 2018, Vol. 38
2018, Vol. 38文章信息
- 许宇田, 童春富.
- XU Yutian, TONG Chunfu.
- 长江口九段沙湿地海三棱藨草生物量分配特征及其影响因子
- Biomass allocation characteristics of Scirpus mariqueter and corresponding influencing factors in Jiuduansha Shoals of the Yangtze Estuary
- 生态学报. 2018, 38(19): 7034-7044
- Acta Ecologica Sinica. 2018, 38(19): 7034-7044
- http://dx.doi.org/10.5846/stxb201710241903
-
文章历史
- 收稿日期: 2017-10-24
- 网络出版日期: 2018-06-20

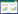




克隆植物又称无性系植物, 它们将资源分配于各构件时会进行权衡, 从而使生长和繁殖的效益达到最佳[1-3]。这种资源分配通过植物生物量可以体现出来。生物量是植物的基本生物学特征和功能性状, 是物质和能量积累的基本体现; 生物量分配是决定其资源获取、生长竞争以及繁殖能力的重要因素, 对植物生态策略的形成至关重要[4-5]。湿地是水陆交替的过渡带, 是地球上重要的生态系统类型之一。生活于其中的湿地植物相当一部分具有克隆生长特征。它们通过功能性状的可塑性来调节资源配置, 选择不同的生物量分配模式以适应复杂多变的环境, 从而最大效率地利用资源来提高其生存适合度和与竞争能力[6]。湿地克隆植物生物量分配已成为湿地生态学研究中的一大关键性问题[7-8]。
海三棱藨草(Scirpus mariqueter)为我国特有种, 主要分布在长江口和杭州湾一带的潮间带盐沼湿地[9]。它是莎草科藨草属的一种多年生耐盐草本植物, 生长期为4—11月, 花期为5—6月; 可进行有性繁殖或营养繁殖, 常形成单优植物群落; 具有促淤保滩、净化水质, 以及生物栖息地等多种生态功能[10-12]。目前针对海三棱藨草的研究已涉及种群物候与分布格局、繁殖生态学、种间竞争能力、生态价值与功能、生物量动态与分配、植物生理等诸多领域[10, 13-22]。海三棱藨草生物量分配的相关研究表明, 不同环境和干扰条件下海三棱藨草在生物量分配上表现出很强的生态适应性, 密度等形态参数和高程梯度、沉积物有机碳含量等环境指标在海三棱藨草的生物量分配中扮演着非常重要的角色[15-16, 18, 23-24]。已有的研究分别在这两个方面进行了有益的探索, 为进一步开展相关研究提供了良好的基础。本文参考已有研究, 在现场取样调研的基础上, 以海三棱藨草生物量分配特征为切入点, 综合植物形态特征与环境因子, 展开系统分析, 探究影响海三棱藨草生物量分配的主要因子及其作用机理。研究结果可以为海三棱藨草的培育与恢复, 盐沼湿地生态系统的保护与合理利用提供科学依据。
1 材料与方法 1.1 研究区域研究区域位于长江口九段沙湿地国家级自然保护区。九段沙为长江河口新生沙洲, 位于南北槽之间, 主要由江亚南沙、上沙、中沙、下沙及邻近浅水水域组成。九段沙土壤发育过程短, 潮滩沉积物以淤泥和粉砂为主。该区域属于亚热带季风性气候区, 年降水量约1143 mm, 年平均气温为15.7℃[25]。附近水域潮汐属于非正规半日浅海潮, 平均潮差2.67 m, 最大潮差4.62 m; 冬季盐度被高盐度咸水控制, 约为3‰—15‰, 夏季主要受淡水控制, 约为1‰—6‰; 在长江来水来沙与海潮的共同作用下, 区域水动力条件复杂[26]。近年来, 九段沙湿地处在长江口高强度开发的大环境中, 在自然干扰和人为干扰下, 平面扩张减慢, 水下滩淤涨受到抑制, 中高滩淤涨速度加快, 呈现“长高不长大”的变化特点[27]。沙洲整体上北淤南冲, 淤积区域集中在上、中、下沙中高滩区、江亚南沙与上沙沙尾, 以及靠近北槽深水航道南导堤两侧; 冲刷区域主要为九段沙南侧区域[28-29]。沙洲的冲淤动态与盐沼湿地植物群落的发育与演替密切相关。
九段沙盐沼植被具有明显的成带分布特征, 植物种类少, 群落结构简单, 常形成单优群落, 在群落交错带出现分布范围很窄的混生群落。原有自然植被优势种为芦苇(Phragmites australis)、海三棱藨草和藨草(S.triqueter)。1997年在中沙区域引种了互花米草(Spartina alterniflora), 目前已扩散至中沙和下沙大部分区域, 形成了大面积的单优群落。海三棱藨草主要分布在植被带前缘, 为重要的先锋物种, 受波浪与潮汐作用较大。在严重冲刷岸段基本没有海三棱藨草分布, 往往出现侵蚀陡坎, 从光滩直接过渡到芦苇、互花米草分布区。
1.2 研究方法 1.2.1 样品采集海三棱藨草的生物量分配随季节变化[15], 但其在10月份左右开始停止生长、结籽, 并开始衰老, 生物量达到最大[17], 地下生物量在植株开始衰老后相对稳定[14], 可以进行生物量分配的分析与研究。因此作者于2016年10月在九段沙开展了现场取样调研工作。海三棱藨草群落是该区域的特征植被, 根据文献资料以及现场观测, 遵循典型性、代表性和一致性的原则, 分别在九段沙各沙洲选择群落发育典型、空间分布均匀、原生状态完好的研究区, 利用GPS确定采样站点。采样站点设置如图 1所示。J1—J3为3个取样站点, 分别为江亚南沙(J1)、上沙(J2)、中下沙(J3)。主要采样区基本为淤涨区。

|
| 图 1 九段沙采样站点图 Fig. 1 Location of sampling sites in Jiuduansha Shoals |
