 2018, Vol. 38
2018, Vol. 38文章信息
- 王伟, 方青青, 王国强, 李瑞敏, 薛宝林, 王红旗.
- WANG Wei, FANG Qingqing, WANG Guoqiang, LI Ruimin, XUE Baolin, WANG Hongqi.
- 呼伦贝尔草原区CO2源、汇及时空分布模拟研究
- Simulation of CO2 source, sink, and flux temporal and spatial distributions in Hulun Buir grassland
- 生态学报. 2018, 38(20): 7288-7299
- Acta Ecologica Sinica. 2018, 38(20): 7288-7299
- http://dx.doi.org/10.5846/stxb201709091623
-
文章历史
- 收稿日期: 2017-09-09
- 网络出版日期: 2018-07-09






2. 中国地质环境监测院, 北京 100081
2. China Institute of Geo-environmental Monitoring, Beijing 100081, China
我国内蒙古草原是典型的半干旱草原, 占据我国北方的广大面积, 是欧亚大陆草原的重要部分, 也是目前中国最佳的天然牧场之一[1-2], 它为中国北方许多生态系统功能(如碳汇, 水土保持和基因库等)提供了重要生态屏障[3-4]。很多研究表明草原生态系统表现为碳汇[5-7], 但也有研究指出草原生态系统的结构相对单一, 受气候变化和人类活动影响明显, 并不是一个稳定的碳汇[8]。我国北方温带典型草原区位于半干旱气候带, 气候-生态系统过渡带, 对气候变化和人类活动响应很敏感, 其碳固定能力水平在全球陆地生态系统中处于最低水平[9], 其生态系统演变经常受干旱气候胁迫, 这可能导致了其碳积累能力下降[10]。由于气候变化和人类活动对于其土壤和植被的影响, 可能导致了草原生态系统内碳源、汇的改变, 因此充分了解特定的气候条件下半干旱草原生态系统中的CO2源、汇特征十分必要。近年来, 关于典型草原区域二氧化碳通量的研究主要集中在具有观测资料的地区, 对于缺资料草原区的净生态系统交换量(NEE)的研究却比较缺乏。
涡度协方差系统测定CO2通量, 误差相对较小, 目前已经成为国际上直接测定大气和群落CO2通量的标准方法[11], 但是涡度协方差系统仍然是小尺度的观测手段, 难以直接外推到更大尺度。于贵瑞等人[12]提出估算区域内净生态系统二氧化碳交换量(NEE)的最常见的方法是用涡度相关技术获取的实测数据和卫星遥感数据构建区域尺度的相关模型。近年来, 植被光合呼吸作用模型(VPRM)作为一种光能利用率模型在国外被广泛应用于陆地生态系统碳通量模拟研究中[13-15], 但国内关于VPRM模型在草原生态系统为主的区域碳通量模拟的应用却很少被关注。VPRM不仅具有CO2通量模型(例如NASA-CASA、SiB2和VPM)的许多特征并且比早期模型更精细地模拟了地表二氧化碳通量[13]。
本研究采用VPRM模拟缺资料的半干旱草原区的净生态系统二氧化碳交换量(NEE)及分析其时空分布现状, 并结合野外调研来定量分析研究区内大气限制和植被类型对二氧化碳通量的响应, 为更进一步评价近年来半干旱草原区域内的碳循环特征提供一定的科学依据。
1 研究区域自然概况本研究选取位于呼伦贝尔市西南区作为研究区, 地处115°31′—121°02′E、47°19′—50°10′N, 其中呼伦贝尔草原区为研究区的主要组成部分(图 1)。呼伦贝尔草原是世界著名的天然牧场, 被认为是我国北方的重要的生态屏障, 但是相较于锡林郭勒典型草原, 由于其通量资料相对缺乏, 所以其碳循环时空分布鲜少有人研究。近年来随着草原生态日益恶化可能导致了其碳源、汇出现了较大波动, 但是由于该地区缺乏充分观测资料, 区域CO2通量模拟研究相对欠缺。

|
| 图 1 研究区概况示意图 Fig. 1 The schematic map of the study area |
本研究的通量资料源于锡林郭勒典型草原东南部的内蒙古通量站, 锡林郭勒典型草原是亚欧大陆典型草原区最具有代表性的羊草草原。呼伦贝尔草原区与锡林郭勒典型草原区都是温带丛生禾草典型草原(图 1), 同时呼伦贝尔草原与锡林郭勒草原区具有很多相似的草原生境特征(表 1)[16-18]。
| 生境特征 Ecologic niche character |
呼伦贝尔草原 Hulun Buir grassland |
锡林郭勒草原 Xilin Gol grassland |
| 气候类型Climate type | 半干旱温带大陆性气候 | 半干旱温带大陆性气候 |
| 海拔Elevation/m | 650—700 | 800—1200 |
| 多年平均降水量Mean annual precipitation/mm | 240—350 | 200—350 |
| 多年平均气温Mean annual temperature/(℃) | -1—2 | -1—5 |
| 土壤类型Soil type | 栗钙土、黑钙土 | 栗钙土 |
| 代表植物群系Representative plant formation | 大针茅、羊草、克氏针茅 | 大针茅、羊草 |
| 草原类型Grassland type | 典型草原为主, 草甸草原分布其中 | 典型草原为主, 草甸草原分布其中 |
在研究区内共选取了48个点进行采样(图 1)。试验的采样时间是于2016年7月29日至8月7日。在采样点的位置处设置一个1 m×1 m样方, 试验调查了每个样方里面内的所有物种, 记录各物种株高。随机在样方内选取两个0.25 m2的区域, 剪取地表以上的所有植物后装袋, 然后在75℃恒温下烘干至恒重后测量其干重得到其地表生物量并记录。
