 2018, Vol. 38
2018, Vol. 38文章信息
- 陈斌, 赵家豪, 关庆伟, 薛建辉, 郭英荣.
- CHEN Bin, ZHAO Jiahao, GUAN Qingwei, XUE Jianhui, GUO Yingrong.
- 江西武夷山中亚热带南方铁杉针阔混交林群落组成与结构
- Community composition and structure of a mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest in Jiangxi Wuyishan, China
- 生态学报. 2018, 38(20): 7359-7372
- Acta Ecologica Sinica. 2018, 38(20): 7359-7372
- http://dx.doi.org/10.5846/stxb201709051597
-
文章历史
- 收稿日期: 2017-09-05
- 网络出版日期: 2018-07-09






2. 南京林业大学生物与环境学院, 南京 210037;
3. 江苏省中国科学院植物研究所, 南京 210014;
4. 江西武夷山国家级自然保护区管理局, 铅山 334500
2. College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China;
3. Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing 210014, China;
4. Administrative Bureau of Jiangxi Wuyishan National Nature Reserve, Yanshan 334500, China
环境污染、生境破坏、气候变化等因素综合导致了全球范围内生物多样性的急剧下降[1], 严重削弱了生物多样性的生态、社会、经济服务功能与价值, 使得生物多样性的保护和维持成为全人类关注的焦点[2-3]。对群落的组成和结构等基本特征的研究, 是解析生态系统功能与过程, 理解生物多样性调控机制的重要基础[4-5]。生态过程的时空尺度依赖性决定了通过小面积、短时期、一次性样地调查的传统手段很难反映影响多样性的因素, 无法准确描述物种组成变化的特征, 进而难以揭示物种分布、群落动态和生物多样性维持等的调控机制[6-7]。自1980年巴拿马Barro Colorado Island(BCI)50 hm2热带雨林长期监测样地的建设开始[6-7], 大型固定样地逐渐成为开展森林多样性监测和生态学研究的重要平台[8-9]。
到目前为止, 中国生态学研究者已在我国温带、亚热带和热带地区建立了多种森林植被类型的固定样地[10]。然而亚热带地区已建的样地多属海拔1500 m以下的常绿阔叶林和常绿落叶阔叶混交林[11-17], 针对该气候区内更高海拔处以针叶树为优势种的针叶(阔叶混交)林而设置的固定样地鲜有报道。因此, 在亚热带建设针叶森林类型的固定样地, 是对中国森林多样性监测的有力补充, 将为我国森林植被类型、生物地理分布格局和生物多样性调控机制等方面的研究提供基础数据。
武夷山地处中亚热带, 是我国大陆东南最高山地, 也是生物多样性的热点区域之一[18]。位于江西武夷山国家级自然保护区海拔1500—1900 m范围, 保存有完好的亚热带中山针叶阔叶混交林和山地暖性常绿针叶林等以针叶树为优势种的森林群落类型[19-20]。保护区内的南方铁杉(Tsuga chinensis var. tchekiangensis)的分布面积达1560 hm2, 是这一特有珍贵物种在我国少有的较大天然种群分布区域[21]。参照巴拿马BCI固定样地的设置规范, 于2014年在江西武夷山国家级自然保护区内建立了一块面积为6.4 hm2的南方铁杉针阔混交林动态监测固定样地。本文基于首次木本植物调查的结果, 从物种组成、区系特征、径级结构和空间分布等方面对南方铁杉针阔混交林进行介绍与分析, 并比较其与我国其他原生或典型的亚热带阔叶林和温带针叶(阔叶混交)林的特殊性。
1 材料与方法 1.1 研究区概况江西武夷山国家级自然保护区(117°39′30″—117°55′47″ N, 27°48′11″—28°00′35″ E)位于江西省铅山县南沿、武夷山脉北段西北坡, 总面积16007 hm2。区内地貌形态为强烈侵蚀的岩浆岩中山地貌, 平均海拔1200 m, 最低点300 m, 最高点2160.8 m。该区域属中亚热带季风气候, 年平均气温13.2—14.8℃;月平均气温以一月3.6℃为最低, 七月23.8℃为最高;年均降水量1814—3545 mm, 主要集中在4—6月;年均蒸发量553—941 mm;年均湿度80%—92%[20]。
保护区的主要生态系统类型为中亚热带中山山地森林生态系统。随着海拔的升高, 常见植被类型依次为毛竹林(350—1200 m)、常绿阔叶林(1300 m以下)、常绿落叶阔叶混交林(1300—1800 m)、中山针阔混交林(1500—1900 m)、山地暖性常绿针叶林(1700—1900 m)、山顶苔藓矮林(1700—2000 m)和山顶灌丛草甸(1900 m以上的山顶或山脊)[19-20]。保护区内植物地理区系成分复杂多样, 以温带—亚热带为主;植物种类十分丰富, 共有高等植物292科1126属2829种。国家重点保护野生植物有红豆杉(Taxus chinensis)、香榧(Torreya grandis)、鹅掌楸(Liriodendron chinense)、川黄檗(Phellodendron chinense)、南方铁杉等21种。此外, 保护区也是黑麂(Muntiacus crinifrons)、黄腹角雉(Tragopan caboti)、白颈长尾雉(Syrmaticus ellioti)等多种国家一级重点保护野生动物的栖息地[20]。
1.2 样地设置2014年, 参照森林固定大样地设置技术规范[8], 于江西武夷山国家级自然保护区黄岗山西北坡设置中心点坐标为27°50′26″ N, 117°45′43″ E, 面积为6.4 hm2(400 m×160 m)的中亚热带南方铁杉针阔混交林动态监测固定样地(简称JWY样地)。样地平均海拔1800 m, 最低海拔1758 m, 最高海拔1843 m, 高差约85 m(图 1)。运用全站仪把样地分成160个20 m×20 m的样方, 每个样方进一步细分为16个5 m×5 m的小样方。以样地西北角为原点, 西东向为x轴正方向, 北南向为y轴正方向, 在每个小样方内对所有胸径(DBH)≥1 cm的木本植物进行物种鉴别、坐标定位、胸径测量和个体挂牌。