每站点设置5块样地, 每样地设置2个植被取样样方, 样方面积为50 cm×50 cm。现场记录各样方海三棱藨草的盖度、株数、株高, 齐地刈割采集植物地上部分。在每个样方内, 设置25 cm×25 cm小样方, 用铁铲向下挖掘至约25—30 cm, 确保将样方内植株地下部分全部挖出。挖出的泥样用0.5 mm孔径网筛淘洗, 得到海三棱藨草地下部分。
沉积物样品与植物样品采集同步进行。在每个植物样方内, 使用内径3.5 cm的柱状取样器, 采集深度30 cm的沉积物柱样, 现场分割成6段5 cm长的泥柱。现场用便携式仪器测定记录沉积物氧化还原电位、pH、温度, 孔隙水的盐度等指标。每个指标在每个样方中重复测定2次。
1.2.2 样品处理(1) 海三棱藨草
在实验室内, 将植物样品用自来水冲洗多遍后, 再用Mill-Q水冲洗。按照茎叶、果实分离地上部分, 并记录开花株数、结籽数; 按照须根、根茎、球茎、茎基(茎叶埋在土面以下部分)分离地下部分, 并测量记录茎基长与根茎长。分离后的各部分在60℃下烘干至恒重, 称量得到生物量干重, 精确至0.01 g。
(2)沉积物
沉积物柱样在实验室称取湿重后, 于60℃烘干至恒重, 称取干重, 精确至0.01 g, 用以计算含水率、容积密度。烘干的土壤样品分别用马尔文激光粒度仪(MS2000)测粒度, 用元素分析仪(Vario EL III)测定总碳、总氮含量。
1.2.3 数据处理计算植株密度、平均株高和茎基(截留茎)高度, 将其作为海三棱藨草整体表形参数。计算得到根茎、果实、球茎各部分生物量比例分别表征克隆生殖投资、有性生殖投资以及生长投资; 计算果实与根茎重之比、果实与球茎重之比以及根茎与球茎重之比来表征有性和无性繁殖之间的对比关系[15]。
采用统计软件包IBM SPSS Statistics 22进行数据处理。采用单因素方差分析(one-way ANOVA)对不同站点间生境因子、海三棱藨草的相关参数进行显著性分析, 在此基础上, 以boferroni检验方法进一步做组间两两比较, 分析不同站点两两之间的差异。以P<0.05表示显著差异, P<0.01表示极显著差异。采用统计软件Primer 5.2.8对海三棱藨草生物量分配特征与环境变量进行非线性回归分析。用BVSTEP程序找出与海三棱藨草生物量分配特征匹配最好的生境因子组合, 并计算相关系数; 再用RELATE程序检验两者之间的相关性是否显著。所有数据在使用时需经过平方根变换, 以满足数据的正态性要求; 生境因子的相关数据需要进行标准化处理, 以避免量纲差异造成的影响[30]。
2 结果与分析 2.1 海三棱藨草表形特征不同站点海三棱藨草表形特征如图 2所示。单因素方差分析结果表明, 植株密度和植株高度在不同站点间存在显著差异(P<0.05), 而茎基高度并未发现显著不同(P>0.05)。不同站点两两比较的结果表明, 植株密度在J3站点显著高于J2站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);植株高度在J1站点显著高于J3站点(P<0.05), J2介于J1、J3之间, 与两者差异均不显著(P>0.05);茎基高度在各站点两两之间均不存在显著差异(P>0.05)。

|
| 图 2 不同站点海三棱藨草表形特征(平均值±标准误) Fig. 2 Morphological characteristics of S. mariqueter in the different sites (Mean ± SE) 上方标注不同小写字母的柱子表示相互之间存在显著差异(P<0.05) |
不同站点海三棱藨草各部分的生物量分配如图 3所示。单因素方差分析结果表明, 海三棱藨草的茎叶、果实、须根、根茎生物量所占比例在不同站点间存在显著差异(P<0.05), 而球茎生物量所占比例并未发现显著不同(P>0.05)。不同站点两两比较的结果表明, 茎叶生物量所占的比例在J2站点显著高于J3站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);果实生物量所占的比例在J1站点显著高于J2和J3站点(P<0.05), J2和J3之间差异并不显著(P>0.05);须根生物量所占的比例在J3站点显著高于J2站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);根茎生物量所占的比例在J3站点显著高于J2站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);球茎生物量所占比例在不同站点两两之间并未发现显著不同(P>0.05)。

|
| 图 3 不同站点海三棱藨草生物量分配特征(平均值±标准误) Fig. 3 omass allocation characteristics of S. mariqueter in the different sites (Mean± SE) 上方标注不同小写字母的柱状图表示同一类别生物量在不同站点间存在显著差异(P<0.05);J1—J3为3个取样站点, 其中, J1:江亚南沙, Jiuduansha Shoals 1;J2:上沙, Jiuduansha Shoals 2;J3:下沙, Jiuduansha Shoals 3 |
不同站点海三棱藨草繁殖与生长之间的对比关系如图 4所示。单因素方差分析结果表明, 果实与根茎之比、果实与球茎之比在不同站点间差异显著(P<0.05), 而根茎与球茎之比在不同站点间并未发现显著差异(P>0.05)。不同站点两两比较的结果表明, 果实与根茎之比在J1站点显著高于J2和J3站点(P<0.05), J2和J3之间并无显著差异(P>0.05);果实与球茎之比在J1站点显著高于J3站点(P<0.05), J2介于两者之间, 与J1、J3均无显著差异(P>0.05)。

