 2018, Vol. 38
2018, Vol. 38文章信息
- 王秋玲, 周广胜.
- WANG Qiuling, ZHOU Guangsheng.
- 春玉米持续干旱过程中常用气孔导度模型的比较研究
- Comparisons between common stomatal conductance models under progressive drought in spring maize
- 生态学报. 2018, 38(19): 6846-6856
- Acta Ecologica Sinica. 2018, 38(19): 6846-6856
- http://dx.doi.org/10.5846/stxb201711082000
-
文章历史
- 收稿日期: 2017-11-08
- 网络出版日期: 2018-06-20





2. 南京信息工程大学应用气象学院, 南京 210044;
3. 南京信息工程大学气象灾害预警协同创新中心, 南京 210044
2. College of Applied Meteorology, Nanjing University of Information Science & Technology, Nanjing 210044, China;
3. Collaborative Innovation Center on Forecast Meteorological Disaster Warning and Assessment, Nanjing University of Information Science & Technology, Nanjing 210044, China
干旱胁迫是在世界范围内限制植物生产的最主要因素之一[1-2]。全球气候变化被认为对植物生长区域干旱频率和程度均有影响[3-4]。气孔控制叶片和大气之间的水汽(H2O)和二氧化碳(CO2)传输, 影响植物的生产力和水分利用效率[5], 进而影响全球碳循环和水循环[6]。植物叶片气孔导度受辐射、温度和水汽压亏缺等环境因子影响[7], 木质部汁液中的脱落酸(ABA)和叶片水势均参与植株水平的气孔控制, 且对不同物种的作用不同[8]。在对气孔的环境响应机理认识不足时, 模型模拟成为最有效和适宜的工具[9-10]。
目前, 常用气孔导度模型主要包括基于气孔导度与环境因子关系建立的经验模型[11]、基于气孔导度与光合作用关系建立的半经验模型[12-13]以及基于最优气孔行为理论建立的模型[14]。气孔导度模型不仅被广泛用于全球气候模式[15-16], 而且被用于各类生态系统模拟, 如小麦[17-18]、玉米[19-20]、森林[10, 21-22]、草地[23]和芦苇湿地[24]等生态系统。研究发现, 气孔导度模型的模拟准确性受研究对象、试验区域、环境条件、时间尺度的影响[25]。现有研究在选择气孔导度模型时缺乏相关依据[26], 不同研究者对于不同类型生态系统、甚至同一类型生态系统所用的气孔导度模型并不相同, 在模拟干旱影响时, 未考虑到模型的适用范围, 难以准确反映水分亏缺程度[27]。
玉米是世界第一大粮食作物, 在整个生长期间对干旱非常敏感[28]。作为东北地区的主栽作物之一, 玉米播种面积占东北三省粮食作物播种面积的55.2%, 年产量占全国玉米产量的33.8%[29]。准确模拟水分胁迫条件下玉米的气孔行为, 对于理解玉米对干旱的响应和适应机制具有重要意义[30]。为此, 本研究以春玉米为研究对象, 基于拔节期-抽雄期持续干旱田间模拟试验的气体交换参数观测资料, 研究4种常用气孔导度模型在干旱过程的适用性, 揭示引入土壤水分响应函数对不同气孔导度模型模拟效果的影响, 同时探讨4种常用气孔导度模型适用的土壤水分范围, 为气孔导度模型在干旱区域的应用及作物干旱监测和预警提供依据。
1 研究地区与研究方法 1.1 试验设计和环境条件试验在辽宁省锦州市生态与农业气象中心进行(41°09′N, 121°12′E, 海拔27.4 m)。试验区地处亚欧大陆东北部, 属温带半湿润季风气候。该区多年(1981—2010)平均气温为9.9℃, 平均降水量为568 mm, 年极端最高气温41.8℃, 年极端最低气温-31.3℃, 年无霜期144—180 d。试验区土壤为典型棕壤土, pH值为6.3, 有机质1.8%, 全氮含量0.1%, 0—100 cm土壤田间持水量平均值22.3%, 凋萎湿度6.5%, 容重1.616 g/cm3[31]。
控制试验于2016年在农田土壤水分控制试验场进行。试验小区为5 m×3 m, 小区间有深2 m、宽0.15 m水泥隔离层防止不同水分控制区之间横向水分交换。试验小区上方有大型电动防雨棚用于遮挡自然降水。供试玉米品种为丹玉405, 为中熟品种。2016年5月23日播种, 2016年9月28日收获。玉米行距为50 cm, 株距为35 cm, 种植密度与大田一致, 为5.3株/m2。播种后施肥, 所用肥料为控得安控释掺混肥料, 总养分≥ 50%, 控释氮≥ 21%, 每公顷600 kg。拔节前所有处理均充分灌水, 6月29日进行最后一次灌水后处理开始。试验设对照(Control)、干旱处理1(Drought 1, 简称D1)和干旱处理2(Drought 2, 简称D2)共3个处理, 每个处理3个重复。对照处理Control在全生育期内维持土壤水分条件适宜; D1处理为拔节期开始控水4周后(7月27日)复水, D2处理为拔节期开始控水6周后(8月10日)复水; 复水参照对照处理。每隔7d进行一次观测, 共取样观测5次。观测时各处理春玉米的发育期见表 1。为研究气孔导度模型在春玉米持续干旱过程中的适用性, 气孔导度模拟数据将只选取复水前数据(n=39)。
| 处理 Treatment |
7月1日 July 1st |
7月7日 July 7th |
7月14日 July 14th |
7月22日 July 22th |
7月29日 July 29th |
| 对照Control | 拔节初期 | 拔节期 | 大喇叭口期 | 大喇叭口期 | 抽雄期 |