从国家气象科学数据共享服务平台(http://data.cma.cn/site/index.html)获得了8个气象站点(图 1)的2016年内时间分辨率为3h的气象数据, 包括气压(Pa), 风速(m/s), 气温(℃), 降水(mm)和相对湿度(%)等。从青藏高原科学数据中心(http://ns.itpcas.ac.cn/)获得了2016年内空间分辨率为0.5°×0.5°, 时间分辨率为1 h的空间太阳辐射(W/m2)数据。
2.2 基于EC的通量数据从位于内蒙古东部的内蒙古站(43°32′N, 116°40′E)收集二氧化碳通量数据, 数据包括内蒙古站2004年和2005年内时间分辨率为0.5h的净生态系统二氧化碳交换量(NEE, μmol m-2 s-1)实测数据, 光合有效辐射(PAR, photosynthetically active radiation, μmol m-2 s-1), 大气温度(Ts, ℃)。呼伦贝尔草原与锡林郭勒草原的植被类型分布(主要以禾本科植物为主的典型草原)和气候条件很相近(图 1、表 1)。根据前人研究, 假设生态系统结构特征的较小空间变异性不会导致该模型出现很大的不确定性误差[7], 虽然下垫面类型对于净生态系统二氧化碳交换量的估算有着一定的影响, 但研究区主要为禾本科植物为主的草原生态系统, 其垂向结构和水平结构较为相似, 所以认为位于锡林郭勒草原的内蒙古站实测数据用于VPRM模型的参数率定和验证所得到的模型参数同样适用于呼伦贝尔草原。
2.3 卫星遥感数据及数据处理本研究所采取遥感数据源为2004年至2005年和2016年研究区内, 时间间隔为8 d, 空间分辨率为500 m2的MODIS数据(https://modis.gsfc.nasa.gov/data/dataprod/mod13.php; MODerate Resolution Imaging Spectroradiometer, MODIS, MOD09A1)。利用获取的遥感数据计算EVI(Enhanced Vegetation Index, 增强型植被指数)和LSWI(Land Surface Water Index, 地表水分指数)用于VPRM模型参数率定。
EVI和LSWI的计算公式如下:
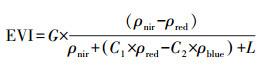
|
(1) |
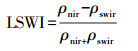
|
(2) |
式中:ρnir表示近红外波段的反射率; ρblue和ρred分别表示蓝色和红色光谱带的反射率; ρswir表示短波红外波段的反射率。G, C1, C2和L是参数, 分别设置为2.5, 6, 7.5和1。
2.4 Vegetation Photosynthesis and Respiration Model (VPRM)模型植被光合呼吸模型(VPRM)是一种基于光能利用效率(LUE)的陆地生态系统诊断模型, 它是在VPM模型[13]的基础上发展而来。同VPM相比, VPRM增加了呼吸项(Reco)及其相关呼吸参数α和β(在接近冻结温度时呼吸基础率), 以及用于反映光合有效辐射与光合作用关系的半饱和值参数PAR0, 虽然VPRM利用遥感数据的间接测量以及其参数验证会给模型带来不确定性, 但模型的参数较少, 易获取, 并可以为研究区提供连续的, 长期的时间序列碳通量模拟数据。以下给出了VPRM模型的简要描述, NEE(净生态系统二氧化碳交换量, Net Ecosystem Exchange)的计算主要包括光照驱动计算的GPP(总初级生产力, Gross Primary Production)和由温度驱动计算的Reco(生态系统植被呼吸通量, Ecosystem Respiration)。
总初级生产力GPP计算过程如下:

|
(3) |
式中, PAR:光合有效辐射, μmol m-2 s-1;FAPARPAV:光合有效辐射吸收比, 即植被吸收的光和有效辐射与光合有效辐射的比值, 并认为EVI与其是线性关系[19], 公式表示为:
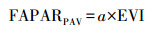
|
(4) |
式中, a是经验参数。
光能利用率(或是光量子效率)由温度, 水分胁迫, 和叶片性状的决定:
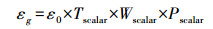
|
(5) |
式中, εg是光能利用率(或是光量子效率), ε0是最大光能利用率(或是最大光量子效率), Tscalar、Wscalar和Pscalar分别代表温度, 水分胁迫, 和叶片性状对光能利用率影响的函数, 其计算公式分别为:

|
(6) |

|
(7) |
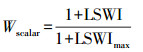
|
(8) |
式中, T代表气温, Tmin、Tmax和Topt分别指光合作用的最小、最大和最适温度, 当气温低于Tmin则Tscalar取值为0。LSWImax是每个站点(或模式格点)生长季内最大的LSWI值。Tscalar、Pscalar和Wscalar值均在[0, 1]范围内。
Mahadevan等人引入呼吸项(Reco)以及相关的呼吸参数α和β到VPM模型中并将呼吸作用简化为温度的线性关系[13], 其公式如下:
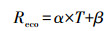
|
(9) |
式中, α和β分别代表与植被类型有关的经验参数, T为气温
根据以上公式:NEE可以通过GPP和Reco进行估算, 公式如下:

|
(10) |
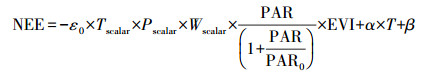
|
(11) |
式中, ε0、PAR0、α和β分别是VPRM模型的四个参数, 通过通量观测数据进行率定。
根据以上介绍, 给出了模型结构如图 2。

|
| 图 2 VPRM模型框架 Fig. 2 The framework of VPRM LSWI:地表水分指数, Land Surface Water Index; EVI:增强型植被指数, Enhanced Vegetation Index; Tscalar, Wscalar和Pscalar:分别代表温度, 水分胁迫, 和叶片性状对植物光能利用率影响的函数, The functions of temperature, water and leaf phenology on light use efficiency of vegetation; T:气温, Temperature; ε0:最大光能利用率, Maximum light use efficiency; α和β:经验参数, Empirical parameter; PAR:光合有效辐射, Photosynthetically Active Radiation; FAPARPAV:光合有效辐射吸收比, The Fraction of Photosynthetically Active Radiation |
由于涡度相关技术观测系统是建立在下垫面条件相似的草原生态系统中, 其不确定性会降低很多;此外, 相关研究表明在内蒙古站羊草草原地区2005年被认为是干旱年[10], 与此类似, 呼伦贝尔草原在2016年也表现为特干旱年。基于相似的生态系统结构及气候特征, 因此我们假设实测数据在研究中, 可适用于VPRM模型的参数的率定和数据验证。研究从2004至2005年的CO2通量实测中有效数据(共1019个)中随机选取了552个数据用于VPRM模型的参数率定, 467个数据用于模型的结果验证。根据图 2所示的计算框架, 通过公式(1)—(11)进行参数率定获得最优的ε0、PAR0、α和β参数。
假设夜间植物不会进行光合作用即GPP为零, 由公式(9)—(10)可知NEE与气温T在VPRM模型中被认为是线性关系, 通过线性回归拟和, 得到α和β。研究拟合了2004年至2005年内时间间隔为0.5 h, 1 d和1个月的NEE和气温(T)时间序列数据, 其中时间间隔为1个月的拟合效果最佳(拟合方程为y=0.0656x+1.1183, R2=0.66, P < 0.01)。其他时间尺度(0.5 h, 1 d和1个月)不存在线性关系, 其原因可能是在短时间尺度上, 环境因子等出现短时间波动的变化影响了植被呼吸作用, 导致NEE没有表现出对于夜间气温的敏感性[20-22]。已知实测数据NEE值与计算得到Reco值, 根据公式(10)可以进一步计算出GPP值, 采用非线性最小二乘法对GPP与ε0和PAR0的方程进行拟合, 确定最优参数ε0和PAR0。从而得到本研究中VPRM模型的所有参数优化结果如下表 2。
| 参数 Parameters |
单位 Unit |
参数值 Value |
| 光能利用率ε0 | — | 0.128 |
| 半饱和值参数PAR0 | μmol m-2 s-1 | 233.69 |
| 经验参数α | μmol CO2 m-2 s-1 C-1 | 0.0656 |
| 经验参数β | μmol CO2 m-2 s-1 | 1.1183 |
| α和β分别代表与植被类型有关的经验参数 | ||
采用VPRM模型模拟出了与实测数据同期的NEE模拟值, 并得到NEE模拟值和实测值的关系拟合图(图 3)。从模拟结果可以看出, 率定期和验证期内的R2分别为0.79(P < 0.01)和0.66(P < 0.01), 具有较好的拟合效果。

|
| 图 3 VPRM模型率定与验证的实测值与模拟值的回归分析 Fig. 3 Regression analysis of measured and simulated values of calibration and validation of VPRM Model |
根据VPRM模型模拟出了2016年呼伦贝尔草原的NEE值, 进一步计算呼伦贝尔草原的NEE空间平均值得到了其年内变化曲线(图 4), 可以看出呼伦贝尔草原的NEE变化趋势是在先增后减(年初和年末水平低, 年中较高), 其变化趋势与当地气温与降水的变化趋势一致。气温作为VPRM模型中的一个输入变量, 也是影响日NEE变化的主要因素之一, 在一定的温度范围内, 温度升高会促使植物的光合作用和呼吸作用速率加快, 但是过高的温度会导致光合作用下降, 碳素转化效率下降[23], 对于草原型地区而言适宜植物光合作用温度为20—30℃之间[24]。降雨会改变区域内的空气湿度和土壤水分, 这是促进植物光合作用、呼吸作用和土壤呼吸作用的重要因素, 其变化趋势与NEE值有着明显相关关系。

|
| 图 4 NEE与气温和降水逐日的2016年内变化曲线图 Fig. 4 NEE with the temperature and precipitation daily changes in the curve in 2016 |
通过图 4可以发现, 呼伦贝尔草原在全年中大部分时期的NEE空间平均值大于零, 其全年NEE值为47.27 g C/m2, 即在2016年内呼伦贝尔草原表现为弱的碳源(与岳泓宇等人[25]的文献统计结果相似)。但也有很多研究认为草原生态系统为主的地区表现为碳汇[5-7]。其实对于草原地区的碳源、汇效应一直有所争议, 其随年气候条件变化而频繁波动, 稳定性较差[8]。