|
| 图 4 不同站点海三棱藨草繁殖与生长特征(平均值±标准误) Fig. 4 Characteristics of reproduction and growth of S.mariqueter in the different sites(Mean± SE) 上方标注不同小写字母的柱状图表示同一类别生物量比率在不同站点间存在显著差异(P<0.05);FRR:果实/根茎比率, Ratio of fruit to rhizome biomass; FCR:果实/球茎比率, Ratio of fruit to corm biomass; RCR:根茎/球茎比率, Ratio of rhizome to corm biomass |
海三棱藨草不同参数相关特征如表 1所示。植株密度与根茎、须根生物量比例呈极显著正相关(P<0.01), 而与茎基高度显著负相关(P<0.05)。茎基高度与根茎生物量比例呈极显著负相关(P<0.01), 与须根生物量比例显著负相关(P<0.05), 与茎叶生物量比例显著正相关(P<0.05)。茎叶生物量比例与球茎、根茎、须根生物量比例均呈现极显著负相关关系(P<0.01)。须根生物量比例与球茎、根茎生物量比例呈现极显著正相关关系(P<0.01);同时球茎与根茎生物量比例也呈现极显著正相关关系(P<0.01)。
| 植物生长特征 Plant growth characteristics |
球茎生物量比例 Corm biomass allocation |
根茎生物量比例 Rhizome biomass allocation |
须根生物量比例 Root biomass allocation |
果实生物量比例 Fruit biomass allocation |
茎叶生物量比例 Shoot biomass allocation |
茎基高度 Stembase height |
植株高度 Shoot height |
| 植株密度Density | 0.447 | 0.653** | 0.702** | -0.251 | -0.489 | -0.638* | -0.359 |
| 植株高度Shoot height | -0.128 | -0.268 | -0.129 | 0.505 | 0.019 | 0.282 | |
| 茎基高度 Stembase height |
-0.498 | -0.761** | -0.626* | 0.284 | 0.526* | ||
| 茎叶生物量比例 Shoot biomass allocation |
-0.955** | -0.701** | -0.905** | -0.345 | |||
| 果实生物量比例 Fruit biomass allocation |
0.199 | -0.252 | 0.122 | ||||
| 须根生物量比例 Root biomass allocation |
0.836** | 0.716** | |||||
| 根茎生物量比例 Rhizome biomass allocation |
0.683** | ||||||
| **, P<0.01;*, P<0.05 | |||||||
不同站点沉积物理化性质如图 5所示。单因素方差分析结果表明, 不同站点的孔隙水盐度、沉积物电导率、沉积物的容积密度(5—10、25—30 cm土层)、总碳含量(10—15、20—25、25—30 cm土层)、总氮含量(5—10 cm土层)存在显著差异(P<0.05), 而pH、氧化还原电位以及同一深度的含水率、其他深度的柱段的容积密度以及总碳总氮含量在不同站点间并未发现显著不同(P>0.05)。不同站点两两比较的结果表明, 沉积物电导率在J1站点显著低于J2和J3(P<0.05), J2和J3两者之间并无显著差异(P>0.05);孔隙水盐度在J3站点显著高于J1和J2站点(P<0.05), J2低于J1, 但两者之间并无显著差异(P>0.05)。深度5—10 cm沉积物容积密度在J2站点显著高于J3站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);深度25—30 cm沉积物容积密度在J2站点显著低于J1和J3站点(P<0.05), 而J1和J3则不存在显著差异(P>0.05)。10—15 cm的碳百分含量在J1站点显著高于J3站点(P<0.05), J2介于两者之间, 与J1、J3均无显著差异(P>0.05);20—25 cm的碳百分含量在J2站点显著高于J3站点(P<0.05), J1介于两者之间, 与J2、J3均无显著差异(P>0.05);25—30 cm的碳百分含量在J3站点显著低于J1和J2站点(P<0.05), J1低于J2, 但是两者之间无显著差异(P>0.05)。5—10 cm的氮百分含量在J1站点显著高于J2和J3站点(P<0.05), J2和J3之间差异不显著(P>0.05)。

|
| 图 5 不同站点沉积物理化性质 Fig. 5 Physical and chemical characteristics of the sediment in the different sites 图中标注不同小写字母表示不同站点间的显著性差异(P<0.05) |