| 干旱处理1 Drought 1 | 拔节初期 | 拔节期 | 大喇叭口期 | 大喇叭口期 | 抽雄期 |
| 干旱处理2 Drought 2 | 拔节初期 | 拔节期 | 大喇叭口期 | 大喇叭口期 | 抽雄期 |
采用烘干称重法测定0—50 cm每10 cm土壤分层含水量, 每个处理均有3个重复。土壤含水量θ的计算公式如下:

|
(1) |
土壤相对湿度SRWC(%)的计算式如下:
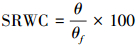
|
(2) |
式中, WC是湿土和空盒重(g), WP是空盒重(g), WD是干土和空盒重(g), θf是田间持水量(%)。
1.2.2 叶片气体交换参数选择晴朗无云天气9:30—11:30进行叶片气体交换参数的观测。利用LI-6400便携式光合作用系统(LI-COR Inc., Lincoln, NE, USA), 采用荧光叶室测定春玉米顶部第一片展开叶光合速率(An, μmol m-2 s-1)、蒸腾速率(Tr, mmol m-2 s-1)、气孔导度(gs, mol m-2 s-1)等叶片气体交换参数以及叶温(Tl, ℃)、大气CO2浓度(μmol/mol)、相对湿度(hs)等环境因子。叶室内PAR设置为1500 μmol m-2 s-1, 该值为玉米光饱和平均值。叶室内温度和CO2浓度与外界一致, 流速设置为300 μmol/s。每个处理选择3株玉米。
1.3 气孔导度模型 1.3.1 Jarvis模型Jarvis模型[11]是典型阶乘型的经验模型, 模型假设不考虑变量之间的协同作用, 是一连串单因子校正系数的函数。模型主要包括5个环境变量, 即辐射、温度、湿度、CO2浓度和叶片水势。本研究中, 不考虑辐射和外界CO2浓度的影响, 水汽压亏缺D的影响函数采用Leuning提出的形式[13], 温度T的影响函数采用Hofstra和Hesketh提出的二次曲线方程[32]:
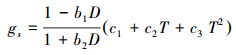
|
(3) |
式中, D表示水汽压亏缺(kPa), b1、b2、c1、c2、c3均为拟合参数。
1.3.2 Ball-Woodrow-Berry模型Ball等考虑到气孔具有维持胞间CO2浓度为常数的功能, 在稳定状态下当叶片表层CO2浓度和大气湿度不变时, 气孔导度与净光合速率具有线性关系, 据此提出了Ball-Woodrow-Berry模型[12] (简称BWB模型)。
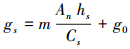
|
(4) |
式中, Cs表示叶片表面CO2浓度(μmol/mol), m、g0为拟合参数。
1.3.3 Ball-Berry-Leuning模型由于BWB模型不能描述在低CO2浓度时气孔行为, 在Cs达到CO2补偿点Γ时, 不能准确模拟气孔导度。Leuning将模型中Cs修正为Cs-Γ[33]。另外, 由于气孔对D响应比hs更直接, Leuning进一步修正模型, 将模型中hs修正为D的函数[13] (简称BBL模型)。
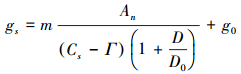
|
(5) |
式中, Γ表示CO2补偿点(μmol/mol), m、g0和D0为拟合参数。按照Brooks和Farquhar提出的经验方法[34], C3植物的补偿点与叶温存在关系ΓC3=42.7+1.68 (Tl-25) +0.012 (Tl-25) 2, 根据Yu等研究, C4植物的补偿点约为C3植物的1/10[20], 即ΓC4=0.1 ΓC3, 以此计算春玉米CO2补偿点。
1.3.4 最优气孔导度模型Medlyn等结合最优气孔行为理论[35], 认为气孔最大化碳获得的同时最小化水分损失, 并假设大气CO2浓度Ca远大于CO2补偿点, 且只考虑RuBP再生限制光合作用, 提出了以下模型[14](简称USO模型):
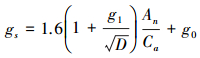
|
(6) |

|
(7) |
式中, g0和g1为拟合参数, g1与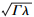
考虑到植物叶片气孔导度是大气干燥度、土壤供水能力与植物生物学特性共同作用的结果, 在本研究中引入土壤湿度的三段线性函数[36-37]作为水分响应函数以修正气孔导度模型:

|
(8) |
式中, θw、θ、θf分别为土壤凋萎湿度、土壤实际含水量和土壤田间持水量。0—50 cm平均田间持水量为21.8%, 土壤凋萎湿度为6.1%(均为质量含水量)。
引入水分响应函数后修正的Jarvis模型、BWB模型、BBL模型和USO模型分别简称Jarvis-M模型、BWB-M模型、BBL-M模型和USO-M模型。
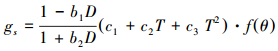
|
(9) |
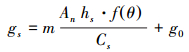
|
(10) |
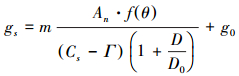
|
(11) |
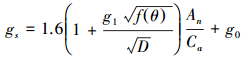
|
(12) |
试验观测数据采用Sigmaplot 12.5 (SYSTAT Software, USA)进行拟合, 确定模型参数并给出拟合度(R2)和显著性。以均方根误差(RMSE)、归一化均方根误差(NRMSE)、相对误差(RE)、模型斜率(b0)和AIC信息准则(Akaike information criterion)评价各模型模拟效果。RMSE、NRMSE、AIC、RE和b0的计算公式如下:
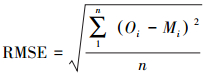
|
(13) |
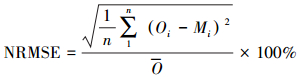
|
(14) |

|
(15) |

|
(16) |

|
(17) |
