动物4种40头, 优势种为日本沼虾(Macrobrachium nipponensis), 占受蓄水影响河段底栖动物的57.5%;2016年1月采集到底栖动物1123头69种, 其中在受蓄水影响河段采集到底栖动物16种238头, 优势种为锯齿新米虾(Neocaridina denticulata), 占受蓄水影响河段底栖动物的14.2%(表 1)。总体来说, 无论在2015年7月还是在2016年1月, 4条支流的非回水区底栖动物个体数量和物种数量均高于回水区, 此情况在长期受蓄水影响的支流中表现尤为明显。
| 月份 Month |
河流类型 Tributary type |
非回水区 River reaches unaffected by reservoir impoundment |
回水区 River reaches affected by reservoir impoundment |
非回水区优势种 Dominant species in reaches unaffected by reservoir impoundment |
回水区优势种 Dominant species in reaches affected by reservoir impoundment |
| 7 | 周期受蓄水影响支流 | 142头18种 | 101头15种 | 扁蜉Heptagenia 花翅蜉Baetiella |
沼蛤L. lacustris |
| 长期受蓄水影响支流 | 372头52种 | 40头4种 | 扁蜉Heptagenia 河蚬C. fluminea 环棱螺Bellamya 四节蜉Baetis |
日本沼虾M. nipponensis | |
| 1 | 周期受蓄水影响支流 | 200头25种 | 111头14种 | 二翅蜉Cloeon 刺肢米虾C. spinipoda 锯齿新米虾 N. denticulata 小蜉Ephemerella |
斑点小划蝽M. guttata |
| 长期受蓄水影响支流 | 685头51种 | 127头8种 | 椭圆萝卜螺R. swinhoei 四节蜉Baetis 细裳蜉Leptophlebia 蜉蝣Ephemera |
摇蚊Chironomidae 锯齿新米虾 N. denticulata |
在本研究中分别对三峡库区不同水文类型支流的非回水区和回水区底栖动物密度、生物量进行了分析, 结果显示:在2015年7月, 周期性受蓄水影响支流非回水区与回水区的底栖动物平均密度和平均生物量均无差异;长期受蓄水影响支流非回水区与回水区的底栖动物平均密度无显著差异(F=3.920, P=0.076), 但非回水区的底栖动物生物量显著高于回水区的底栖动物生物量(χ2=4.875, P=0.027)(图 3)。在2016年1月, 周期性受蓄水影响支流非回水区与回水区的底栖动物平均密度和平均生物量均无差异;长期受蓄水影响支流非回水区与回水区的底栖动物平均密度无显著差异(χ2=3.692, P=0.055), 但非回水区的底栖动物生物量显著高于回水区的底栖动物生物量(χ2=5.026, P=0.025)(图 4)。

|
| 图 3 三峡水库低水位期(7月)库区不同水文类型支流大型底栖动物密度和生物量 Fig. 3 Benthic macroinvertebrates density and biomass in tributaries of different hydrological regimes in Three Gorges Reservoir region when the reservoir keeping at its low water level (in July) |

|
| 图 4 三峡水库高水位期(1月)库区不同水文类型支流大型底栖动物密度和生物量 Fig. 4 Benthic macroinvertebrates density and biomass in tributaries of different hydrological regimes in Three Gorges Reservoir region when the reservoir keeping at its high water level (in January) |
研究结果表明, 三峡水库成库运行至今, 长期蓄水对河流底栖动物密度没有显著影响, 但对底栖动物生物量产生了显著影响;周期性蓄水对河流底栖动物密度和生物量均无显著影响。
2.3 底栖动物物种多样性对三峡库区不同水文类型支流非回水区与回水区的底栖动物多样性进行研究, 研究结果显示:在2015年7月, 周期性受蓄水影响支流的非回水区与回水区各多样性指标没有显著差异;长期受蓄水影响支流的非回水区和回水区之间部分多样性指标有极显著差异(表 2), 非回水区的Shannon-Wiener指数和Margalef丰富度指数均极显著地高于回水区(P < 0.01), Pielou均匀度指数没有差异(P=0.484)。在2016年1月, 周期性受蓄水影响支流的非回水区与回水区各多样性指标没有显著差异;长期受蓄水影响支流的非回水区与回水区之间部分多样性指标有极显著差异(表 3), 非回水区的Shannon-Wiener指数和Margalef丰富度指数极显著地高于回水区(P < 0.01), Pielou均匀度指数没有差异(P=0.362)。