|
| 图 1 江西武夷山中亚热带南方铁杉针阔混交林样地等高线图 Fig. 1 Contour map of Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
JWY样地内记录的物种名称及分类均以中国植物志[22]为标准。属级和科级植物区系类型的划分分别以吴征镒[23]和吴征镒等[24]为标准。根据中国植物志[22]及实地观测划分林层树种。运用R软件(Version 3.4.0)进行数据分析和制图。其中, 文中所用的统计数据, 除胸高断面积外, 均基于独立个体数;物种重要值= (相对多度+相对频度+相对优势度)/3;种-面积曲线和个体数-面积曲线分析采取随机取样法(随机取样9999次);径级结构分析以2 cm为间隔单位;采用Gini指数和Lorenz asymmetry指数[25]对具体物种的种群径级大小不均等性进行量化分析;对具体物种的种群分布格局采用方差/均值比法、聚块性指标和负二项参数法进行综合分析[26]。
2 结果与分析 2.1 物种组成JWY样地内DBH≥1 cm的木本植物独立个体共有14415株(2252株/hm2), 包括分支的个体共有18805株(2938株/hm2)。样地内物种共有89种, 隶属29科53属, 包括裸子植物3科3属4种, 被子植物26科50属85种。其中, 针叶树种4种, 占总种数4.5%;常绿阔叶树种35种, 占比39.3%;落叶阔叶树种50种, 占比56.2%。常绿阔叶和落叶阔叶树种多度分别为8216和5229, 它们的多度之和达到总多度的93.3%。而针叶树种多度仅为970。常绿阔叶、落叶阔叶和针叶树种的胸高断面积分别为14.15, 5.50和18.23 m2/hm2, 比值为2.6:1:3.3。此外, 常绿阔叶、落叶阔叶和针叶树种的重要值分别为45.1, 34.4和21.5, 比值为2.1:1.6:1。
在科水平上, 样地内个体数最多的是杜鹃花科(图 2), 共有3741株, 占个体数的25.96%;其次为山茶科(2899株, 20.10%)和木兰科(2043株, 14.17%);样地内胸高断面积最大的科为松科(图 2), 达到18.12 m2/hm2, 占整个样地的47.83%;其次为壳斗科(7.95 m2/hm2, 20.97%)和杜鹃花科(3.71 m2/hm2, 9.79%)。

|
| 图 2 江西武夷山中亚热带南方铁杉针阔混交林样地木本植物多度和胸高断面积排名前8位的科 Fig. 2 Top eight woody plant families in the ranks of abundance and basal area at breast height in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
在物种水平上(表 1), 多度超过1000的物种依次为岩柃(Eurya saxicola)、闽皖八角(Illicium minwanense)、鹿角杜鹃(Rhododendron latoucheae)、贵定桤叶树(Clethra cavaleriei)和灯笼树(Enkianthus chinensis), 它们的个体数共占样地总个体数的64.5%。这5个物种中, 除闽皖八角属于大乔木外, 其他4个物种均属于灌木(或小乔木)。此外, 样地内多度大于500的物种还包括南方铁杉和多脉青冈(Cyclobalanopsis multinervis)。这两种树种均属于大乔木, 是样地主林层的主要组成。对物种多度累计分布的分析发现(图 3), 个体数最多的4个物种, 其数量之和已经超过了总个体数的一半。多度排名前28位的物种, 他们的个体数之和达到总个体数的95%。而多度排名后40位的物种, 他们的个体数之和仅为总个体数的1%。根据每公顷个体数“不多于1”和“1到10之间”的稀有种和偶见种定义[7], 样地内共有稀有种32个, 偶见种34个, 分别占物种总数的36.0%和38.2%;而它们的个体数分别占总个体数的0.5%和6.4%。

|
| 图 3 江西武夷山中亚热带南方铁杉针阔混交林样地物种多度的累计分布图 Fig. 3 Cumulative distribution of species abundance in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
从平均胸径上看(表 1), 样地内所有独立个体的胸径平均值为9.25 cm, 最大值为102.7 cm(南方铁杉)。平均胸径超过30 cm的物种为南方铁杉(33.93 cm)和亮叶桦(Betula luminifera, 33.00 cm), 但后者在整个样地中只有1株;平均胸径超过20 cm的其他物种还包括红豆杉(24.17 cm)、多脉青冈(22.67 cm), 色木槭(Acer mono, 22.00 cm)、秀丽槭(Acer elegantulum, 21.49 cm)、黄山松(Pinus taiwanensis, 21.03 cm)、青冈(Cyclobalanopsis glauca, 20.54 cm)和山乌桕(Sapium discolor, 20.07 cm)。其中红豆杉和山乌桕在整个样地中的个体数仅分别为10株和3株。
| 物种名 Species | 科 Family | 多度 Abundance | 平均胸径 Mean DBH/cm | 胸高断面积 Basal area / (m2/hm2) | 重要值 Important value |
| 南方铁杉 Tsuga chinensis var. tchekiangensis | Pinaceae | 894 | 33.93 | 17.7112 | 19.55 |
| 岩柃Eurya saxicola | Theaceae | 2445 | 4.84 | 1.0775 | 8.68 |
| 多脉青冈Cyclobalanopsis multinervis | Fagaceae | 576 | 22.67 | 5.8337 | 8.26 |
| 闽皖八角Illicium minwanense | Magnoliaceae | 2032 | 6.97 | 1.9113 | 8.15 |
| 鹿角杜鹃Rhododendron latoucheae | Ericaceae | 1828 | 5.65 | 1.0742 | 7.09 |
| 贵定桤叶树Clethra cavaleriei | Clethraceae | 1606 | 5.82 | 1.0002 | 6.4 |
| 灯笼树Enkianthus chinensis | Ericaceae | 1382 | 5.65 | 0.9439 | 5.71 |