式中, Oi表示实测值; Mi表示观测值; n为样本数; k为模型中参数的个数。R2越接近1, 表明模型模拟效果越好; RMSE、NRMSE和RE值越小, 说明模拟效果越好; 模型斜率b0反映了模型造成高估或低估, 当b0>1时, 表示高估, b0 < 1时表示低估, b0越接近1, 模拟效果越好; AIC可以估计模型的复杂程度和模型拟合效果的优劣, 其值越小, 表示模型的模拟效果越好。
2 结果与分析 2.1 观测期间土壤水分及气孔导度变化春玉米持续干旱过程中土壤水分变化见图 1。观测期间对照处理Control的土壤相对湿度(SRWC)基本维持在65%±5%范围内。两个干旱处理D1和D2的SRWC均随处理时间延长而降低, 在7月22日, D1和D2的SRWC分别比对照处理Control的SRWC低43.94%和43.90%。7月27日处理D1复水, 在7月29日, 处理D1的SRWC有一定上升, 干旱处理D2的SRWC比对照处理Control的SRWC低51.62%。观测期间, D2处理的SRWC平均每天降低1.39%。观测期间各处理叶片气孔导度值见表 2。

|
| 图 1 观测期间各处理土壤水分变化 Fig. 1 The soil relative water content of each treatment during the experiment |
| 处理 Treatment |
7月1日 July 1st |
7月7日 July 7th |
7月14日 July 14th |
7月22日 July 22th |
7月29日 July 29th |
| 对照Control | 0.39±0.07 | 0.34±0.04 | 0.28±0.02 | 0.17±0.02 | 0.24±0.03 |
| 干旱处理1 Drought 1 | 0.39±0.03 | 0.30±0.05 | 0.21±0.05 | 0.18±0.03 | 0.18±0.01 |
| 干旱处理2 Drought 2 | 0.40±0.05 | 0.25±0.01 | 0.18±0.02 | 0.17±0.01 | 0.17±0.04 |
采用复水前观测数据进行气孔导度模拟, Jarvis模型、BWB模型、BBL模型和USO模型及其水分修正模型模拟结果见表 3, 所有模型均通过显著性检验(P < 0.05)。Jarvis模型、BWB模型、BBL模型和USO模型及其水分修正模型模拟值与观测值的线性关系见图 2。BWB模型、BBL模型和USO模型模拟值与观测值较接近; 引入土壤水分响应函数后, BWB-M模型和USO-M模型的模拟值分别较BWB模型和USO模型模拟值更接近观测值, 而BBL-M模拟值较BBL模型略远离观测值。未引入土壤水分响应函数时, 模型模拟效果表现为BBL模型R2最高, 其次是USO模型和BWB模型, Jarvis模型最低; 引入水分响应函数后, 模型模拟效果表现为USO-M模型R2最高, 其次是BBL-M模型和BWB-M模型, Jarvis-M模型最低。引入土壤水分响应函数后, BWB和USO模型的R2有所提高, 分别由0.77和0.89提高到0.81和0.91;而Jarvis和BBL模型的R2反而降低, 分别由0.56和0.92下降到0.38和0.85。
| 模型 Models |
参数Parameters | R2 | F | P | ||||
| b1 | b2 | c1 | c2 | c3 | ||||
| Jarvis | 1.5462 | 2.0589 | -22.1183 | 1.1995 | -0.0165 | 0.56 | 10.704 | < 0.0001 |
| Jarvis-M | 0.7833 | -0.9006 | 14.6826 | -0.8619 | 0.0133 | 0.38 | 5.1067 | 0.0025 |
| m | g0 | |||||||
| BWB | 7.5426 | -0.0925 | 0.77 | 120.9157 | < 0.0001 | |||
| BWB-M | 6.3946 | 0.1314 | 0.81 | 162.0454 | < 0.0001 | |||
| m | g0 | D0 | ||||||
| BBL | 6.4979 | -0.0316 | 1.8512 | 0.92 | 197.3679 | < 0.0001 | ||
| BBL-M | 5.5813 | 0.1406 | 2.0727 | 0.85 | 106.0741 | < 0.0001 | ||
| g1 | g0 | |||||||
| USO | 1.4456 | -0.048 | 0.89 | 305.9229 | < 0.0001 | |||
| USO-M | 1.3143 | 0.015 | 0.91 | 370.777 | < 0.0001 | |||

|
| 图 2 气孔导度模型及其水分修正模型模拟结果 Fig. 2 Comparison of observed and estimated stomatal conductance (gs) 图中黑色虚线为1 : 1线 |
进一步给出了气孔导度模型模拟效果评价指标(表 4)。在持续干旱条件下, Jarvis模型和BBL模型在引入土壤水分响应函数后, 模拟效果均降低, 表现为b0远离1, 模型低估情况加重, RMSE、NRMSE、RE和AIC均有所增加, NRMSE分别增加4.70%和3.45%, RE分别上升6.02%和2.00%。Jarvis模型在引入土壤水分修正项后模拟效果下降幅度超过BBL模型。BWB模型和USO模型在引入土壤水分响应函数后, 模拟效果均提高, 表现为b0向1靠近, 模型低估情况改善, RMSE、NRMSE、RE和AIC均有所降低, NRMSE分别降低1.99%和1.02%, RE分别降低3.20%和0.63%。BWB模型在引入水分响应函数后模拟效果改进幅度较USO模型大。总体来看, 未引入土壤水分响应函数时, 模型模拟效果表现为BBL模型最优, 其次是USO模型, BWB模型, Jarvis模型最差; 在引入土壤水分响应函数后, 模型模拟效果表现为USO-M模型最优, 其次是BBL-M模型, BWB-M模型, Jarvis-M模型最差。引入土壤水分响应函数对Jarvis模型和BWB模型的模拟效果影响较大, 对BBL模型和USO模型的模拟效果影响较小。