| 河流类型 Tributary types |
指数 Index |
非回水区 River reaches unaffected by reservoir impoundment |
回水区 River reaches affected by reservoir impoundment |
单因素方差分析 One-way ANOVA |
秩和检验 Kruskal-Wallis Test |
|||
| F | P | χ2 | P | |||||
| 周期性受蓄水影响河流 | Shannon-Wiener指数 | 1.15±0.04 | 0.89±0.18 | 0.837 | 0.360 | |||
| Tributaries periodically affected | Margalef丰富度指数 | 1.42±0.14 | 1.19±0.27 | 0.536 | 0.464 | |||
| by reservoir impoundment | Pielou均匀度指数 | 0.78±0.05 | 0.85±0.07 | 0.094 | 0.766 | |||
| 长期受蓄水影响河流 | Shannon-Wiener指数 | 1.92±0.14 | 0.82±0.14 | 23.799 | 0.001** | |||
| Tributaries continuously affected | Margalef丰富度指数 | 2.69±0.31 | 0.77±0.12 | 17.444 | 0.002** | |||
| by reservoir impoundment | Pielou均匀度指数 | 0.83±0.02 | 0.86±0.03 | 0.528 | 0.484 | |||
| *表示差异显著P < 0.05;**表示差异极显著P < 0.01 | ||||||||
| 河流类型 Tributary types |
指数 Index |
非回水区 River reaches unaffected by reservoir impoundment |
回水区 River reaches affected by reservoir impoundment |
单因素方差分析 One-way ANOVA |
秩和检验 Kruskal-Wallis Test |
|||
| F | P | χ2 | P | |||||
| 周期性受蓄水影响河流 | Shannon-Wiener指数 | 1.06±0.29 | 1.27±0.11 | 0.378 | 0.558 | |||
| Tributaries periodically affected | Margalef丰富度指数 | 1.58±0.46 | 1.82±0.26 | 0.540 | 0.462 | |||
| by reservoir impoundment | Pielou均匀度指数 | 0.67±0.11 | 0.80±0.05 | 0.540 | 0.462 | |||
| 长期受蓄水影响河流 | Shannon-Wiener指数 | 1.74±0.20 | 0.77±0.06 | 8.308 | 0.004** | |||
| Tributaries continuously affected | Margalef丰富度指数 | 2.88±0.42 | 0.66±0.10 | 26.644 | 0.000** | |||
| by reservoir impoundment | Pielou均匀度指数 | 0.69±0.06 | 0.79±0.10 | 0.911 | 0.362 | |||
| *表示差异显著P < 0.05;**表示差异极显著P < 0.01 | ||||||||
研究结果表明, 三峡水库成库运行至今, 长期蓄水对河流底栖动物多样性有极显著影响, 周期性蓄水对河流底栖动物多样性影响不大。
2.4 底栖动物与环境因子的关系对三峡库区4条支流的底栖动物与水体环境因子的关系进行典范对应分析(Canonical Correspondence Analysis, CCA), 分析结果表明, 在2015年7月, 前两排序轴特征值分别是0.715和0.645, 底栖动物物种与环境相关系数分别是0.965和0.980, 相关性较高, 排序结果可靠[20];在2016年1月, 前两排序轴特征值分别是0.522和0.368, 底栖动物物种与环境相关系数分别是0.981和0.947, 相关性较高(表 4)。
| 典范对应分析 Canonical correspondence analysis |