| 青冈Cyclobalanopsis glauca | Fagaceae | 198 | 20.54 | 1.71 | 2.95 |
| 老鸹铃Styrax hemsleyanus | Styracaceae | 275 | 10.43 | 0.6193 | 2.44 |
| 云锦杜鹃Rhododendron fortunei | Ericaceae | 288 | 11.82 | 0.6685 | 2.33 |
| 猴头杜鹃Rhododendron simiarum | Ericaceae | 199 | 16.16 | 0.9949 | 1.96 |
| 中华石楠Photinia beauverdiana | Rosaceae | 176 | 7.23 | 0.2395 | 1.83 |
| 白檀Symplocos paniculata | Symplocaceae | 245 | 8.08 | 0.3238 | 1.78 |
| 微毛柃Eurya hebeclados | Theaceae | 280 | 6.92 | 0.2341 | 1.72 |
| 叶萼山矾Symplocos phyllocalyx | Symplocaceae | 186 | 5.96 | 0.1143 | 1.42 |
| 秀丽槭Acer elegantulum | Aceraceae | 76 | 21.49 | 0.5019 | 1.31 |
| 红麸杨Rhus punjabensis var. sinica | Anacardiaceae | 121 | 6.39 | 0.0872 | 1.25 |
| 三桠乌药Lindera obtusiloba | Lauraceae | 132 | 8.79 | 0.1649 | 1.17 |
| 红柴枝Meliosma oldhamii | Sabiaceae | 117 | 8.07 | 0.1256 | 1.14 |
| 天目紫茎Stewartia gemmata | Theaceae | 110 | 11.91 | 0.2866 | 1.07 |
| 茶荚蒾Viburnum setigerum | Caprifoliaceae | 95 | 4.11 | 0.0396 | 1.06 |
| 柯Lithocarpus glaber | Fagaceae | 57 | 17.97 | 0.3915 | 1.01 |
| 重要值>1的物种总和 | 13318 | 9.31 | 36.05 | 88 | |
| 全部89种物种总和 | 14415 | 9.25 | 37.89 | 100 |
从胸高断面积上看(表 1), 样地内所有个体数的胸高断面积之和为37.89 m2/hm2, 仅南方铁杉的胸高断面积就达到17.71 m2/hm2, 而多脉青冈的胸高断面积也达到了5.83 m2/hm2。作为样地主林层的主要构成, 这两种树种的胸高断面积之和占样地总胸高断面积的62.13%。此外, 胸高断面积超过1 m2/hm2的物种还包括大乔木型树种闽皖八角(1.91 m2/hm2)和青冈(1.71 m2/hm2), 以及灌木(或小乔木)型的岩柃(1.08 m2/hm2)、鹿角杜鹃(1.07 m2/hm2)和贵定桤叶树(1.00 m2/hm2)。
通过对物种重要值的分析发现(表 1), 样地内重要值>5的物种依次为南方铁杉、岩柃、多脉青冈、闽皖八角、鹿角杜鹃、贵定桤叶树和灯笼树共7种。其中南方铁杉是重要值超过10的唯一物种, 达到19.55。这也充分体现了南方铁杉在样地当前群落结构中的优势地位。重要值>1的物种还包括青冈、老鸹铃(Styrax hemsleyanus)、云锦杜鹃(Rhododendron fortunei)等15个物种。此外, 重要值< 0.1的物种共计38种, 占总物种数的43.7%。
对种-面积曲线的分析发现(图 4), 在取样面积较小的初始阶段, 物种数随面积的增加而迅速增加。当取样面积达到0.8 hm2时, 出现近45个物种, 占样地总物种数的一半。而当取样面积超过0.8 hm2后, 物种数的增速明显放缓并逐渐稳定。通过对物种数(S)与取样面积(A)进行幂函数关系的拟合, 得到方程:
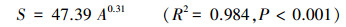
|

|
| 图 4 江西武夷山中亚热带南方铁杉针阔混交林样地的种-面积曲线和个体数-面积曲线 Fig. 4 Species-area and individual-area curves in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
对个体数-面积曲线的分析发现(图 4), 个体数随取样面积的增加而线性增加。通过对个体数(N)与取样面积(A)进行经过原点的线性关系拟合, 得到方程:
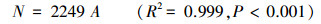
|
由表 2可以看出, JWY样地29科木本植物共划分为7个分布区类型, 53属共划分为8个分布区类型。科水平上的区系组成主要为热带成分, 共计15科, 占总科数的51.7%;其中又以泛热带类型最多, 有9科, 占热带成分总科数的60.0%。科级温带成分共10科, 占总科数的34.5%。与科级区系特征不同的是, 属水平上的区系组成却以温带成分为主, 共计34属, 占总属数的64.2%;其中北温带类型最多, 东亚及北美间断分布类型次之, 两者占温带成分的79.4%。属级热带成分共19个, 占总属数的35.8%。
| 分布区类型 Areal types | 科数 No. of families | 科的比例 Proportion of families /% | 属数 No. of genera | 属的比例 Proportion of genera /% |
| 1.世界广布Widespread | 4 | 13.8 | 0 | 0.0 |
| 2.泛热带Pantropic | 9 | 31.0 | 7 | 13.2 |
| 3.东亚(热带、亚热带)及热带南美间断 Trop. & Subtr. E. Asia & (S.) Trop. Amer. disjuncted | 4 | 13.8 | 5 | 9.4 |
| 4.旧世界热带Old World Tropics | 0 | 0.0 | 1 | 1.9 |
| 6.热带亚洲至热带非洲Trop. Asia to Trop. Africa | 1 | 3.4 | 0 | 0.0 |
| 7.热带亚洲Trop. Asia | 1 | 3.4 | 6 | 11.3 |