| 模型 Models |
均方根误差 RMSE Root mean square error/ (mol m-2 s-1) |
归一化均方根误差NRMSE Normalized root mean square error/% |
相对误差RE Relative error/% |
模型斜率b0 Model slope |
AIC信息准则 Akaike information criterion |
| Jarvis | 0.0661 | 25.00 | 22.93 | 0.945 | -200 |
| Jarvis-M | 0.0785 | 29.70 | 28.95 | 0.923 | -186 |
| BWB | 0.0481 | 18.19 | 16.35 | 0.971 | -231 |
| BWB-M | 0.0428 | 16.20 | 13.15 | 0.977 | -240 |
| BBL | 0.0287 | 10.86 | 9.92 | 0.990 | -269 |
| BBL-M | 0.0378 | 14.31 | 11.91 | 0.982 | -247 |
| USO | 0.0326 | 12.34 | 11.32 | 0.992 | -261 |
| USO-M | 0.0299 | 11.32 | 10.69 | 0.993 | -268 |
根据剩余气孔导度(模拟值-观测值)与土壤相对湿度(SRWC)关系(图 3), 在低土壤水分下, Jarvis模型、BWB模型、BBL模型和USO模型均造成气孔导度高估, 且Jarvis模型和BWB模型高估较重; 在高土壤水分下, Jarvis模型、BWB模型、BBL模型和USO模型均造成气孔导度低估, 且Jarvis模型和BWB模型低估较重。引入土壤水分响应函数后, Jarvis-M模型在低土壤水分下严重低估, 高土壤水分下高估; BWB-M模型、BBL-M模型和USO-M模型的剩余气孔导度几乎不受土壤相对湿度的影响。

|
| 图 3 剩余气孔导度(模拟值-观测值)与SRWC关系 Fig. 3 The relationship between the residuals (estimated gs-observed gs) and soil relative water content 图中黑色虚线表示剩余气孔导度为0;黑色实线为初始模型剩余气孔导度与土壤相对含水量的线性关系; 红色实线为水分修正模型剩余气孔导度与土壤相对含水量的线性关系 |
总体来看, Jarvis模型在当前持续干旱条件下表现不稳定, 低估和高估幅度较大, 其次是BWB模型, 而BBL模型和USO模型在持续干旱条件下相对稳定, 低估和高估幅度较小。土壤水分响应函数的引入, 对Jarvis模型和BWB模型影响相对较大, 而对BBL模型和USO模型影响相对较小。
叶片气孔导度观测值及4种气孔导度模型及其水分修正模型的模拟值与SRWC所建立的回归方程, 均通过0.05水平显著性检验(表 5)。图 4给出了叶片气孔导度观测值与SRWC拟合曲线的95%置信区间及模拟值与SRWC拟合曲线。以模拟值与SRWC的拟合曲线是否超出观测值与SRWC拟合曲线的95%置信区间为判断依据, 在当前土壤水分条件下(33% < SRWC < 83%), BBL模型和USO模型模拟值基本在观测值置信区间范围内, 表明BBL模型和USO模型具有一定的稳定性, 无论是否引入水分响应函数, 在当前土壤水分条件下具有明显优势, 可以应用在持续干旱过程。BWB模型在SRWC > 76%时, 模拟值与SRWC的拟合曲线超出观测值与SRWC拟合曲线的95%置信区间, 在引入水分响应函数后, 模拟值基本在观测值置信区间范围内, 表明BWB模型的适用土壤水分范围为33% < SRWC < 76%, 在高于该范围时, 须引入水分响应函数。尽管Jarvis模型模拟效果一般, 但在当前土壤水分范围内(33% < SRWC < 83%)均适用, 在引入水分响应函数后, 在SRWC < 50%时超出范围, 即修正后的模型适用于SRWC > 60%。
| 模型 Models |
回归方程 Regression equations |
R2 | F | P |
| gs | y=0.1525-0.0015x+0.00005588x2 | 0.635 | 31.3158 | < 0.0001 |
| Jarvis | y=0.3248-0.0066x+0.000089913x2 | 0.6676 | 36.1547 | < 0.0001 |
| Jarvis-M | y=-0.17+0.007x+0.0000028045x2 | 0.9308 | 242.1215 | < 0.0001 |
| BWB | y=0.0053+0.0058x-0.00002058x2 | 0.4269 | 13.4092 | < 0.0001 |
| BWB-M | y=0.0354+0.0025x+0.000025158x2 | 0.9256 | 223.839 | < 0.0001 |
| BBL | y=0.0112+0.0044x-0.000000000000081642x2 | 0.574 | 24.2487 | < 0.0001 |
| BBL-M | y=-0.0395+0.0053x-0.000000000000010381x2 | 0.8864 | 140.4964 | < 0.0001 |
| USO | y=-0.0095+0.0053x-0.0000080023x2 | 0.5515 | 22.1339 | < 0.0001 |
| USO-M | y=-0.0268+0.0052x-0.0000020706x2 | 0.7098 | 44.0183 | < 0.0001 |

|