为了进一步分析呼伦贝尔草原的NEE值空间分布特征与植被之间的响应关系, 研究采取在草原生长季(7月1日—8月31日)的NEE平均值(图 5), 分析其空间分布与草原植被类型分布之间的关系。从其分布关系可以看出, 整个区域内NEE值由西南到东北方向依次递减, 根据趋势来看NEE在空间分布由草原区向草甸区和森林区逐渐降低。有研究表明:不同植被类型在一定程度上通过直接和间接的途径影响土壤碳排放。通过图 5中左上角的植被类型分布图[26]可以看出分布于呼伦贝尔草原西南地区的克氏针茅草原和大针茅草原是碳排放最高的区域;中部地区的羊草草原是过渡区, 而东北地区的线叶菊等杂草草甸草原是碳排放最低的区域(与穆少杰等人的研究结果类似[23])。在比较干旱的季节里, 降水事件可能会强烈地激发土壤呼吸[27], 从而使其NEE值大于零, 整个地区都表现为弱的碳源(NEE值较低)。

|
| 图 5 研究区内NEE生长季内平均值空间分布图 Fig. 5 Spatial distribution of NEE values in the study area during the growing season |
气候条件对于生态系统的碳循环有着至关重要的影响, 其中降雨和气温是表征区域气候条件的主要因子, 也是影响植物生理作用的主要环境限制因子[9, 25, 27]。研究根据气象站点资料, 利用呼伦贝尔草原区长时间序列(近50余年)的气温和降水变化(图 6), 分析该地区的2016年的气候状况。

|
| 图 6 呼伦贝尔草原1960—2016年温度及降水时间序列 Fig. 6 The annual precipitation and temperature of Hulun Buir grassland from 1960—2016 |
2016年正是该地区的多年降雨量最小值年, 该年的年平均降雨量(153 mm)远低于该地区50余年的多年平均降雨量(288.85 mm);该年平均气温(-0.205℃)与多年平均气温(-0.22℃)很相近, 但在2016年夏季(植被生长旺盛时期)呼伦贝尔草原内很多地区出现极端高温天气, 短时间局部气温甚至高达44.1℃, 这是该地区有气温纪录以来的历史最高气温[28-30], 这在一定程度上降低了该地区的碳积累。为了进一步分析其干旱程度, 研究采用降水量距平百分率作为评价干旱程度指标。降水量距平百分率能直观反映降水异常引起的干旱, 用降水量与常年同期气候平均降水量之差占常年同期气候平均降水量相比的百分率表示[31], 计算公式如下:

|
(12) |
式中, Pa表示2016年降水量距平百分率, 单位是%;Pr表示2016年的年降雨量, 单位为mm;Pr表示近50年的多年平均降水量, 单位为mm;n表示年数。
采用国家气象干旱等级标准进行评价干旱情况[32], 对于年尺度而言Pa≤-0.45则被认为是“特旱”等级, 由公式12计算可得Pa=-47.03(≤-0.45)。所以可以认为呼伦贝尔草原在2016年表现为特旱年。
结合郝彦宾等人在内蒙古通量站附近的相似研究得到锡林河流域的羊草草原中2005年的年降水量为132 mm, 表现为干旱年, 且在整个生长季生态系统都表现为碳源[10, 31, 33]。类似的, 呼伦贝尔草原在2016年表现为严重干旱年, 也表现为弱的碳源。结合前人相关研究, 原因主要是降水量和气温双重影响, 降水减少以及波动性增加(图 6)会使土壤微生物呼吸上升;温度增加, 促使植物和土壤的呼吸作用增加, 但是光合作用却受到了水分胁迫和高温的影响, 造成了GPP比Reco下降的幅度更大, 这被认为是干旱胁迫降低了草原地区的碳累积[10, 34-35]。通过呼伦贝尔草原的区域年平均气温及年降水时间序列(图 6)可以看出呼伦贝尔草原区的年平均气温有着明显的上升趋势, 年平均降水量有略微的下降趋势, 这样的气候变化可能对近年来半干旱草原区的CO2循环造成了剧烈的影响, 这可能暗示着近年来区域气候变化可能会导致位于半干旱区的呼伦贝尔草原出现碳汇功能正在丧失的趋势。
为了进一步探讨草原碳循环与降雨之间的关系, 本研究通过对线性、指数、对数、二次函数和幂函数分别进行模拟月尺度的降雨和NEE的相关关系, 最后得出二次函数的拟合效果最优。很多研究表明:降雨是限制半干旱草原区固碳能力主要限制因子之一[10, 36-37], 可以看出降水量与NEE在日尺度上并没有表现出明显的相关性(图 7), 但是降水在月尺度上对呼伦贝尔草原内的NEE有极显著的正相关关系(R2=0.938, P < 0.001), 有着良好的拟合效果(图 7), 这可能仍与小时间尺度上气候环境因子变化剧烈有关[20-22]。

|
| 图 7 NEE与日降水量的关系图和NEE与月降水量的关系图 Fig. 7 Fitting graph of NEE and Daily Precipitation and NEE with Monthly Precipitation |
结果表明:在干旱条件下, 在一定的范围内随着降雨增加对于半干旱草原生态系统碳呼吸是先促进后抑制的过程。在干旱的气候条件下草地生态系统中的降雨量增加可能会强烈的激发土壤呼吸, 增加区域内的CO2排放[23, 27, 35], 而随着降雨的增加, 影响了水分在土壤空隙对CO2扩散的阻滞、对微生物活动的刺激等方面可能会抑制土壤呼吸导致其增速会降低[24], 同时在干旱胁迫下, 随着降雨量增加, 植物光合作用强度会逐渐增加, 导致NEE月累计值出现下降的趋势[37-39], 这与Ran等人的研究结果类似[7, 40]。