不同站点海三棱藨草生物量分配特征与沉积物理化指标、海三棱藨草表形参数非线性回归分析结果如表 2所示。海三棱藨草的果实与根茎的生物量之比和5—10、25—30 cm总碳含量, 0—5、5—10 cm总氮含量, 0—5 cm容积密度, 茎基高度等6种因子的组合极显著相关(P<0.01);而其他生物量分配指标与本文涉及到的环境因子、植物表形参数均不存在显著相关性(P>0.05)。
| 特征 Characteristics |
相关性系数 Correlation |
筛选因子组合 Selection combination |
P |
| 果实/球茎比Fruit/corm biomass ratio | 0.094 | 6, 8, 14, 19, 22 | 0.212 |
| 果实/根茎比Fruit/rhizome biomass ratio | 0.341 | 7, 11—13, 24, 32 | 0.006 |
| 根茎/球茎比Rhizome/corm biomass ratio | 0.172 | 2, 5 | 0.124 |
| 根茎生物量比例Rhizome biomass allocation | 0.129 | 6, 11—14, 16, 21, 32 | 0.151 |
| 果实生物量比例Fruit biomass allocation | -0.015 | 5, 12, 31 | 0.443 |
| 茎叶生物量比例Shoot biomass allocation | 0.172 | 1, 8, 14, 16, 22, 24, 26, 30 | 0.087 |
| 球茎生物量比例Corm biomass allocation | -0.157 | 2, 4, 8—10, 12, 15, 19, 26, 30 | 0.99 |
| 须根生物量比例Root biomass allocation | 0.165 | 3, 6, 7, 11—13, 18, 20, 21, 23, 26—30 | 0.138 |
| 因子: 1:沉积物孔隙水盐度; 2:沉积物电导率; 3:沉积物温度; 4:沉积物pH; 5:沉积物氧化还原电位; 6:0—5 cm沉积物总碳含量; 7: 5—10 cm沉积物总碳含量; 8: 10—15 cm沉积物总碳含量; 9: 15—20 cm沉积物总碳含量; 10: 20—25 cm沉积物总碳含量; 11: 25—30 cm沉积物总碳含量; 12: 0—5 cm沉积物总氮含量; 13: 5—10 cm沉积物总氮含量; 14: 10—15 cm沉积物总氮含量; 15: 15—20 cm沉积物总氮含量; 16: 20—25 cm沉积物总氮含量; 17: 25—30 cm沉积物总氮含量; 18: 0—5 cm沉积物含水率; 19: 5—10 cm沉积物含水率; 20: 10—15 cm沉积物含水率; 21: 15—20 cm沉积物含水率; 22: 20—25 cm沉积物含水率; 23: 25—30 cm沉积物含水率; 24: 0—5 cm沉积物容积密度; 25: 5—10 cm沉积物容积密度; 26: 10—15 cm沉积物容积密度; 27: 15—20 cm沉积物容积密度; 28: 20—25 cm沉积物容积密度; 29: 25—30 cm沉积物容积密度; 30:海三棱藨草植株密度; 31:海三棱藨草株高; 32:海三棱藨草茎基高度 | |||
已有研究表明, 具有克隆生长特征的植物的生物量分配受植株高度和密度等自身生长特征的影响[3, 31-34]。植物种内存在对有限空间与资源的竞争, 生物量分配格局是决定植物资源获取、竞争以及繁殖能力的重要因素[35]。植物自身生长情况如密度变化等会影响种内竞争强度, 使得植物生物量分配发生变化[20, 34, 36]。克隆植物生物量分配普遍支持最优化理论, 即通过对器官间生物量分配的权衡, 对资源竞争作出反应, 以保证植物能最大化利用受限资源, 应对不断变化的环境条件[6, 32, 37]。
目前针对海三棱藨草生物量分配的研究中, 有结果显示随着植株高度增加, 海三棱藨草的营养投资减少, 而分配更多的资源给有性繁殖[15]。但是本次研究并未发现海三棱藨草植株高度与各部分生物量间存在显著相关性。同时, 本次研究显示, 植株密度和根茎以及须根生物量比例呈极显著正相关, 这在已有研究的基础上进一步证明了密度是影响克隆植物生物量分配的一个重要因子[20, 36]。根茎生物量比例可以反映出植物对无性繁殖的投资, 因此由本文研究结果可知, 随密度的增加, 海三棱藨草对无性繁殖的投资增加, 这与之前的部分研究结果相一致[18, 38]。但目前关于繁殖投资对密度的响应的研究, 也存在不同的结论, 如有研究表明在生物因素(种间、种内竞争等)的干扰下, 海三棱藨草主要采取有性繁殖的策略来维持适合度[15, 39], 因为种子传播可使植物避开不利场所, 而增加对无性繁殖的资源分配可能会增加其种间竞争强度[1]。之所以出现这种情况, 可能与本文研究中海三棱藨草的地下器官所需资源受限的程度有关。增加无性繁殖投资这一生物量分配模式可以被认为是克隆植物的保守策略。由于海三棱藨草对土壤营养的需求较低[40], 并且通过无性系分株比通过种子形成幼苗的方式成活率更高, 无性繁殖形成聚集的群体利用不同小生境中克隆分株的形态可塑性和克隆生理整合作用[41-42], 可以最大限度地吸收和利用环境资源, 缓解环境中资源不均或干扰胁迫对生长和繁殖带来的不利影响, 提高个体的生存和竞争力。海三棱藨草生物量分配符合最优化理论, 即随着密度增加, 生存空间受到限制, 种内竞争加剧, 海三棱藨草通过权衡生物量分配来实现对矿质营养和水分等资源与环境的最大化利用, 提高对生境的适合度。