| 图 4 气孔导度模拟值及观测值与SRWC拟合曲线 Fig. 4 The fitting curves of stomatal conductance and SRWC 图中黑色虚线表示观测值与土壤相对含水量拟合曲线的95%置信区间上、下限 |
全球变暖, 森林砍伐和城市化预计将增加未来干旱的严重性和频率[3]。气孔导度体现了植物H2O保护和CO2捕获的矛盾需求之间的权衡[38]。气孔导度的准确模拟直接影响到植物生产力和水分利用效率[17], 以及植被与大气之间通量交换模拟的准确性[39]。鱼腾飞等[10]对极端干旱区多枝怪柳叶片气孔导度对空气湿度的响应研究表明, BWB模型模拟效果优于BBL模型。高冠龙等[21]研究表明, Jarvis模型对极端干旱条件下中幼龄胡杨林气孔导度的模拟精度高于BWB模型。Ji等[19]研究发现, USO模型无论是否引入水分响应函数在不同年际间玉米的模拟中均比BWB模型和Jarvis模型表现更稳定和更准确, 且USO-M模型比USO模型更准确。本研究中, BBL模型在春玉米持续干旱过程中表现最好, 其次是USO模型和BWB模型, Jarvis模型表现最差。研究结果的不同, 表明受研究对象、试验区域、环境条件、时间尺度等的影响, 气孔导度模型模拟效果有所不同, 在选择气孔导度模型时, 更应综合考虑这些影响因素。
当前, 大部分气孔导度模型的应用未考虑土壤水分变化对叶片气孔导度的影响[40], 主要原因是目前对土壤水分影响叶片气孔导度的机理认识仍然不足[27]。本研究中引入土壤湿度的三段线性函数表征土壤供水能力对气孔导度的影响, 结果表明, 在持续干旱条件下, 引入土壤水分响应函数会提高BWB模型和USO模型的拟合效果, 而降低Jarvis模型和BBL模型模拟效果。引入土壤水分响应函数后, 模型模拟效果表现为USO-M模型> BBL-M模型>BWB-M模型>Jarvis-M模型。以气孔导度模型模拟值与SRWC的拟合曲线是否超出观测值与SRWC拟合曲线的95%置信区间为判断依据, 在土壤水分范围为33% < SRWC < 83%条件下, 无论是否引入水分响应函数, BBL模型和USO模型均适用, 表明BBL模型和USO模型具有一定的稳定性。BWB模型的适用土壤水分范围为33% < SRWC < 76%, 在高于该范围时, 须引入土壤水分响应函数。尽管Jarvis模型模拟效果一般, 但在土壤水分范围在33% < SRWC < 83%条件下均适用。
Jarvis模型假设各环境因子独立作用于植物气孔, 忽视环境因子间的相互作用。模型中参数没有明确生物学意义, 且随品种和地域而变, 需要许多覆盖范围广泛的环境条件数据来估计参数值, 参数确定较复杂[41]。本研究中通过田间模拟试验获得的土壤湿度范围较宽, 但总体数据量较少, 可能是Jarvis模型模拟效果较差的原因之一, 未来可以考虑在干旱条件下进行光合-CO2浓度响应曲线和光合-光响应曲线的观测, 以获得覆盖更广范围的其他环境条件数据进一步估算Jarvis模型参数。水分胁迫会改变气孔导度与光合速率之间的线性关系, 因此, BWB模型在水分胁迫条件下不再适用。Zhang等[42]研究表明, 必须考虑气孔导度模型参数受干旱影响发生下降, 否则会高估气孔导度。BWB模型的参数个数较少, 便于对模型改进后用于极端环境条件。本研究中引入土壤水分响应函数对BWB模型进行修正, 发现修正后模型可在当前持续干旱条件下的土壤水分范围内适用。根据以往研究, BWB模型和BBL模型中参数m在C4植物的平均值为5.8±3.8[43], 本研究结果中, BWB模型中参数m在春玉米的值为7.5426, BBL模型中参数m的值为6.4979, 基本在平均范围内, 同时, 引入土壤水分响应函数后, 改进的BWB模型和BBL模型中参数m均有所降低, 与前人研究结果一致。基于最优气孔行为理论发展的USO模型中参数g1, 与边际水消耗λ具有一定的比例关系, 参数具有生物学含义, 可以用来描述植物的水分利用策略[14]。但λ与土壤湿度的定量关系在当前应用中采用经验拟合方法[16], 这限制了模型的应用。试验期间春玉米处于拔节期—抽雄期, 试验通过持续干旱控水达到的土壤相对湿度范围为33%—83%, 春玉米在更高水分区数据缺少, 难以反映各模型在更高水分区的适用性。未来应综合考虑研究对象、研究区域、环境条件等选择合适的气孔导度模型, 在模型应用中注意模型的适用土壤水分范围及干旱对气孔导度的影响, 以提高模型模拟植被应对水分亏缺的能力, 为准确预测气候变化背景下植物和大气之间的碳、水交换提供依据。
| [1] | Lobell D B, Roberts M J, Schlenker W, Braun N, Little B B, Rejesus R M, Hammer G L. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Science,, 2014, 344(6183): 516–519. DOI:10.1126/science.1251423 |
| [2] | Gholipoor M, Choudhary S, Sinclair T R, Messina C D, Cooper M. Transpiration response of maize hybrids to atmospheric vapour pressure deficit. Journal of Agronomy and Crop Science, 2013, 199(3): 155–160. DOI:10.1111/jac.2013.199.issue-3 |
| [3] | Cairns J E, Sonder K, Zaidi P H, Verhulst N, Mahuku G, Babu R, Nair S K, Das B, Govaerts B, Vinayan M T, Rashid Z, Noor J J, Devi P, Vicente F S, Prasanna B M. Chapter one-Maize production in a changing climate:impacts, adaptation, and mitigation strategies. Advances in Agronomy, 2012, 114: 1–58. DOI:10.1016/B978-0-12-394275-3.00006-7 |