对于草原地区草地生物量大部分集中在地下, 地下生物量对于估算草地生态系统碳储量及其碳固持效应都至关重要, 而且土壤碳储量远远高于植物碳储量[41-42]。不仅如此, 土壤呼吸能占到草原生态系统总呼吸量的75%[43]。半干旱草原生态系统中碳主要存在于土壤和植被中, 而土壤中碳储存量占其碳储量的绝大部分, 土壤呼吸释放的CO2是草原生态系统碳循环的主要贡献之一, 是人类活动影响碳循环的关键生态学过程[22, 41, 44], 土壤碳库损失可能是CO2排放量增加的重要原因之一, 这有可能是由于该区域在干旱条件下降水气温影响下对生态系统CO2排放的促进比对植被光合作用促进效果更加显著。有研究表明, 半干旱草原的土壤呼吸会在7—8月达到了峰值[45], 呼伦贝尔草原在7—8月份内气温和降水量都处于全年最高水平, 所以气温和降水量增加可能是促进土壤呼吸的重要因素, 这一定程度上是使草原生态系统由碳汇变为碳源的重要原因。前文提到草原地区的碳源、汇效应是受环境因子影响波动较大, 所以研究区在气候干旱, 高温少雨的特殊气候条件下, 在年时间尺度上表现为微弱的碳源, 这一方面可能是草原生态系统的碳循环受气候因子的影响出现了波动, 另一方面通过图 6可知呼伦贝尔草原气温升高趋势十分明显, 而降水存在下降趋势, 这也暗示了近年来我国北方典型草原可能受到了气候变化的影响, 其碳汇功能可能正在丧失。
3.4 生物量与CO2通量的关系分析对于草原生态系统地上生物量与地下生物量、土壤有机质含量以及土壤结构都有着密切的联系, 地上生物量的增加能够提高生态系统的生产力, 增加碳的吸收;但是同时也会使土壤中有机质含量、根系量增加, 促进生态系统碳呼吸增加。为了进一步研究地上生物量与碳源汇的关系, 通过对采样期内(共10 d)的所测得地上生物量(AGB)分别与同时期内在该采样点位置的NEE模拟值进行相关分析, 地上生物量(AGB)与GPP和Reco是正相关的对数函数关系, 与NEE则是负相关的线性函数关系(图 8)。

|
| 图 8 AGB与GPP和Reco的关系图; AGB与NEE的关系图 Fig. 8 Fitting graph of AGB and GPP and Reco; Fitting graph of AGB and NEE |
GPP和Reco与地上生物量(AGB)的拟合效果良好, 决定系数R2分别达到了0.89和0.9(图 8)。许多草原研究提出了GPP或Reco和AGB之间的正相关性[20, 46-48]。研究最后采用对数方程表征这一相关关系。进一步说明随着地上生物量的增长, 到达一定阈值后, 受环境因子的限制其GPP和Reco的增速会有所下降。AGB与NEE具有明显的负相关性(图 8), 这与王忠美[49]等人的研究一致。AGB与NEE表现出了极显著的负相关关系(R2=0.68, P < 0.01), 说明了地上生物量是草原生态系统CO2循环中十分重要的影响因子, 而增加地上生物量能有效的抑制二氧化碳排放。这也侧面说明了由于气候变化的影响可能导致了草原生态系统中地上生物量的减少, 打破了生态系统光合作用与呼吸作用产生的CO2收支平衡。通过图 7可以进一步分析, 在整个呼伦贝尔草原内在2016年的干旱气候条件的情况下, 当每平方米的地表生物量达到152 g/m2左右, 该地区的地表二氧化碳交换将达到收支平衡(NEE为零)。
3.5 结论通过对位于半干旱草原区的呼伦贝尔草原2016年内的NEE空间平均值进行模拟发现, NEE的全年累计值为47.27 g C/m2, 表明2016年该区域表现为微弱的碳源。在空间分布上, 与植被类型关系密切, 克氏针茅草原和大针茅草原是碳排放最高的区域;而线叶菊等杂草草甸草原是碳排放最低的区域。
在干旱条件下, 夏季高温少雨产生的干旱胁迫是使该地区在2016年成为了碳源的主要原因之一。降水量在月尺度上, 在小范围内降水的增加会使该区域CO2通量进一步增加, 但是达到一定阈值后CO2通量出现降低的趋势, 趋于向碳汇的方向发展。
地上生物量与GPP和Reco表现出了极显著的正相关关系, 与NEE表现出了极显著的负相关关系。说明了地上生物量增加能有效的降低二氧化碳排放, 而且预测了地上生物量达到152 g/m2时, 整个地区在2016年的气候条件下二氧化碳交换达到收支平衡。
| [1] | 马秀枝.开垦和放牧对内蒙古草原土壤碳库和温室气体通量的影响[D].北京: 中国科学院研究生院(植物研究所), 2006. |
| [2] | Committee on Scholarly Communication with the People's Republic of China (U.S.). Grasslands and Grassland Sciences in Northern China. Washington, DC: National Academy Press, 1992. |
| [3] | Xu H G, Tang X P, Liu J Y, Hui D, Wu J, Ming Z, Yang Q W, Lei C, Zhao H J, Yan L. China's progress toward the significant reduction of the rate of biodiversity loss. BioScience, 2009, 59(10): 843–852. DOI:10.1525/bio.2009.59.10.6 |
| [4] | Wu J G, Zhang Q, Li A, Liang C Z. Historical landscape dynamics of Inner Mongolia:patterns, drivers, and impacts. Landscape Ecology, 2015, 30(9): 1579–1598. DOI:10.1007/s10980-015-0209-1 |