7月July | 1月January | |||||||
| 排序轴1 Axes1 |
排序轴2 Axes2 |
排序轴3 Axes3 |
排序轴4 Axes4 |
排序轴1 Axes1 |
排序轴2 Axes2 |
排序轴3 Axes3 |
排序轴4 Axes4 |
||
| 特征值Eigenvalues | 0.715 | 0.645 | 0.444 | 0.278 | 0.522 | 0.368 | 0.360 | 0.304 | |
| 物种-环境相关性 Species-environment correlations |
0.965 | 0.980 | 0.958 | 0.862 | 0.981 | 0.947 | 0.955 | 0.749 | |
| 物种累积百分比变化率 Cumulative percentage variance of species data |
14.4 | 27.4 | 36.4 | 42.0 | 12.9 | 21.9 | 30.8 | 38.3 | |
| 物种-环境累积百分比变化率 Cumulative percentage variance of species-environment relation |
29.3 | 55.8 | 74.0 | 85.4 | 25.1 | 42.8 | 60.1 | 74.7 | |
在2015年7月三峡水库低水位期, 共筛选出6个对四条支流底栖动物有显著影响的环境因子, 分别是水深(P=0.002)、流速(P=0.002)、硝酸根离子(P=0.002)、溶解氧(P=0.004)、水温(P=0.020)和电导率(P=0.032)。在2016年1月三峡水库高水位期, 共筛选出7个对四条支流底栖动物有显著影响的环境因子, 分别是水温(P=0.016)、溶解氧(P=0.022)、总磷(P=0.002)、流速(P=0.018)、深度(P=0.030)、电导率(P=0.042)和透明度(P=0.042)。
对三峡库区四条支流的底栖动物与水体环境因子的排序分析表明, 在2015年7月三峡水库低水位期, 水体温度和水深(第一排序轴)、水体电导率和溶解氧(第二排序轴)以及硝态氮对底栖动物分布具有影响(图 5), 底栖动物随硝态氮浓度增大而减少, 说明硝态氮浓度越高越不利于底栖动物生存, 这与周然的研究结果相似[21]。在2016年1月三峡水库高水位期, 排序结果表明水体总磷和电导率(第一轴)、水温和溶解氧(第二轴)对底栖动物具有影响(图 5)。上述结果表明, 影响三峡库区河流底栖动物分布的水体环境因子在不同季节有一定的差异。

|
| 图 5 三峡库区支流大型底栖动物与水体主要环境因子典范对应分析排序图 Fig. 5 Correlation plots of canonical correspondence analysis between the benthic macroinvertebrates in tributaries of Three Gorges Reservoir region and water environmental variables CV:流速Current velocity; WT:水温Water temperature; Dep:水深Water depth; SD:水体透明度Water column transparency; Con:电导率Specific conductance; DO:溶解氧含量Dissolved oxygen content; NO3-:硝酸根离子浓度Nitrate concentration; TP:总磷Total phosphorus; 7月: Spc-1.刺肢米虾Caridina spinipoda; Spc-2.日本沼虾Macrobrachium nipponensis; Spc-3.秀丽白虾Palaemon modestus; Spc-4.锯齿新米虾Neocaridina denticulata; Spc-5.扁蜉Heptagenia sp.; Spc-6.细裳蜉Leptophlebia sp.; Spc-7.细蜉Caenis sp.; Spc-8.花翅蜉Baetiella sp.; Spc-9.四节蜉Baetis sp.; Spc-10.宽基蜉Choroterpes sp.; Spc-11.扁蚴蜉Ecdyonurus sp.; Spc-12.似动蜉Cinygmina sp.; Spc-13.黑四节蜉Nigrobaetis sp.; Spc-14.稻象甲Echinocnemus sp.; Spc-15.溪泥甲属Elmis sp.; Spc-16.稻水象甲Lissorhoptrus oryzophilus; Spc-17.水黾Aquarius elongates; Spc-18.纹石蚕Hydropsyche sp.; Spc-19.沼蛤Limnoperna fortune; Spc-20.铜锈环棱螺Bellamya aeruginosa; Spc-21.