| 热带成分(分布型2—4, 6, 7)Tropical | 15 | 51.7 | 19 | 35.8 |
| 8.北温带N. Temp. | 9 | 31.0 | 15 | 28.3 |
| 9.东亚及北美间断E. Asia & N. Amer. disjuncted | 1 | 3.4 | 12 | 22.6 |
| 10.旧世界温带Old World Temp. | 0 | 0.0 | 2 | 3.8 |
| 14.东亚E. Asia | 0 | 0.0 | 5 | 9.4 |
| 温带成分(分布型8, 9, 10, 14)Temperate | 10 | 34.5 | 34 | 64.2 |
| 合计Total | 29 | 53 |
以2 cm为分类单元的径级结构图显示, JWY样地中总独立个体数的径级分布呈明显的倒“J”型特征(图 5)。根据这一分布特征的实际情况, 将样地内林木胸径分为DBH≤10 cm, 10 cm<DBH≤30 cm和DBH>30 cm的小、中、大径木3个径级区间。胸径处于这3个区间的个体数分别占总个体数的76.9%、17.8%和5.3%。主林层树种个体数占总个体数的12.1%, 其径级分布总体呈偏正态分布型(图 5), 并在胸径6和23 cm左右有两个明显的高峰, 此外, 在胸径65 cm左右还出现一个小峰。胸径处于小、中、大径木区间内的个体数分别占该林层总个体数的19.2%、41.1%和39.7%。次林层树种个体数占总个体数的52.3%, 径级分布呈现倒“J”型向“L”型过渡的特点(图 5), 约80%的个体分布在小径木区间内, 而属于大径木的个体数仅占该林层的0.8%。林下层物种个体数占总个体数的35.7%, 展现明显的“L”型分布(图 5), 超过90%的个体为小径木, 仅有16株大径级个体。

|
| 图 5 江西武夷山中亚热带南方铁杉针阔混交林样地内全部物种以及不同林层物种的径级结构图 Fig. 5 Diameter at breast height (DBH) distribution of all species as well as different vertical layers in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
重要值排名前5位树种的径级结构特征可总体归纳为三种类型。第一类为偏正态分布型, 且存在多峰现象, 物种为主林层主要构成的南方铁杉和多脉青冈。南方铁杉的径级分布分别在胸径8, 40和65 cm左右形成数个较为明显的峰(图 6), 小、中、大径木分别占物种总个体数的16.3%、30.6%和53.1%。多脉青冈的径级分布则分别在5和22 cm左右形成两个较为明显的峰(图 6), 分别有22.1%、49.0%和28.9%的个体分布在小、中、大径木区间。第二类为倒“J”向“L”过渡型, 物种为次林层优势种闽皖八角(图 6), 其个体数的78.6%为小径木, 且个体数随着胸径的增加而逐步减少。第三类为典型的“L”型, 物种为林下层优势种岩柃(图 6)与鹿角杜鹃(图 6), 它们均有超过95%的个体集中分布在小径木区间内, 仅有鹿角杜鹃的零星个体在大径木区间内间断分布。

|
| 图 6 江西武夷山中亚热带南方铁杉针阔混交林样地内5个主要物种的径级结构及种群个体大小不均等性特征 Fig. 6 Diameter at breast height (DBH) distribution and population inequality characteristics of five major species in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot Gini指数(G)用以描述种群内个体大小不均等性程度;Lorenz asymmetry指数(LA)用以描述大、小个体对不均等程度的贡献性 |
从大小不均等性的角度分析上述5个主要物种的径级结构特征发现(图 6), 乔木树种(南方铁杉, 多脉青冈和闽皖八角)的径级不均等性程度(即, Gini指数)普遍高于灌木(或小乔木)物种(岩柃和鹿角杜鹃)。此外, 次林层和林下层物种(闽皖八角、岩柃、鹿角杜鹃)种群径级Lorenz asymmetry(LA)指数均处于0.9—1.0之间, 说明这些物种的径级较小个体和径级较大个体对种群内个体大小差异性的影响程度相当。而主林层树种(南方铁杉和多脉青冈)的LA指数处于0.7—0.8之间, 说明径级较小个体是引起种群内个体大小差异性的主要原因。
2.4 空间分布通过对JWY样地内重要值排名前5位树种的空间总体分布特征的分析发现(表 3), 各树种分布的方差/均值比都大于1, 且都具有统计学上的显著性;此外, 聚块指数均大于1, 而负二项参数均小于1。这些源自不同分析方法的结果相互佐证, 明确表明这些树种在样地内都呈现出聚集分布的特征。通过对比各物种间的参数指标发现, 这5个物种的种群聚集分布程度由强到弱依次为岩柃、南方铁杉、鹿角杜鹃、闽皖八角、多脉青冈。结合样地的地形因素分析发现(图 7), 南方铁杉、岩柃和鹿角杜鹃在上、中、下坡位均有大量分布, 而多脉青冈和闽皖八角主要分布在中、下坡位。此外, 相较于坡沟, 南方铁杉、岩柃、多脉青冈和鹿角杜鹃更倾向于生长在坡脊。而闽皖八角并不存在对坡沟或坡脊的偏好性。
| 物种 Species | 径级 DBH class | 方差/均值比法 Variance to mean ratio test | 聚块指数 Patchiness index | 负二项参数 Negative binomial index | 结论 Conclusion | |
| 比值 Ratio | t值 t value | |||||
| 南方铁杉 | 种群整体 | 2.10 | 39.47* | 4.16 | 0.32 | 聚集分布 |
| Tsuga chinensis var. tchekiangensis | 小径木 | 2.19 | 42.58* | 21.87 | 0.05 | 聚集分布 |
| 中径木 | 2.13 | 40.59* | 11.60 | 0.09 | 聚集分布 | |
| 大径木 | 1.12 | 4.41* | 1.67 | 1.50 | 聚集分布 | |
| 岩柃 | 种群整体 | 5.77 | 170.47* | 5.99 | 0.20 | 聚集分布 |
| Eurya saxicola | 小径木 | 5.80 | 171.64* | 6.21 | 0.19 | 聚集分布 |
| 中径木 | 1.19 | 6.81* | 6.48 | 0.18 | 聚集分布 | |