| [5] | Zhang L, Sun R, Xu Z W, Qiao C, Jiang G Q. Diurnal and seasonal variations in carbon dioxide exchange in ecosystems in the Zhangye Oasis Area, Northwest China. PLoS One, 2015, 10(3): e0120660. DOI:10.1371/journal.pone.0120660 |
| [6] | Han G X, Xing Q H, Yu J B, Luo Y Q, Li D J, Yang L Q, Wang G M, Mao P L, Xie B H, Mikle N. Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta. Agriculture, Ecosystems & Environment, 2014, 196(1793): 187–198. |
| [7] | Ran Y H, Li X, Sun R, Kljun N, Zhang L, Wang X F, Zhu G F. Spatial representativeness and uncertainty of eddy covariance carbon flux measurements for upscaling net ecosystem productivity to the grid scale. Agricultural and Forest Meteorology, 2016, 230-231: 114–127. DOI:10.1016/j.agrformet.2016.05.008 |
| [8] | 耿绍波, 鲁绍伟, 饶良懿, 杨晓菲, 高东, 冯宗红. 基于涡度相关技术测算地表碳通量研究进展. 世界林业研究, 2010, 23(3): 24–28. |
| [9] | 赵娜, 邵新庆, 吕进英, 王堃. 草地生态系统碳汇浅析. 草原与草坪, 2011, 31(6): 75–82. DOI:10.3969/j.issn.1009-5500.2011.06.019 |
| [10] | 郝彦宾, 王艳芬, 崔骁勇. 干旱胁迫降低了内蒙古羊草草原的碳累积. 植物生态学报, 2010, 34(8): 898–906. DOI:10.3773/j.issn.1005-264x.2010.08.002 |
| [11] | Baldocchi D, Valentini R, Running S, Oechel W, Dahlman R. Strategies for measuring and modelling carbon dioxide and water vapour fluxes over terrestrial ecosystems. Global Change Biology, 1996, 2(3): 159–168. DOI:10.1111/gcb.1996.2.issue-3 |
| [12] | 于贵瑞, 孙晓敏. 中国陆地生态系统碳通量观测技术及时空变化特征. 北京: 科学出版社, 2008. |
| [13] | Mahadevan P, Wofsy S C, Matross D M, Xiao X M, Dunn A L, Lin J C, Gerbig C, Munger J W, Chow V Y, Gottlieb E W. A satellite-based biosphere parameterization for net ecosystem CO2 exchange:vegetation photosynthesis and respiration model (VPRM). Global Biogeochemical Cycles, 2008, 22(2): GB2005. |
| [14] | Hilton T W, Davis K J, Keller K, Urban N M. Improving North American terrestrial CO2 flux diagnosis using spatial structure in land surface model residuals. Biogeosciences, 2013, 10(7): 4607–4625. DOI:10.5194/bg-10-4607-2013 |
| [15] | Ahmadov R, Gerbig C, Kretschmer R, Koerner S, Neininger B, Dolman A J, Sarrat C. Mesoscalar covariance of transport and CO2 fluxes:evidence from observations and simulations using the WRF-VPRM coupled atmosphere-biosphere model. Journal of Geophysical Research:Atmospheres, 2007, 112(D22): D22107. DOI:10.1029/2007JD008552 |
| [16] | 陈素华, 宫春宁. 内蒙古草原气候特点与草原生态类型区域划分. 气象科技, 2005, 33(4): 340–344. DOI:10.3969/j.issn.1671-6345.2005.04.011 |
| [17] | 郑晓翾, 王瑞东, 靳甜甜, 木丽芬, 刘国华. 呼伦贝尔草原不同草地利用方式下生物多样性与生物量的关系. 生态学报, 2008, 28(11): 5392–5400. DOI:10.3321/j.issn:1000-0933.2008.11.022 |