绘环棱螺Bellamya limnophila; Spc-22.泉膀胱螺Physa fontinalis; Spc-23.椭圆萝卜螺Radix swinhoei; Spc-24.尖膀胱螺Physa acuta; Spc-25.河蚬Corbicula fluminea; Spc-26.方形环棱螺Bellamya quadrata; Spc-27.光滑狭口螺Stenothyra glabra; Spc-28.丝蟌属Lesies sp.; Spc-29.蟌属Coenagrion sp.; Spc-30.近蜓属Periaeschna sp.; Spc-31.虎蜻Epitheca marginata; Spc-32.东方春蜓属Orientogomphus sp.; Spc-33.按蚊Anopheles sp.; Spc-34.摇蚊Chironomidae; Spc-35.新宾纺蚋Simulium xinbinense; Spc-36.大蚊Tiplua sp. 1月: Spc-1.锯齿新米虾Neocaridina denticulata; Spc-2.刺肢米虾Caridina spinipoda; Spc-3.日本沼虾Macrobrachium nipponensis; Spc-4.蜉蝣Ephemera sp.; Spc-5.四节蜉Baetis sp.; Spc-6.黑四节蜉Nigrobaetis sp.; Spc-7.丽翅蜉属Alainites sp.; Spc-8.二翅蜉Cloeon sp.; Spc-9.扁蜉Heptagenia sp.; Spc-10.小蜉Ephemerella sp.; Spc-11.细裳蜉Leptophlebia sp.; Spc-12.沼蛤Limnoperna fortune; Spc-13.河蚬Corbicula fluminea; Spc-14.背角无齿蚌Anodonta woodiana; Spc-15.椭圆萝卜螺Radix swinhoei; Spc-16.泉膀胱螺Physa fontinalis; Spc-17.铜锈环棱螺Bellamya aeruginosa; Spc-18.梨形环棱螺Bellamya purificata; Spc-19.仙女虫Nais sp.; Spc-20.尾鳃蚓Branchiura sp.; Spc-21.斑点小划蝽M. guttata; Spc-22.丝蟌属Lesies sp.; Spc-23.大蜻蜓Anotogaster sieboldii; Spc-24.摇蚊Chironomidae |
水体环境是影响底栖动物分布的主要因素, 段学花对不同河床底质中底栖动物群落结构及多样性进行了研究, 发现底质差异是造成河流底栖动物群落结构不同的关键因素[22], 卵石底质为底栖动物营造了良好的栖息地环境, 底栖动物多样性较高, 淤泥底质和沙质底质空隙小, 导致栖息地多样性低, 底栖动物多样性也相对较低[23-27]。在本研究中发现, 不管是周期受蓄水影响支流还是长期受蓄水影响支流, 两种类型支流的非回水区河床底质均以卵石底质为主, 卵石底质覆盖率达到70%以上, 因此底栖动物多样性均较高;与之成鲜明对比的是, 两种类型支流的回水区由于水流速度缓慢泥沙淤积, 河床底质均以淤泥或沙质为主, 底栖动物多样性不高。
不同底栖动物类群具有不同的生活习性, 这决定了底栖动物主要在何种环境中生活, 例如多数蜉蝣目(Ephemeroptera)昆虫为刮食者, 刮食卵石表面的藻类, 卵石底质多存在于流水生境, 这决定了多数蜉蝣目昆虫主要生活在流水生境中;再如毛翅目(Trichoptera)中的纹石蚕(Hydropsyche sp.)作为收集者主要收集水中有机颗粒, 其防网型巢穴结构决定了只有在流水环境中才能够有效收集有机颗粒, 因此纹石蚕也主要生活在流水生境中。另外, 身体形态也对底栖动物分布起着重要作用, 如呈扁平状的扁蜉(Heptagenia sp.)和呈小鱼状的四节蜉(Baetis sp.), 它们的身体形态使之更适于生活在流水生境中, 这是造成表 1所示结果的原因。
三峡水库长期受蓄水影响支流的回水区受水库影响很大, 自水库建成运行后一直处于静水状态且水深增大, 水文环境发生巨大改变是造成其与非回水区的底栖动物群落结构发生差异的主要原因。长期受蓄水影响支流的回水区内成库前原有适应流水生境的底栖动物大量减少并迁移到其他适宜的生境中, 如喜好栖息于浅滩沙质生境的软体动物会逐渐减少, 迁移到非回水区浅滩生境[28]。支流的回水区与非回水区各自形成稳定的底栖动物群落结构, 回水区主要由适应静水生境的底栖动物类群构成, 非回水区主要由适应流水生境的底栖动物类群构成。在本研究中, 适应静水生境的底栖动物与适应流水生境的底栖动物个体质量的差异是造成三峡水库长期受蓄水影响支流的回水区与非回水区底栖动物生物量差异的原因, 回水区以虾类为主, 虾类个体质量在0.0672—0.2687 g之间, 非回水区有较多河蚬和环棱螺等软体动物, 河蚬的个体质量在1.3141—1.8431 g之间, 环棱螺个体质量在0.6131—2.4430 g之间, 河蚬和环棱螺个体质量显著高于虾类。李金国等人的研究也指出急流区(非回水区)底栖动物的数量和生物量要多于稳水区(回水区)[29]。