| 大径木 | — | — | — | — | — | |
| 多脉青冈 | 种群整体 | 1.39 | 14.01* | 2.74 | 0.57 | 聚集分布 |
| Cyclobalanopsis multinervis | 小径木 | 1.22 | 7.82* | 5.41 | 0.23 | 聚集分布 |
| 中径木 | 1.19 | 6.73* | 2.71 | 0.59 | 聚集分布 | |
| 大径木 | 1.19 | 6.75* | 3.91 | 0.34 | 聚集分布 | |
| 闽皖八角 | 种群整体 | 2.48 | 52.86* | 2.86 | 0.54 | 聚集分布 |
| Illicium minwanense | 小径木 | 2.53 | 54.62* | 3.45 | 0.41 | 聚集分布 |
| 中径木 | 1.33 | 11.92* | 2.98 | 0.51 | 聚集分布 | |
| 大径木 | — | — | — | — | — | |
| 鹿角杜鹃 | 种群整体 | 3.00 | 71.55* | 3.80 | 0.36 | 聚集分布 |
| Rhododendron latoucheae | 小径木 | 3.01 | 72.02* | 3.96 | 0.34 | 聚集分布 |
| 中径木 | 1.18 | 6.40* | 6.39 | 0.19 | 聚集分布 | |
| 大径木 | — | — | — | — | — | |
| *, P < 0.05;—, 因样本数量过少而失去统计意义。指数解读:(1)在T检验结果显著的基础上, 方差/均值比>1代表聚集分布, < 1代表均匀分布;(2)聚块指数 < 1为均匀分布, =1为随机分布, >1为聚集分布;(3)负二项参数值越小聚集程度越大, 而参数值趋于无穷大(一般8以上), 则接近随机分布[26] | ||||||

|
| 图 7 江西武夷山中亚热带南方铁杉针阔混交林样地内5个主要物种小、中、大径级个体的空间分布 Fig. 7 Spatial distribution of 5 major species in small, medium and large classes of diameter at breast height (DBH) in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot 小径木(DBH ≤ 10 cm);中径木(10 cm < DBH ≤ 30 cm);大径木(DBH > 30 cm) |
通过进一步对上述各树种的小、中、大径木个体空间分布特征的分析发现(表 3), 南方铁杉的聚集分布程度由小径木至大径木急剧下降, 大径木的聚块指数甚至接近于1。对于另一主林层树种多脉青冈来说, 小径木的聚集程度较高, 而中、大径木之间处于相似水平。主要以中、小径木形式存在的其他3个次林层或林下层物种中, 岩柃和闽皖八角的聚集程度在两个径级间的变化不大;而鹿角杜鹃自小径木至中径木的聚集程度有小幅度增加。结合样地的地形因素分析发现(图 7), 南方铁杉在山坡上的分布重心随着径级等级的减小而逐渐上移, 而对于其他物种来说, 未发现类似情况。此外, 所有物种均显示出小径木分布区域与中、大径木分布区域高度重叠的特征。
其他树种的分布格局亦展现出种间特异性特征。例如, 青冈和中华石楠(Photinia beauverdiana)在上、中、下坡位均有较多分布, 但具体分布区域却显示出互补性特征(图 8)。同时, 还有一些树种展现出对坡位的选择性。例如, 同样为杜鹃花属, 云锦杜鹃主要分布在样地的右上坡区域, 而猴头杜鹃(Rhododendron simiarum)则主要集中分布在样地的左下坡区域(图 8)。此外, 微毛柃(Eurya hebeclados)主要散布在下坡位和坡沟, 而黄山松却集中分布在坡顶的一小块坡脊区域(图 8)。

|
| 图 8 江西武夷山中亚热带南方铁杉针阔混交林样地内6个其他物种的空间分布 Fig. 8 Spatial distribution of 6 other species in Jiangxi Wuyishan mid-subtropical coniferous (Tsuga chinensis var. tchekiangensis) and broadleaf mixed forest plot |
大型固定样地相较于传统的小型、暂时性样地的核心优势, 除了可以在较大面积范围内对研究对象进行长期动态监测外, 还在于其具有标准化的设置规范和调查方法, 这使得样地间进行科学系统地定性和定量对比分析成为可能[6-9]。通过比较JWY样地与国内已建立的数个具有相同气候带(即, 亚热带)或类似林分类型(即, 针阔混交林或针叶林)的森林固定样地(表 4)的植物区系特征发现, JWY样地的植物区系组成在科水平上与广东鼎湖山[11]、福建武夷山[15]、浙江乌岩岭[16]、古田山[12]、天童山[13]、湖南八大公山[14]等亚热带阔叶林样地相同, 均以泛热带分布区类型为代表的热带成分为主。但在属水平的区系组成上, JWY样地与同处于中亚热带海拔1000 m以上的八大公山常绿落叶阔叶混交林样地[14]一样, 已经转变为以北温带分布区类型最多的温带成分为主。而北温带类型也恰恰是山西关帝山[27]、吉林长白山云冷林和落叶松林[28]等温带样地的主要区系类型。这说明, 高海拔使得处于中亚热带的JWY样地具有由亚热带向温带过度的森林群落特征。
| 文献 Reference | 地区 Location | 样地坐标 Coordinates | 气候带 Climate zone | 林分类型 Forest type | 海拔 Elevation /m | 年均降 水量△ Annual mean precipitation/ mm | 面积 Area/ hm2 | 科数 No. of families | 属数 No. of genera | 种数 No. of species | 属级主要 区系成分 Most abundant genera areal types | 独立个体数 Independent abundance /(/hm2) | 胸高断面积 Basal area /(m2/hm2) |
| [11] | 广东鼎湖山 | 23°10′26″ N, 112°32′10″ E | 南亚热带 | 常绿阔叶林 | 230—470 | 1985 | 20 | 59 | 119 | 210 | 热带 | 3581 | 30.2 |