| [4] | Sicher R C Jr, Kim S H. Photosynthesis, growth and maize yields in the context of global change//Prioul C, Thévenot M T, eds. Advances in Maize, Essential Reviews in Experimental Biology. London: Society of Experimental Biology, 2011, 3: 373-392. |
| [5] | Vialet-Chabrand S R M, Matthews J S A, McAusland L, Blatt M R, Griffiths H, Lawson T. Temporal dynamics of stomatal behavior:modeling and implications for photosynthesis and water use. Plant Physiology, 2017, 174(2): 603–613. DOI:10.1104/pp.17.00125 |
| [6] | Marchin R M, Broadhead A A, Bostic L E, Dunn R R, Hoffmann W A. Stomatal acclimation to vapour pressure deficit doubles transpiration of small tree seedlings with warming. Plant, Cell & Environment, 2016, 39(10): 2221–2234. |
| [7] | Bunce J A. Responses of stomatal conductance to light, humidity and temperature in winter wheat and barley grown at three concentrations of carbon dioxide in the field. Global Change Biology, 2000, 6(4): 371–382. DOI:10.1046/j.1365-2486.2000.00314.x |
| [8] | Tardieu F, Davies W J. Integration of hydraulic and chemical signalling in the control of stomatal conductance and water status of droughted plants. Plant, Cell & Environment, 1993, 16(4): 341–349. |
| [9] | Hetherington A M, Woodward F I. The role of stomata in sensing and driving environmental change. Nature, 2003, 424(6951): 901–908. DOI:10.1038/nature01843 |
| [10] | 鱼腾飞, 冯起, 司建华. 极端干旱区多枝柽柳叶片气孔导度的环境响应模拟. 植物生态学报, 2012, 36(6): 483–490. |
| [11] | Jarvis P G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philosophical Transactions of the Royal Society B:Biological Sciences, 1976, 273(927): 593–610. DOI:10.1098/rstb.1976.0035 |
| [12] | Ball J T, Woodrow I E, Berry J A. A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions//Biggins J, ed. Progress in Photosynthesis Research. Dordrecht: Springer, 1987: 221-224. |
| [13] | Leuning R. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant, Cell & Environment, 1995, 18(4): 339–355. |
| [14] | Medlyn B E, Duursma R A, Eamus D, Ellsworth D S, Prentice I C, Barton C V M, Crous K Y, De Angelis P, Freeman M, Wingate L. Reconciling the optimal and empirical approaches to modelling stomatal conductance. Global Change Biology, 2011, 17(6): 2134–2144. DOI:10.1111/gcb.2011.17.issue-6 |
| [15] | Bonan G B, Williams M, Fisher R A, Oleson K W. Modeling stomatal conductance in the earth system:linking leaf water-use efficiency and water transport along the soil-plant-atmosphere continuum. Geoscientific Model Development, 2014, 7(5): 2193–2222. DOI:10.5194/gmd-7-2193-2014 |