三峡水库周期性受蓄水影响支流的回水区受三峡水库影响程度相对较小可能是造成其与非回水区底栖动物群落结构无差异的原因。每年7月, 三峡水库维持145 m低水位运行, 此时周期性受蓄水影响支流回水区不受三峡水库蓄水影响, 河流流态为自然流动水体, 与非回水区水体流态相同, 因此回水区底栖动物状况与非回水区没有显著差异。每年1月, 三峡水库维持175 m高水位运行, 此时周期性受蓄水影响支流的回水区受三峡水库蓄水影响, 水深加深、流速变缓, 悬浊物大量沉积, 为摇蚊幼虫生存带来了良好条件[30]。摇蚊幼虫拥有大约100个属, 每个属有至少一个种, 这决定了即使在1月份三峡水库高水位时期, 周期性受蓄水影响支流的回水区底栖动物物种多样性也不会显著降低, 因此该时期回水区与非回水区底栖动物多样性没有显著差异。
本研究未对两个时期底栖动物进行直接比较, 主要是考虑到不同底栖动物生活史不同, 底栖动物存在形式会随时间变化发生改变, 如摇蚊羽化、蜻蜓变态发育等, 这会对底栖动物群落结构造成一定影响, 不过仍可以从结果中看出1月份底栖动物密度和生物量较7月份高。蒋万祥等人[31]对香溪河的研究以及徐小雨[32]等人对菜子湖的调查均表明底栖动物密度和生物量在冬季最大。
3.2 底栖动物与环境因子的关系环境因子是影响底栖动物群落结构的直接因素[4]。影响底栖动物分布的环境因子非常复杂, 水深是影响底栖动物群落结构的主要环境因子之一, 本研究表明, 7月份水深是影响底栖动物分布的最主要因素, 水深大的地方底栖动物多样性低, 原因是在水深大的地方河床底部光照不足, 影响底栖动物生长。环境温度也是影响底栖动物正常生长发育及繁殖的因素之一[33-35]。本研究图 5显示, 水温是影响底栖动物分布的主要环境因子之一, 一方面温度可以直接影响底栖动物的生长繁殖, 另一方面温度还会影响沿岸植物的凋落过程, 进而间接影响底栖动物的食物来源。此外, 蓄水导致河流状态最直接的变化是水流速度变缓, 流速变缓导致泥沙淤积, 河床底质发生改变, 着生藻类和水草不能很好的附着生长, 底栖动物缺少必要的食物来源和良好的栖息环境, 多样性自然会降低。
总之, 影响底栖动物分布的环境因子非常复杂, 并且随着季节的变化而变化。除了底栖动物群落结构本身随着时间变化而变化之外, 三峡水库也随时间变化周期性的蓄排水, 这使得影响底栖动物分布的环境因子更加复杂多变。
本研究通过对三峡库区两种类型支流底栖动物的研究得出以下结论:三峡水库形成及运行对长期受蓄水影响的支流底栖动物群落结构及其多样性有显著影响;与长期受三峡水库蓄水影响的支流相比, 周期性受蓄水影响的支流受三峡水库蓄水影响程度更轻, 底栖动物群落结构在支流回水区与非回水区之间没有显著差异。研究表明, 三峡水库支流的非回水区由于不受三峡水库蓄水影响, 其底栖动物多样性比回水区高, 因此在对三峡库区底栖动物生物多样性研究和资源保护中应对三峡水库支流非回水区的底栖动物加以重视。
| [1] | Higgins R P, Thiel H. Introduction to the Study of Meiofauna. Washington, DC: Smithsonian Institution Press, 1988. |
| [2] | Hutchinson G E, Edmondson Y H. A Treatise on Limnology, Vol. 4:The Zoobenthos. New York: John Wiley and Sons Inc., 1993. |
| [3] | Vanni M J. Nutrient cycling by animals in freshwater ecosystems. Annual Review of Ecology and Systematics, 2002, 33(1): 341–370. DOI:10.1146/annurev.ecolsys.33.010802.150519 |
| [4] | Linke S, Bailey R C, Schwindt J. Temporal variability of stream bioassessments using benthic macroinvertebrates. Freshwater Biology, 1999, 42(3): 575–584. DOI:10.1046/j.1365-2427.1999.00492.x |
| [5] | 王备新.大型底栖无脊椎动物水质生物评价研究[D].南京: 南京农业大学, 2003. |
| [6] | 王丽婧, 席春燕, 郑丙辉. 三峡库区流域水环境保护分区. 应用生态学报, 2011, 22(4): 1039–1044. |