| [15] | 福建武夷山 | 27°35′37″ N, 117°45′54″ E | 中亚热带 | 常绿阔叶林 | 470—520 | 1486—2100 | 1.44 | 34 | 65 | 139 | 热带 | 5014 | 34.0 |
| [16] | 浙江乌岩岭 | 27°42′16″ N, 119°40′13″ E | 中亚热带 | 常绿阔叶林 | 869—1144 | 2195.8 | 9 | 47 | 92 | 200 | 热带 | 7933 | 44.6 |
| [12] | 浙江古田山 | 29°15′13″ N, 118°07′12″ E | 中亚热带 | 常绿阔叶林 | 446—715 | 1963.7 | 24 | 49 | 103 | 159 | 热带 | 5863 ╪ | 36.9 |
| [14] | 湖南八大公山 | 29°46′2″ N, 110°5′15″ E | 中亚热带 | 常绿落叶阔 叶混交林 | 1355—1456 | 2105.4 | 25 | 53 | 114 | 238 | 温带 | 7462 ╪ | — |
| [13] | 浙江天童山 | 29°48′49″ N, 121°47′07″ E | 中亚热带 | 常绿阔叶林 | 304—603 | — | 20 | 51 | 94 | 152 | 热带 | 4730 | 39.0 |
| [17] | 湖北七姊妹山 | 30°02′05″ N, 109°43′50″ E | 中亚热带 | 常绿落叶阔 叶混交林 | 1356—1486 | 1876 | 6 | 57 | 124 | 245 | — | 6099 | 27.9 |
| [27] | 山西关帝山 | 37°51′ N, 111°27′ E | 暖温带 | 针叶林 | 1976—2039 | 822.6 | 4 | 11 | 22 | 30 | 温带 | 2567 | 45.6 |
| [28] | 吉林长白山 落叶松林 | 42°04′ N, 128°14′ E | 温带 | 针阔混交林 | 1425—1435 | 695—1340 | 4 | 8 | 16 | 22 | 温带 | 924 | 41.1 |
| [28] | 吉林长白山 云冷杉林 | 42°08′ N, 128°08′ E | 温带 | 针叶林 | 1244—1252 | 695—1340 | 4 | 6 | 12 | 22 | 温带 | 2160 | 54.7 |
| [29] | 吉林长白山 红松林 | 42°23′ N, 128°05′ E | 温带 | 针阔混交林 | 792—810 | 600—1340 | 25 | 18 | 32 | 52 | — | 1556 | 43.2 |
| [30] | 黑龙江小兴 安岭 | 47°10′50″ N, 128°53′20″ E* | 温带 | 针阔混交林 | 425—508 | 676 | 9 | 20 | 34 | 48 | — | 2373 | 42.3 |
| *, 样地所在地区(保护区)的坐标;—, 未明确报告;╪, 未明确说明是否是按独立个体计数;△, 样地所在地区(保护区)的年均降水量 | |||||||||||||
此外, 与同处亚热带(即, 相近维度)的低海拔阔叶林样地(表 4)相比, JWY样地内木本植物的科、属和物种数均显著偏少(表 5);而与相似林分类型的温带(即, 较高纬度)样地(表 4)相比, 其科、属和物种数又占明显优势(表 5)。这一结果可能反映了与温度相关的海拔梯度和纬度梯度对生物多样性的影响, 即, 生物多样性随着海拔和纬度的增加总体上呈现下降的趋势[31-32]。需要指出的是, 很多关于生物多样性与海拔梯度相关关系的研究指出, 具体的变化趋势还受到气候等因素的影响[33]。例如, 湿润山地的生物多样性大多随海拔的上升而递减;而在干旱山地则呈现出海拔中段多样性最高的格局[34]。由于本文所比较的亚热带样地均来自我国东、南部年均降水量在1500 mm, 甚至2000 mm以上(表 4)的湿润地区, 因此JWY样地与这些同处于亚热带的较低海拔样地的差异可能体现了湿润山地生物多样性对海拔的响应特征。另一个值得关注的现象是:与亚热带低海拔阔叶林样地相比JWY样地单位面积独立个体数显著偏低, 胸高断面积水平却具有相似水平;而与较高纬度的温带针叶(阔叶混交)林相比, 其单位面积独立个体数相似而胸高断面积水平又显著偏小(表 5)。由于反映林分密度的单位面积独立个体数和反映林分蓄积量的胸高断面积水平受物种组成、林龄和立地条件等多种因素的影响[35], 使得对上述现象很难进行精准解释。然而, 由于JWY样地和所选取的其他样地均为成熟稳定的原生或典型性森林群落, 且上述比较的结果具有统计学效力, 因此该现象可能在一定程度上反映了森林的林分密度和蓄积量对海拔和纬度变化的响应。
| 特征 Characteristics | 本研究样地 The plot of current research | 表 4亚热带阔叶林样地 Subtropical broadleaf forest plots in Table 4 | 表 4温带针叶(阔叶混交)林样地 Temperate coniferous (and broadleaf mixed) forest plots in Table 4 | |||
| 均值95%置信区间 95% confidential interval of the mean | t值 t value | 均值95%置信区间 95% confidential interval of the mean | t值 t value | |||
| 科数No. of families | 29 | 42.4—57.6 | 6.75*** | 5.0—20.2 | -5.96** | |
| 属数No. of genera | 53 | 82.9—120.3 | 6.36*** | 11.2—35.2 | -6.9** | |
| 种数No. of species | 89 | 152.6—231.1 | 6.41*** | 17.0—52.6 | -8.46** | |
| 独立个体数 Independent abundance/hm2 | 2252 | 4394.4—7229.0 | 6.15*** | 1081.7—2750.3 | -1.12NS | |
| 胸高断面积 Basal area/(m2/hm2) | 37.89 | 29.0—41.8 | -0.99NS | 38.6—52.2 | 3.06* | |
| *** P < 0.001;** P < 0.01;* P < 0.05;NS P > 0.05 | ||||||