| [18] | 杨勇, 李兰花, 王保林, 宋向阳, 刘爱军, 王明玖. 基于改进的CASA模型模拟锡林郭勒草原植被净初级生产力. 生态学杂志, 2015, 34(8): 2344–2352. |
| [19] | Monteith J L. Solar radiation and productivity in tropical ecosystems. Journal of Applied Ecology, 1972, 9(3): 747–766. DOI:10.2307/2401901 |
| [20] | Xu L K, Baldocchi D D. Seasonal variation in carbon dioxide exchange over a Mediterranean annual grassland in California. Agricultural and Forest Meteorology, 2004, 123(1/2): 79–96. |
| [21] | Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grünwald T, Havránková K, Ilvesniemi H, Janous D, Knohl A, Laurila T, Lohila A, Loustau D, Matteucci G, Meyers T, Miglietta F, Ourcival J M, Pumpanen J, Rambal S, Rotenberg E, Sanz M, Tenhunen J, Seufert G, Vaccari F, Vesala T, Yakir D, Valentini R. On the separation of net ecosystem exchange into assimilation and ecosystem respiration:review and improved algorithm. Global Change Biology, 2005, 11(9): 1424–1439. DOI:10.1111/gcb.2005.11.issue-9 |
| [22] | Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 1998, 4(2): 217–227. DOI:10.1046/j.1365-2486.1998.00128.x |
| [23] | 穆少杰, 周可新, 陈奕兆, 杨齐, 李建龙. 内蒙古典型草原不同群落净生态系统生产力的动态变化. 生态学杂志, 2014, 33(4): 885–895. |
| [24] | 杜占池, 杨宗贵, 崔骁勇. 草原植物光合生理生态研究. 中国草地, 1999(3): 20–27. |
| [25] | 岳泓宇, 贾志斌, 梅宝玲, 田淑新. 内蒙古草原温室气体交换通量. 生态学报, 2016, 36(24): 7929–7941. |
| [26] | 中国科学院生态环境研究中心.中国陆地生态系统分类结构.[2018-01-22]. http://www.ecosystem.csdb.cn/ecosys/index.jsp |
| [27] | 陈全胜, 李凌浩, 韩兴国, 阎志丹, 董云社. 土壤呼吸对全球温暖化的响应. 地学前缘, 2003, 10(4): 506–506. DOI:10.3321/j.issn:1005-2321.2003.04.040 |
| [28] | 张敬超, 金磊. 内蒙古鄂温克族自治旗2016年夏季异常干旱分析. 畜牧与饲料科学, 2017, 38(4): 58–59. DOI:10.3969/j.issn.1672-5190.2017.04.019 |
| [29] | 吴昊, 越昆, 那晓东. 呼伦贝尔市2016年夏季干旱情况分析. 内蒙古气象, 2016(6): 10–14. |
| [30] | 李学红. 呼伦贝尔2016年气候条件对天然牧草生长季的影响. 农家科技(下旬刊), 2016(11): 299–299. |
| [31] | 孙晓东, 刘桂香, 包玉海, 路艳峰. 基于降水距平百分率的苏尼特草原干旱特征分析. 内蒙古农业大学学报:自然科学版, 2016, 37(2): 47–54. |
| [32] | 中国气象局. GB/T 20481-2006气象干旱等级. 北京: 中国标准出版社, 2006. |
| [33] | 郝彦宾.内蒙古羊草草原碳通量观测及其驱动机制分析[D].北京: 中国科学院研究生院(植物研究所), 2006. |
| [34] | Harper C W, Blair J M, Fay P A, Knapp A K, Carlisle J D. Increased rainfall variability and reduced rainfall amount decreases soil CO2 flux in a grassland ecosystem. Global Change Biology, 2005, 11(2): 322–334. DOI:10.1111/gcb.2005.11.issue-2 |
| [35] | Knapp A K, Fay P A, Blair J M, Collins S L, Smith M D, Carlisle J D, Harper C W, Danner B T, Lett M S, McCarron J K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science, 2002, 298(5601): 2202–2205. DOI:10.1126/science.1076347 |