3.2 环境因子对海三棱藨草生物量分配的影响根据已有研究, 除自身生长因子之外, 具有克隆生长特性的盐沼植物生物量分配还受到环境因子的影响[43-45]。如氮素可通过改变氮在叶片中的分配格局影响叶片的光合作用, 并通过改变碳水化合物的库源关系和能量消耗水平而调节碳同化物质在体内的分配, 从而影响其生长和生物量分配[46-49]; 土壤的含水率、透气性等对克隆植物的生物量积累与分配等均有显著影响[50-52]; 温度变化通过直接改变植物的光合能力、生长速率, 间接改变土壤含水率、营养物质的可利用性, 影响植物生长以及生物量分配[53-54]。不同区域的环境因子往往存在明显、甚至显著的差异, 这可能是导致海三棱藨草具有不同的生物量分配特征的关键所在。本文研究结果显示, 特定深度的沉积物总碳、总氮含量、容积密度以及茎基高度等因子组合与部分海三棱藨草生物量分配指标呈极显著相关关系。结合已有研究, 进一步证实了土壤营养元素(碳氮含量、总磷含量等)对海三棱藨草生物量及生物量分配存在重要作用[20, 23, 39, 55]。尽管有控制实验显示海三棱藨草须根、茎叶和果实生物量分配受到盐度和水位的显著影响[56], 然而本文研究中并未发现海三棱藨草生物量分配与沉积物盐度[57-59]、含水率[40]以及温度[60]等环境因子存在显著相关性。这可能是由于环境因子的变化在实验控制条件下与自然状态下存在差异, 并且环境因子间还存在着相互作用, 可能与高程[15, 20, 24]、潮汐水动力[61-62]等其他因子的影响有关。未来需要涵盖更多的环境因子, 进行综合分析。
本文研究中的茎基高度可以反映出环境中泥沙堆积深度。茎基高度越大, 植物根茎埋藏越深, 海三棱藨草受泥沙堆积的影响越大。从本文研究结果来看, 茎基高度不仅是非线性回归筛选出的与果实根茎之比相关的因子之一, 而且与根茎生物量比例呈极显著负相关, 与植株密度、须根生物量比例显著负相关, 与茎叶生物量分配显著正相关。与之前对盐沼克隆植物短尖苔草(Carex brevicuspis)的研究相比[63], 沉积物堆积产生的影响在植株密度变化上较为一致, 对其他指标的影响却明显不同。根据本文研究结果推测, 随着沉积物堆积加厚, 沉积压力可能会对植株地下部分, 包括根尖分生组织形成物理胁迫, 不利于地下器官的生长发育, 海三棱藨草会倾向于将更多的资源分配到茎叶, 为有性繁殖提供条件, 减少无性繁殖投资比例。这与已有研究认为, 在生物量分配的权衡中, 克隆植物在非生物因素的压力下更倾向于采取无性繁殖的方式[39]的观点存在明显不同, 即如果非生物因素直接影响或限制了无性繁殖, 克隆植物同样会选择有性繁殖策略。
综上所述, 海三棱藨草生物量分配特征是其自身生长特征与环境因子综合作用的结果。从本文研究结果来看, 除了本文涉及的环境因子外, 可能还存在其他因子也会对海三棱藨草的生物量及分配特征产生影响。在今后的研究中需要拓展生境因子涵盖范围, 从而进一步系统、综合研究多因子对于海三棱藨草生物量分配特征的影响。
| [1] | 张玉芬, 张大勇. 克隆植物的无性与有性繁殖对策. 植物生态学报, 2006, 30(1): 174–183. DOI:10.3321/j.issn:1005-264X.2006.01.024 |
| [2] | Wildová R, Gough L, Herben T, Hershock C, Goldberg D E. Architectural and growth traits differ in effects on performance of clonal plants:an analysis using a field-parameterized simulation model. Oikos, 2007, 116(5): 836–852. DOI:10.1111/oik.2007.116.issue-5 |
| [3] | Ikegami M, Whigham D F, Werger M J A. Effects of local density of clonal plants on their sexual and vegetative propagation strategies in a lattice structure model. Ecological Modelling, 2012, 234(3): 51–59. |
| [4] | 王沫竹, 董必成, 李红丽, 于飞海. 根茎型植物扁秆荆三棱对光照强度和养分水平的生长响应及资源分配策略. 生态学报, 2016, 36(24): 8091–8101. |
| [5] | Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4): 207–215. DOI:10.1078/1433-8319-00083 |
| [6] | Xie X F, Hu Y K, Pan X, Liu F H, Song Y B, Dong M. Biomass allocation of stoloniferous and rhizomatous plant in response to resource availability:a phylogenetic meta-analysis. Frontiers in Plant Science, 2016, 7: 603. |
| [7] | Martin C W, Valentine J F. Sexual and asexual reproductive strategies of invasive Eurasian milfoil (Myriophyllum spicatum) in estuarine environments. Hydrobiologia, 2014, 727(1): 177–184. DOI:10.1007/s10750-013-1798-9 |