| [16] | De Kauwe M G, Kala J, Lin Y S, Pitman A J, Medlyn B E, Duursma R A, Abramowitz G, Wang Y P, Miralles D G. A test of an optimal stomatal conductance scheme within the CABLE land surface model. Geoscientific Model Development, 2015, 8(2): 431–452. DOI:10.5194/gmd-8-431-2015 |
| [17] | 魏征, 刘钰, 许迪, 蔡甲冰, 张宝忠. 基于叶片气孔导度提升的冬小麦冠层阻抗估算模型的应用和对比. 科学通报, 2013, 58(18): 1775–1783. |
| [18] | 王治海, 刘建栋, 刘玲, 邬定荣, 毕建杰. 几种气孔导度模型在华北地区适应性研究. 中国农业气象, 2012, 33(3): 412–416. DOI:10.3969/j.issn.1000-6362.2012.03.014 |
| [19] | Ji S, Tong L, Kang S, Li F, Lu H, Du T, Li S, Ding R. A modified optimal stomatal conductance model under water-stressed condition. International Journal of Plant Production, 2017, 11(2): 295–314. |
| [20] | Yu G R, Zhuang J, Yu Z L. An attempt to establish a synthetic model of photosynthesis-transpiration based on stomatal behavior for maize and soybean plants grown in field. Journal of Plant Physiology, 2001, 158(7): 861–874. DOI:10.1078/0176-1617-00177 |
| [21] | 高冠龙, 张小由, 鱼腾飞, 常建峰, 赵虹. 极端干旱条件下胡杨叶片气孔导度模拟. 干旱区地理, 2016, 39(3): 607–612. |
| [22] | 罗紫东, 关华德, 章新平, 刘娜, 张赐成, 王婷. 桂花树冠层气孔导度模型的优化及其参数分析. 生态学报, 2016, 36(13): 3995–4005. |
| [23] | 王玉辉, 周广胜. 羊草叶片气孔导度对环境因子的响应模拟. 植物生态学报, 2000, 24(6): 739–743. DOI:10.3321/j.issn:1005-264X.2000.06.017 |
| [24] | 周莉, 周广胜, 贾庆宇, 吕国红, 谢艳兵, 赵先丽. 盘锦湿地芦苇叶片气孔导度的模拟. 气象与环境学报, 2006, 22(4): 42–46. DOI:10.3969/j.issn.1673-503X.2006.04.008 |
| [25] | 高冠龙, 张小由, 常宗强, 鱼腾飞, 赵虹. 植物气孔导度的环境响应模拟及其尺度扩展. 生态学报, 2016, 36(6): 1491–1500. |
| [26] | Wang H L, Guan H D, Deng Z J, Simmons C T. Optimization of canopy conductance models from concurrent measurements of sap flow and stem water potential on Drooping Sheoak in South Australia. Water Resources Research, 2014, 50(7): 6154–6167. DOI:10.1002/wrcr.v50.7 |
| [27] | Damour G, Simonneau T, Cochard H, Urban L. An overview of models of stomatal conductance at the leaf level. Plant, Cell & Environment, 2010, 33(9): 1419–1438. |
| [28] | Anjum S A, Wang L C, Farooq M, Hussain M, Xue L L, Zhou C M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. Journal of Agronomy and Crop Science, 2011, 197(3): 177–185. DOI:10.1111/jac.2011.197.issue-3 |
| [29] | 赵锦, 杨晓光, 刘志娟, 吕硕, 王静, 陈阜. 全球气候变暖对中国种植制度的可能影响X.气候变化对东北三省春玉米气候适宜性的影响. 中国农业科学, 2014, 47(16): 3143–3156. DOI:10.3864/j.issn.0578-1752.2014.16.003 |
| [30] | 范嘉智, 王丹, 胡亚林, 景盼盼, 王朋朋, 陈吉泉. 最优气孔行为理论和气孔导度模拟. 植物生态学报, 2016, 40(6): 631–642. |
| [31] | 米娜, 蔡福, 张玉书, 纪瑞鹏, 于文颖, 张淑杰, 方缘. 不同生育期持续干旱对玉米的影响及其与减产率的定量关系. 应用生态学报, 2017, 28(5): 1563–1570. |