| [7] | Jorcin A, Nogueira M G. Temporal and spatial patterns based on sediment and sediment-water interface characteristics along a cascade of reservoirs (Paranapanema River, south-east Brazil). Lakes & Reservoirs:Research and Management, 2005, 10(1): 1–12. |
| [8] | Callisto M, Goulart M, Barbosa F A R, Rocha O. Biodiversity assessment of benthic macroinvertebrates along a reservoir cascade in the lower São Francisco river (northeastern Brazil). Brazilian Journal of Biology, 2005, 65(2): 229–240. DOI:10.1590/S1519-69842005000200006 |
| [9] | Downes B J, Lake P S, Schreiber E S G, Glaister A. Habitat structure, resources and diversity:the separate effects of surface roughness and macroalgae on stream invertebrates. Oecologia, 2000, 123(4): 569–581. DOI:10.1007/PL00008862 |
| [10] | Beisel J N, Usseglio-Polatera P, Thomas S, Moreteau J C. Stream community structure in relation to spatial variation:the influence of mesohabitat characteristics. Hydrobiologia, 1998, 389(1/3): 73–88. DOI:10.1023/A:1003519429979 |
| [11] | 蔡立哲. 深圳湾底栖动物生态学. 厦门: 厦门大学出版社, 2015. |
| [12] | Tachet H, Richoux P, Bournaud M, Usseglio-Polatera P.淡水无脊椎动物系统分类、生物及生态学.刘威, 王旭涛, 黄少峰, 译.北京: 中国水利水电出版社, 2015. |
| [13] | 周长发, 苏翠荣, 归鸿. 中国蜉蝣概述. 北京: 科学出版社, 2015. |
| [14] | 辽宁省环境监测实验中心. 辽河流域底栖动物监测图鉴. 北京: 中国环境出版社, 2014. |
| [15] | 大连水产学院. 淡水生物学.上册. 北京: 农业出版社, 1982. |
| [16] | 国家环境保护总局科技标准司. GB 3838-2002地表水环境质量标准. 北京: 中国环境科学出版社, 2002. |
| [17] | Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949. |
| [18] | Margalef R. Information theory in biology. General Systems Yearbook, 1958, 3: 36–71. |
| [19] | Pielou E C. Ecological Diversity. New York: John Wiley and Sons, 1975. |
| [20] | Ter Braak C J F. Canonical correspondence analysis:a new eigenvector technique for multivariate direct gradient analysis. Ecology, 1986, 67(5): 1167–1179. DOI:10.2307/1938672 |
| [21] | 周然, 覃雪波, 彭士涛, 石洪华, 邓仕槐. 渤海湾大型底栖动物调查及与环境因子的相关性. 生态学报, 2014, 34(1): 50–58. |
| [22] | 段学花, 王兆印, 程东升. 典型河床底质组成中底栖动物群落及多样性. 生态学报, 2007, 27(4): 1664–1672. DOI:10.3321/j.issn:1000-0933.2007.04.047 |
| [23] | 段学花, 王兆印, 田世民. 河床底质对大型底栖动物多样性影响的野外试验. 清华大学学报:自然科学版, 2007, 47(9): 1553–1556. |