| [8] | 马俊改, 石福臣. 养分条件对互花米草表型可塑性的影响. 生态学杂志, 2011, 30(3): 459–463. |
| [9] | 袁兴中, 陆健健, 刘红. 河口盐沼植物对大型底栖动物群落的影响. 生态学报, 2002, 22(3): 326–333. DOI:10.3321/j.issn:1000-0933.2002.03.006 |
| [10] | 陈中义. 长江口海三棱藨草的生态价值及利用与保护. 河南科技大学学报:自然科学版, 2005, 26(2): 64–67. |
| [11] | Ge Z M, Zhou X, Wang T H, Wang K Y, Pei E L, Yuan X. Effects of vegetative cover changes on the carrying capacity of migratory shorebirds in a newly formed wetland, Yangtze River estuary, China. Zoological Studies, 2009, 48(6): 769–779. |
| [12] | 王淑琼, 王瀚强, 方燕, 李恺. 崇明岛滨海湿地植物群落固碳能力. 生态学杂志, 2014, 33(4): 915–921. |
| [13] | 张利权, 雍学葵. 海三棱藨草种群的物候与分布格局研究. 植物生态学与地植物学学报, 1992, 16(1): 43–51. |
| [14] | 雍学葵, 张利权. 海三棱藨草种群的繁殖生态学研究. 华东师范大学学报:自然科学版, 1992(4): 94–99. |
| [15] | Sun S C, Gao X M, Cai Y L. Variations in sexual and asexual reproduction of Scirpus mariqueter along an elevational gradient. Ecological Research, 2001, 16(2): 263–274. DOI:10.1046/j.1440-1703.2001.00395.x |
| [16] | Sun S C, Cai Y L, An S Q. Differences in morphology and biomass allocation of Scirpus mariqueter between creekside and inland communities in the Changjiang estuary, China. Wetlands, 2002, 22(4): 786–793. DOI:10.1672/0277-5212(2002)022[0786:DIMABA]2.0.CO;2 |
| [17] | 梅雪英, 张修峰. 长江口湿地海三棱蔗草(Scirpus mariqueter)的储碳、固碳功能研究——以崇明东滩为例. 农业环境科学学报, 2007, 26(1): 360–363. DOI:10.3321/j.issn:1672-2043.2007.01.073 |
| [18] | 王亮, 李静, 杨娟, 蔡永立. 崇明东滩海三棱藨草生殖对策探讨. 信阳师范学院学报:自然科学版, 2008, 21(4): 539–542. |
| [19] | 詹跃武, 何池全, 孟昭寿, 雷雁茹, 郑虹. 海三棱藨草盐沼土壤环境因子的时空分布特征. 海洋环境科学, 2011, 30(6): 765–770. DOI:10.3969/j.issn.1007-6336.2011.06.002 |
| [20] | 陶燕东, 于克锋, 何培民, 孙彬, 历成伟, 赵萌, 方淑波. 围垦后南汇东滩海三棱藨草的空间分布及其影响因子研究. 长江流域资源与环境, 2017, 26(7): 1032–1041. DOI:10.11870/cjlyzyyhj201707009 |
| [21] | 陈怀璞, 张天雨, 葛振鸣, 张利权. 崇明东滩盐沼湿地土壤碳氮储量分布特征. 生态与农村环境学报, 2017, 33(3): 242–251. |
| [22] | Dong L, Li X, Liu X, He K, Jiang X. Determining the effects of major cations (K+, Na+, Ca2+, Mg2+) and pH on Scirpus mariqueter to assess the heavy metal biotoxicity of a tidal flat ecosystem. Journal of Coastal Research, 2017, 33(5): 1086–1094. |
| [23] | 陈华, 王东启, 陈振楼, 杨红霞, 王军, 许世远. 崇明东滩海三棱藨草生长期沉积物有机碳含量变化. 环境科学学报, 2007, 27(1): 135–142. DOI:10.3321/j.issn:0253-2468.2007.01.025 |
| [24] | 孙书存, 蔡永立, 刘红. 长江口盐沼海三棱藨草在高程梯度上的生物量分配(英文). 植物学报, 2001, 43(2): 178–185. DOI:10.3321/j.issn:1672-9072.2001.02.011 |
| [25] | 胡颖慧, 贾建伟, 王磊, 唐玉姝, 郑广宏, 胡山, 孙瑛. 长江口九段沙湿地土壤石油类污染评价及源特征分析. 土壤通报, 2011, 42(1): 195–201. |
| [26] | 刘钰. 九段沙植被分布区碳汇功能评估[D]. 上海: 华东师范大学, 2013. |