于此同时, JWY样地在群落径级结构和物种分布格局方面又表现出与其他森林样地(表 4)的共性特征。例如, 所有森林样地(包括JWY样地)的木本植物总径级分布均呈现明显的倒“J”型特征, 表明各样地的群落更新处于良好稳定状态[12]。对于不同林层的主要优势树种而言, 构建主林层的大型乔木树种多为偏正态的径级分布特征, 且常伴有数个明显的峰, 种群内以中径木或大径木主, 小径木相对较少[11, 13, 29];构建次林层的乔木树种多表现为倒“J”型分布特征, 种群内以小径木为主, 并随着胸径的增加个体数逐渐减少[11, 13, 29];而构成林下层的灌木(小乔木)树种则以“L”型分布为主, 种群内绝大多数为小径木, 大径木极少[11, 13, 29]。在物种分布格局方面, 所有森林样地(包括JWY样地)的绝大多数树种均呈现不同程度的聚集性分布特征, 且很多树种, 尤其是主林层树种的种群聚集程度常表现出随径级增加而降低的趋势[12, 15, 27-28, 30]。这可能是密度制约作用的体现[12]。此外, 绝大多数树种都呈现出各具特点的生境偏好性, 如方位[28]、海拔[11]、坡向[30]、坡度[14]、山脊山谷[17]等。这即反映了植物生长和生态习性的种间特异性[27], 也暗示了生态位分化机制在生物多样性维持方面的重要作用[12]。
南方铁杉在JWY样地森林群落当前的演替阶段是绝对的优势种。它以占样地总个体数6.2%的数量贡献了样地总胸高断面积的46.7%, 是主林层的主要构建者。然而, 基于径级结构和空间分布的分析发现, 南方铁杉的种群在样地中的更新较为缓慢。径级相对较小的个体成为种群内个体大小差异性的主要贡献者, 说明幼树已在该种群中成为“异于主流的少数派”。这与按小、中、大径木区间划分的结构相一致:样地内超过50%的种群个体处于大径木阶段, 而小径木仅占整个种群的16%左右。由于样地处于山体背阴面(西北坡), 南方铁杉小径木对中、上坡位的生境偏好暗示了光源限制可能是该种群更新缓慢的一个重要原因。
本研究基于江西武夷山中亚热带南方铁杉针阔混交林6.4 hm2动态监测固定样地的首次木本植物调查, 较为全面、系统地介绍样地内DBH≥1 cm的木本植物物种组成、区系特征、径级结构和空间分布, 并就其海拔较高而纬度较低的特点与国内同气候带而林分类型不同的样地以及相似林分类型而气候带不同的样地进行比较。研究结果充分显示设置该样地对中国森林多样性监测以及南方铁杉种群保育的重要意义。
致谢: 感谢江西武夷山国家级自然保护区管理局程松林、刘江南、陈凤彬、祝于红、程林、王小民、郑元庆、袁荣斌、吴淑玉、杨涛、雷平、兰文军、董政、贾文军、郭洪兴和邹思成在样地设置及群落调查中给予的帮助。| [1] | Cardinale B J, Duffy J E, Gonzalez A, Hooper D U, Perrings C, Venail P, Narwani A, Mace G M, Tilman D, Wardle D A, Kinzig A P, Daily G C, Loreau M, Grace J B, Larigauderie A, Srivastava D S, Naeem S. Biodiversity loss and its impact on humanity. Nature, 2012, 486(7401): 59–67. DOI:10.1038/nature11148 |
| [2] | Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature, 2007, 448(7150): 188–190. DOI:10.1038/nature05947 |
| [3] | Levine J M, Bascompte J, Adler P B, Allesina S. Beyond pairwise mechanisms of species coexistence in complex communities. Nature, 2017, 546(7656): 56–64. DOI:10.1038/nature22898 |
| [4] | Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning:current knowledge and future challenges. Science, 2001, 294(5543): 804–808. DOI:10.1126/science.1064088 |
| [5] | Tilman D, Reich P B, Knops J M H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature, 2006, 441(7093): 629–632. DOI:10.1038/nature04742 |
| [6] | Condit R. Research in large, long-term tropical forest plots. Trends in Ecology & Evolution, 1995, 10(1): 18–22. |
| [7] | Hubbell S P, Foster R B. Commonness and rarity in a Neotropical forest: implications for tropical tree conservation//Soulé M E, ed. Conservation Biology: The Science of Scarcity and Diversity. Sunderland, Massachusetts: Sinauer Associates, 1986: 205-231. |
| [8] | Condit R. Tropical Forest Census Plots. Berlin, Heidelberg:Springer-Verlag, 1988. |
| [9] | Rees M, Condit R, Crawley M, Pacala S, Tilman D. Long-term studies of vegetation dynamics. Science, 2001, 293(5530): 650–655. DOI:10.1126/science.1062586 |
| [10] | 中国森林生物多样性监测网络.[2017-08-16]. http://www.cfbiodiv.org/index.asp. |
| [11] | 叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274–286. DOI:10.3773/j.issn.1005-264x.2008.02.005 |