| [36] | 鲍芳, 周广胜. 中国草原土壤呼吸作用研究进展. 植物生态学报, 2010, 34(6): 713–726. DOI:10.3773/j.issn.1005-264x.2010.06.011 |
| [37] | 程迁, 莫兴国, 王永芬, 林忠辉. 羊草草原碳循环过程的模拟与验证. 自然资源学报, 2010, 25(1): 60–70. |
| [38] | 李明峰, 董云社, 耿元波, 齐玉春. 温带草原生态系统CO2排放对环境因子变化的响应. 中国农业科学, 2004, 37(11): 1722–1727. DOI:10.3321/j.issn:0578-1752.2004.11.025 |
| [39] | Peng S P, Piao S L, Shen Z H, Ciais P, Sun Z Z, Chen S P, Bacour C, Peylin P, Chen A P. Precipitation amount, seasonality and frequency regulate carbon cycling of a semi-arid grassland ecosystem in Inner Mongolia, China:a modeling analysis. Agricultural and Forest Meteorology, 2013, 178-179: 46–55. DOI:10.1016/j.agrformet.2013.02.002 |
| [40] | Neill C, Davidson E A. Soil carbon accumulation or loss following deforestation for pasture in the Brazilian Amazon//Lal R, Kimble J M, Stewart B A, eds. Global Climate Change and Tropical Ecosystems. Boca Raton, FL: CRC Press, 2000: 526-531. |
| [41] | Ni J. Carbon storage in grasslands of China. Journal of Arid Environments, 2002, 50(2): 205–218. DOI:10.1006/jare.2001.0902 |
| [42] | 马安娜, 于贵瑞, 何念鹏, 王秋凤, 彭舜磊. 中国草地植被地上和地下生物量的关系分析. 第四纪研究, 2014, 34(4): 769–776. DOI:10.3969/j.issn.1001-7410.2014.04.09 |
| [43] | Law B E, Thornton P E, Irvine J, Anthoni P M, Van Tuyl S. Carbon storage and fluxes in ponderosa pine forests at different developmental stages. Global Change Biology, 2001, 7(7): 755–777. DOI:10.1046/j.1354-1013.2001.00439.x |
| [44] | Raich J W, Tufekciogul A. Vegetation and soil respiration:correlations and controls. Biogeochemistry, 2000, 48(1): 71–90. DOI:10.1023/A:1006112000616 |
| [45] | 赵巴音那木拉.施肥对荒漠草原土壤呼吸和生态系统CO2通量的影响[D].呼和浩特: 内蒙古农业大学, 2013. |
| [46] | Flanagan L B, Johnson B G. Interacting effects of temperature, soil moisture and plant biomass production on ecosystem respiration in a northern temperate grassland. Agricultural and Forest Meteorology, 2005, 130(3/4): 237–253. |
| [47] | Suyker A E, Verma S B. Year-round observations of the net ecosystem exchange of carbon dioxide in a native tallgrass prairie. Global Change Biology, 2010, 7(3): 279–289. |
| [48] | Kato T, Tang Y H, Gu S, Cui X Y, Hirota M, Du M Y, Li Y N, Zhao X Q, Oikawa T. Carbon dioxide exchange between the atmosphere and an alpine meadow ecosystem on the Qinghai-Tibetan Plateau, China. Agricultural and Forest Meteorology, 2004, 124(1/2): 121–134. |
| [49] | 王忠美.放牧和开垦对温带典型草原CO2通量的影响[D].北京: 中国农业大学, 2016. |