| [27] | 张田雷. 大型涉水工程对九段沙湿地的冲淤效应研究[D]. 上海: 华东师范大学, 2011. |
| [28] | 陈炜, 李九发, 蒋陈娟, 李占海, 姚弘毅, 徐敏. 长江河口九段沙近期冲淤演变过程研究. 泥沙研究, 2011(1): 15–21. |
| [29] | 茅志昌, 张田雷, 郭建强. 九段沙尾浅滩冲淤演变对长江入海沙量变化的响应. 泥沙研究, 2014(4): 55–59. |
| [30] | Tong C F, Baustian J J, Graham S A, Mendelssohn I A. Salt marsh restoration with sediment-slurry application:effects on benthic macroinvertebrates and associated soil-plant variables. Ecological Engineering, 2013, 51(2): 151–160. |
| [31] | 李丽纯, 林瑞坤, 吴振海, 陈家金, 杨凯, 徐宗焕. 闽江口湿地互花米草地上生物量的月动态特征. 资源科学, 2009, 31(11): 1967–1972. DOI:10.3321/j.issn:1007-7588.2009.11.019 |
| [32] | 黎磊, 周道玮, 盛连喜. 植物种群自疏过程中构件生物量与密度的关系. 生态学报, 2012, 32(13): 3987–3997. |
| [33] | 钟青龙, 戴文龙, 项世亮, 王天厚, 汪承焕. 密度对三种莎草科植物克隆生长的影响. 生态科学, 2016, 35(1): 1–9. DOI:10.3969/j.issn.1671-6361.2016.01.001 |
| [34] | 范高华, 崔桢, 张金伟, 黄迎新, 神祥金, 赵学勇. 密度对尖头叶藜生物量分配格局及异速生长的影响. 生态学报, 2017, 37(15): 5080–5090. |
| [35] | Wang T H, Zhou D W, Wang P, Zhang H X. Size-dependent reproductive effort in Amaranthus retroflexus:the influence of planting density and sowing date. Canadian Journal of Botany, 2006, 84(3): 485–492. DOI:10.1139/b06-011 |
| [36] | Van Kleunen M, Fischer M, Schmid B. Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant. Oikos, 2001, 94(3): 515–524. DOI:10.1034/j.1600-0706.2001.940313.x |
| [37] | Bloom A J, Chapin Ⅲ F S, Mooney H A. Resource limitation in plants-an economic analogy. Annual Review of Ecology and Systematics, 1985, 16(1): 363–392. DOI:10.1146/annurev.es.16.110185.002051 |
| [38] | Loehle C. Partitioning of reproductive effort in clonal plants:a benefit-cost model. Oikos, 1987, 49(2): 199–208. DOI:10.2307/3566027 |
| [39] | 何彦龙. 中低潮滩盐沼植被分异的形成机制研究[D]. 上海: 华东师范大学, 2014. |
| [40] | 汪承焕. 环境变异对崇明东滩优势盐沼植物生长、分布与种间竞争的影响[D]. 上海: 复旦大学, 2009. |
| [41] | 于飞海, 董鸣. 生态学研究热点克隆植物生态学. 植物杂志, 2001(3): 32–33. |
| [42] | Roiloa S R, Hutchings M J. The effects of physiological integration on biomass partitioning in plant modules:an experimental study with the stoloniferous herb Glechoma hederacea. Plant Ecology, 2013, 214(4): 521–530. DOI:10.1007/s11258-013-0186-x |
| [43] | 谭勇, 何东进, 游巍斌, 王韧, 蔡金标, 王鹏, 赵莉莉, 严锦钰, 肖石红. 闽东滨海湿地入侵种互花米草表型可塑性及生物量分配对潮位梯度的响应. 应用与环境生物学报, 2014, 20(5): 863–868. |
| [44] | 王丹, 张银龙, 庞博, 徐明喜, 苏莹莹. 苏州太湖湿地芦苇生物量与水深的动态特征研究. 环境污染与防治, 2010, 32(7): 49–54. DOI:10.3969/j.issn.1001-3865.2010.07.011 |
| [45] | 何军, 赵聪蛟, 清华, 甘琳, 安树青. 土壤水分条件对克隆植物互花米草表型可塑性的影响. 生态学报, 2009, 29(7): 3518–3524. DOI:10.3321/j.issn:1000-0933.2009.07.009 |