| [12] | 祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 2008, 32(2): 262–273. DOI:10.3773/j.issn.1005-264x.2008.02.004 |
| [13] | 杨庆松, 马遵平, 谢玉彬, 张志国, 王樟华, 刘何铭, 李萍, 张娜, 王达力, 杨海波, 方晓峰, 阎恩荣, 王希华. 浙江天童20ha常绿阔叶林动态监测样地的群落特征. 生物多样性, 2011, 19(2): 215–223. |
| [14] | 卢志军, 鲍大川, 郭屹立, 路俊盟, 王庆刚, 何东, 张奎汉, 徐耀粘, 刘海波, 孟红杰, 黄汉东, 魏新增, 廖建雄, 乔秀娟, 江明喜, 谷志容, 廖春林. 八大公山中亚热带山地常绿落叶阔叶混交林物种组成与结构. 植物科学学报, 2013, 31(4): 336–344. |
| [15] | 丁晖, 杨云方, 徐海根, 方炎明, 陈晓, 杨青, 伊贤贵, 徐辉, 温小荣, 徐鲜均. 武夷山典型常绿阔叶林物种组成与群落结构. 生态学报, 2015, 35(4): 1142–1154. |
| [16] | 仲磊, 张杨家豪, 卢品, 顾雪萍, 雷祖培, 蔡延奔, 郑方东, 孙义方, 于明坚. 次生常绿阔叶林的群落结构与物种组成:基于浙江乌岩岭9ha森林动态样地. 生物多样性, 2015, 23(5): 619–629. |
| [17] | 姚良锦, 姚兰, 易咏梅, 艾训儒, 冯广, 刘峻城, 林勇, 黄伟, 丁易, 臧润国. 湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构. 生物多样性, 2017, 25(3): 275–284. |
| [18] | 陈昌笃. 论武夷山在中国生物多样性保护中的地位. 生物多样性, 1999, 7(4): 320–326. DOI:10.3321/j.issn:1005-0094.1999.04.011 |
| [19] | 刘信中, 方福生. 江西武夷山自然保护区科学考察集. 北京: 中国林业出版社, 2001. |
| [20] | 矫恒盛, 钟志宇, 程林, 方毅, 程松林. 江西武夷山自然保护区森林群落木本植物多样性垂直规律研究. 江西林业科技, 2009(1): 6–10. DOI:10.3969/j.issn.1006-2505.2009.01.002 |
| [21] | 袁荣斌, 邹思成, 兰文军, 徐新宇. 江西武夷山国家级自然保护区南方铁杉资源调查初报. 江西林业科技, 2012(4): 37-39, 60-60. DOI:10.3969/j.issn.1006-2505.2012.04.012 |
| [22] | 中国植物志.[2017-08-16]. http://frps.eflora.cn/. |
| [23] | 吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1999(S4): 1–139. |
| [24] | 吴征镒, 周浙昆, 李德铢, 彭华, 孙航. 世界种子植物科的分布区类型系统. 云南植物研究, 2003, 25(3): 245–257. DOI:10.3969/j.issn.2095-0845.2003.03.001 |
| [25] | Damgaard C, Weiner J. Describing inequality in plant size or fecundity. Ecology, 2000, 81(4): 1139–1142. DOI:10.1890/0012-9658(2000)081[1139:DIIPSO]2.0.CO;2 |
| [26] | 郑元润. 不同方法在沙地云杉种群分布格局分析中的适用性研究. 植物生态学报, 1997, 21(5): 480–484. DOI:10.3321/j.issn:1005-264X.1997.05.010 |
| [27] | 杨秀清, 史婵, 王旭刚, 马慧晶, 闫海冰. 关帝山寒温性云杉次生林样地树种组成、结构和空间分布. 应用生态学报, 2017, 28(5): 1421–1430. |
| [28] | 匡旭, 邢丁亮, 张昭臣, 宋厚娟, 王芸芸, 房帅, 原作强, 叶吉, 蔺菲, 王绪高, 郝占庆. 长白山北坡云冷杉林和落叶松林物种组成与群落结构. 应用生态学报, 2014, 25(8): 2149–2157. |
| [29] | 郝占庆, 李步杭, 张健, 王绪高, 叶吉, 姚晓琳. 长白山阔叶红松林样地(CBS):群落组成与结构. 植物生态学报, 2008, 32(2): 238–250. DOI:10.3773/j.issn.1005-264x.2008.02.002 |
| [30] | 徐丽娜, 金光泽. 小兴安岭凉水典型阔叶红松林动态监测样地:物种组成与群落结构. 生物多样性, 2012, 20(4): 470–481. |
| [31] | MacArthur R H. Geographical Ecology:Patterns in the Distribution of Species. New York: Harper & Row, 1972. |
| [32] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220–227. DOI:10.1038/35012228 |
| [33] | McCain C M, Grytnes J A. Elevational gradients in species richness//Encyclopedia of Life Sciences. Chichester: John Wiley & Sons Ltd, 2010. |
| [34] | Grytnes J A, McCain C M. Elevational trends in biodiversity//Levin S A, ed. Encyclopedia of Biodiversity. Amsterdam: Elsevier, 2007: 1-8. |
| [35] | 王斌, 黄俞淞, 李先琨, 向悟生, 丁涛, 黄甫昭, 陆树华, 韩文衡, 文淑均, 何兰军. 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布. 生物多样性, 2014, 22(2): 141–156. |