| [32] | Hofstra G, Hesketh J D. The effect of temperature on stomatal aperture in different species. Canadian Journal of Botany, 1969, 47(8): 1307–1310. DOI:10.1139/b69-184 |
| [33] | Leuning R. Modelling stomatal behaviour and and photosynthesis of Eucalyptus grandis. Australian Journal of Plant Physiology, 1990, 17(2): 159–175. DOI:10.1071/PP9900159 |
| [34] | Brooks A, Farquhar G D. Effect of temperature on the CO2/O2 specificity of ribulose-1, 5-bisphosphate carboxylase/oxygenase and the rate of respiration in the light:estimates from gas-exchange measurements on spinach. Planta, 1985, 165(3): 397–406. DOI:10.1007/BF00392238 |
| [35] | Cowan I R, Farquhar G D. Stomatal function in relation to leaf metabolism and environment. Symposia of the Society for Experimental Biology, 1977, 31: 471–505. |
| [36] | Dolman A J. A multiple-source land surface energy balance model for use in general circulation models. Agricultural and Forest Meteorology, 1993, 65(1/2): 21–45. |
| [37] | Wang Y P, Leuning R. A two-leaf model for canopy conductance, photosynthesis and partitioning of available energy I::model description and comparison with a multi-layered model. Agricultural and Forest Meteorology, 1998, 91(1/2): 89–111. |
| [38] | Pantin F, Monnet F, Jannaud D, Costa J M, Renaud J, Muller B, Simonneau T, Genty B. The dual effect of abscisic acid on stomata. New Phytologist, 2013, 197(1): 65–72. DOI:10.1111/nph.12013 |
| [39] | Ran L M, Pleim J, Song C H, Walker J T, Binkowski F S. A photosynthesis-based two-leaf canopy stomatal conductance model for meteorology and air quality modeling with WRF/CMAQ PX LSM. Journal of Geophysical Research, 2017, 122(3): 1930–1952. |
| [40] | Janka E, Körner O, Rosenqvist E, Ottosen C O. A coupled model of leaf photosynthesis, stomatal conductance, and leaf energy balance for chrysanthemum (Dendranthema grandiflora). Computers and Electronics in Agriculture, 2016, 123: 264–274. DOI:10.1016/j.compag.2016.02.022 |
| [41] | Baldocchi D. A lagrangian random-walk model for simulating water vapor, CO2 and sensible heat flux densities and scalar profiles over and within a soybean canopy. Boundary-Layer Meteorology, 1992, 61(1/2): 113–144. |
| [42] | Zhang N Y, Li G, Yu S X, An D S, Sun Q, Luo W H, Yin X Y. Can the responses of photosynthesis and stomatal conductance to water and nitrogen stress combinations be modeled using a single set of parameters?. Frontiers in Plant Science, 2017, 8: 328. |
| [43] | Miner G L, Bauerle W L, Baldocchi D D. Estimating the sensitivity of stomatal conductance to photosynthesis:a review. Plant, Cell & Environment, 2017, 40(7): 1214–1238. |