| [46] | 王琪, 徐程扬. 氮磷对植物光合作用及碳分配的影响. 山东林业科技, 2005(5): 59–62. DOI:10.3969/j.issn.1002-2724.2005.05.040 |
| [47] | 甘琳, 赵晖, 清华, 肖燕, 李敏, 胡秋香, 安树青. 氮、硫互作对克隆植物互花米草繁殖和生物量累积与分配的影响. 生态学报, 2011, 31(7): 1794–1802. |
| [48] | 陈静, 李玉霖, 崔夺, 毛伟, 赵学勇. 氮素及水分添加对科尔沁沙地4种优势植物地上生物量分配的影响. 中国沙漠, 2014, 34(3): 696–703. |
| [49] | Meziane D, Shipley B. Interacting components of interspecific relative growth rate:constancy and change under differing conditions of light and nutrient supply. Functional Ecology, 1999, 13(5): 611–622. DOI:10.1046/j.1365-2435.1999.00359.x |
| [50] | 覃盈盈, 蒋潇潇, 李峰, 梁士楚. 互花米草在不同生境中的形态可塑性与生物量分配. 海洋环境科学, 2009, 28(6): 657–659. DOI:10.3969/j.issn.1007-6336.2009.06.014 |
| [51] | 李红丽, 智颖飙, 雷光春, 赵磊, 安树青, 邓自发, 周长芳. 咸淡交替灌溉对克隆植物大米草生长繁殖和生物量分配的影响. 生态学报, 2010, 30(7): 1744–1750. |
| [52] | 李亚芳. 洞庭湖湿地克隆植物繁殖分配对水位的响应[D]. 长沙: 中南林业科技大学, 2015. |
| [53] | Klanderud K, Totland Ø. Simulated climate change altered dominance hierarchies and diversity of an alpine biodiversity hotspot. Ecology, 2005, 86(8): 2047–2054. DOI:10.1890/04-1563 |
| [54] | 石冰, 马金妍, 王开运, 巩晋楠, 张超, 刘为华. 崇明东滩围垦芦苇生长、繁殖和生物量分配对大气温度升高的响应. 长江流域资源与环境, 2010, 19(4): 383–388. |
| [55] | 张骞, 李秀珍, 张运清, 孙香丽, 徐艳, 闫中正. 长江口湿地氮施加对海三棱藨草生长及生理特性的影响. 草地学报, 2017, 25(1): 115–121. |
| [56] | Wang C H, Lu M, Yang B, Yang Q, Zhang X D, Hara T, Li B. Effects of environmental gradients on the performances of four dominant plants in a Chinese saltmarsh:implications for plant zonation. Ecological Research, 2010, 25(2): 347–358. DOI:10.1007/s11284-009-0662-x |
| [57] | 韩华. 崇明滩涂湿地不同水盐梯度下植物群落碳氮磷生态化学计量学特征[D]. 上海: 华东师范大学, 2014. |
| [58] | 闫芊. 崇明东滩湿地植被的生态演替[D]. 上海: 华东师范大学, 2006. |
| [59] | 倪刚. 外来种互花米草对海三棱藨草竞争的限制性因子研究[D]. 上海: 上海大学, 2013. |
| [60] | 严格. 崇明东滩湿地盐沼植被生物量及碳储量分布研究[D]. 上海: 华东师范大学, 2014. |
| [61] | Negrin V L, Pratolongo P D, De Villalobos A E, Botté S E, Marcovecchio J E. Biomass, decomposition and nutrient cycling in a SW Atlantic Sarcocornia perennis marsh. Journal of Sea Research, 2014, 97: 50–55. |
| [62] | 任丽娟, 王国祥, 仇乐, 毛志刚, 刘金娥. 江苏潮滩湿地不同生境互花米草形态与生物量分配特征. 生态与农村环境学报, 2010, 26(3): 220–226. DOI:10.3969/j.issn.1673-4831.2010.03.006 |
| [63] | Chen X S, Xie Y H, Deng Z M, Li F, Hou Z Y. A change from phalanx to guerrilla growth form is an effective strategy to acclimate to sedimentation in a wetland sedge species Carex brevicuspis (Cyperaceae). Flora-Morphology, Distribution, Functional Ecology of Plants, 2011, 206(4): 347–350. DOI:10.1016/j.flora.2010.07.006 |