| [24] | Hawkins C P, Murphy M L, Anderson N H. Effects of canopy, substrate composition, and gradient on the structure of macroinvertebrate communities in Cascade Range streams of Oregon. Ecology, 1982, 63(6): 1840–1856. DOI:10.2307/1940125 |
| [25] | Ormerod S J, Edwards R W. The ordination and classification of macroinvertebrate assemblages in the catchment of the River Wye in relation to environmental factors. Freshwater Biology, 1987, 17(3): 533–546. |
| [26] | Collier K J, Ilcock R J, Meredith A S. Influence of substrate type and physico-chemical conditions on macroinvertebrate faunas and biotic indices of some lowland Waikato, New Zealand, streams. New Zealand Journal of Marine and Freshwater Research, 1998, 32(1): 1–19. DOI:10.1080/00288330.1998.9516802 |
| [27] | Buss D F, Baptista D F, Nessimian J L, Egler M. Substrate specificity, environmental degradation and disturbance structuring macroinvertebrate assemblages in neotropical streams. Hydrobiologia, 2004, 518(1/3): 179–188. |
| [28] | 邵美玲, 谢志才, 叶麟, 蔡庆华. 三峡水库蓄水后香溪河库湾底栖动物群落结构的变化. 水生生物学报, 2006, 30(1): 64–69. DOI:10.3321/j.issn:1000-3207.2006.01.012 |
| [29] | 李金国, 王庆成, 严善春, 姚琴, 乔树亮, 吕跃东, 韩壮行. 凉水、帽儿山低级溪流中水生昆虫的群落特征及水质生物评价. 生态学报, 2007, 27(12): 5008–5018. DOI:10.3321/j.issn:1000-0933.2007.12.009 |
| [30] | 熊飞, 李文朝, 潘继征. 云南抚仙湖摇蚊幼虫的空间分布及其环境分析. 应用生态学报, 2007, 18(1): 179–184. DOI:10.3321/j.issn:1001-9332.2007.01.031 |
| [31] | 蒋万祥, 蔡庆华, 唐涛, 渠晓东. 香溪河水系大型底栖动物功能摄食类群生态学. 生态学报, 2009, 29(10): 5207–5218. DOI:10.3321/j.issn:1000-0933.2009.10.005 |
| [32] | 徐小雨, 周立志, 朱文中, 许仁鑫, 曹玲亮, 陈锦云, 王勋. 安徽菜子湖大型底栖动物的群落结构特征. 生态学报, 2011, 31(4): 943–953. |
| [33] | Rosenberg D M, Wiens A P, Sæther O A. Responses to crude oil contamination by Cricotopus (Cricotopus) bicinctus and C. (C.) mackenziensis (Diptera:Chironomidae) in the Fort Simpson area, Northwest Territories. Journal of the Fisheries Research Board of Canada, 1977, 34(2): 254–261. DOI:10.1139/f77-037 |
| [34] | 宋明江, 邓华堂, 朱峰跃, 刘绍平, 段辛斌, 陈大庆. 三峡水库175m蓄水后大宁河底栖动物群落结构季节动态. 水生生物学报, 2015, 39(5): 1046–1053. |
| [35] | 熊春晖, 张瑞雷, 季高华, 冯立辉, 王丽卿. 江苏滆湖大型底栖动物群落结构及其与环境因子的关系. 应用生态学报, 2016, 27(3): 927–936. |