 2018, Vol. 38
2018, Vol. 38文章信息
- 赵志远, 袁琳, 李伟, 朱晓泾, 潘家琳, 陈雅慧, 张利权.
- ZHAO Zhiyuan, YUAN Lin, LI Wei, ZHU Xiaojing, PAN Jialin, CHEN Yahui, ZHANG Liquan.
- 生境异质性及源株密度对互花米草入侵力的影响
- Effects of habitat heterogeneity and ortet density on the invasiveness of Spartina alterniflora
- 生态学报. 2018, 38(18): 6632-6641
- Acta Ecologica Sinica. 2018, 38(18): 6632-6641
- http://dx.doi.org/10.5846/stxb201803300702
-
文章历史
- 收稿日期: 2018-03-30
- 修订日期: 2018-07-02

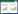




河口湿地位于海陆交界处[1], 在净化环境、提供生物栖息地、保护生物多样性、稳定海岸、维持区域生态平衡、调节碳循环等方面发挥着不可替代的重要作用[2-3], 是单位面积上生态服务价值最高的生态系统类型[4]。受陆海物质交汇、泥沙输移、冲淤演变、径流和潮汐相互作用的影响, 河口湿地具有独特的自然条件和异质生境[5], 沉积物粒径、盐度、氧化还原电位、水动力等与潮汐密切相关的系列环境因子都常常表现出高度的空间异质性[6]。有关河口湿地生境异质性与盐沼植物生长和分布的研究表明, 生境条件的异质分布大大增加了盐沼植物吸收和利用资源的难度, 严重影响其生长、繁殖和扩散[7-8]。
自然植物种群中普遍存在密度效应, 它对植物个体生长及种群动态的调节过程是动态的[9-10]。为了更好地适应不同生境类型, 克隆植物可通过密度调控维持个体及种群稳定, 即随着源株密度及种内竞争的改变, 植株存活率、克隆构型特征以及种群动态等方面均会产生变化[11-13]。王永健和钟章成[14]开展的蝴蝶花(Iris japonica)源株密度的控制实验表明, 不同源株密度对蝴蝶花生长、繁殖及生物量分配有显著影响。Worm和Beusch[15]的研究表明, 初始密度对鳗草(Zostera marina L.)幼苗生长和斑块面积扩大均有明显的正效应。黎磊等[10]研究也表明, 密度效应主要通过影响邻株植物间地上和地下器官的资源竞争强度来调节个体的资源配置。密度效应在河口湿地盐沼植物群落也较为明显, 例如, 随源株密度增加, 海三棱藨草(Scirpus mariqueter)和藨草(Scirpus triqueter)的单株平均生物量减少, 而藨草和糙叶薹草的总生物量增加, 三种莎草科植物存在明显的克隆变异, 表现出明显的密度依赖性及较强的形态可塑性[16]。
互花米草(Spartina alterniflora)隶属禾本科米草属(Spartina, 又名绳草属), 具有强大的克隆整合与环境饰变能力[17-18], 能在热带、亚热带到温带沿海地区宽广的气候带分布, 通常在河口、海湾等沿海滩涂的潮间带形成单优群落[19], 进而破坏生态系统结构和功能, 威胁生物多样性, 是典型的人为引种后扩散蔓延造成生态危害的外来入侵物种[20]。目前, 互花米草入侵机制的研究已广泛开展, 可概括为互花米草的成功入侵取决于互花米草的入侵力、合适的入侵途径以及受体系统的可入侵性[21]。互花米草极强的入侵力与其生物学特性密切相关, 包括对异质生境的广适性、多样的繁育对策、强大的扩散与竞争能力[17, 22-23]。目前, 已围绕淹水[23]、盐度[24]、潮滩冲淤[25]、温度[26]、土壤水分[27]等环境条件, 开展了互花米草对异质生境的适应性研究。然而, 有关互花米草的入侵力是否会受密度调控, 以及在生境异质性及源株密度共同影响下互花米草入侵力的变化尚有待深入研究。
长江河口湿地是典型的异质潮滩, 受流域、河口与海域的多重压力的影响, 河口湿地生态系统更具多变性、脆弱性与敏感性[28-30]。20世纪90年代, 互花米草被引种到长江口潮滩以积淤筑陆、保护河堤。经过定居适应阶段后, 由于其极强的扩散与竞争能力, 现已在潮间带湿地大肆蔓延, 对本地物种构成了严重威胁[31-32]。许多学者致力于长江口互花米草的入侵机制和防治策略的研究[7-8, 33], 但对于异质性河口环境中互花米草生物学特性(例如密度)影响其入侵力的研究尚不深入, 而互花米草入侵初始密度将可能决定其生长与资源分配的相对高低[14], 缺乏对这一过程的了解也不利于互花米草入侵动态分析和优化控制技术建立。因此, 本研究选择不同类型生境, 通过野外移栽实验, 模拟互花米草以不同初始密度入侵不同类型生境, 分析生境异质性及源株密度对互花米草定居、生长和扩散的影响, 深入探讨生境异质性及源株密度对互花米草入侵力的影响, 辨识影响互花米草入侵力的主要因子, 为我国开展互花米草防控及海岸带保护、修复和管理提供科学依据, 具有重要的理论和实践意义。
1 研究区域与研究方法 1.1 研究区域概况南汇东滩湿地(30°50'04"—31°06'47"N, 121°50'50"—121°51'40"E)位于长江河口南岸, 毗邻杭州湾(图 1), 是上海市潮滩淤长最快的湿地之一, 也是典型的异质潮滩[34]。该区域地处北亚热带, 年平均气温15—16℃, 年平均降水量1022 mm, 气候温暖湿润, 属典型的季风气候[35]。潮流属于非正规半日浅海潮流, 水流呈往复运动性质[36]。潮间带优势植物主要为芦苇(Phragmites australis)、海三棱藨草(S. mariqueter)和互花米草(S. alterniflora)。

|
| 图 1 研究区域(a)、移栽样方设置示意图(b)及移栽现场图(c) Fig. 1 Schematic diagram of the study area (a), transplanting area design (b) and transplanting scene (c) N1:北断面植被区, N2:北断面光滩区, S1:南断面植被区, S2:南断面光滩区 |
2016年4月, 在南汇东滩的自然潮滩设置南(粉砂质潮滩)、北(淤泥质潮滩)两条断面, 分别在每条断面的植被区及前沿光滩设置固定实验样方, 其中S1生境中优势物种为海三棱藨草, S2生境为粉砂质光滩, N1生境中优势物种为芦苇, N2生境为淤泥质光滩(图 1)。利用实时动态定位系统(Real-Time Kinematic Global Position System, RTK-GPS)测得S1、S2、N1和N2生境的吴淞高程分别为(2.7±0.12)、(2.3±0.21)、(3.0±0.29) m和(2.4±0.9) m, 均在互花米草可分布高程范围内[37]。4月10日, 在南汇东滩潮间带互花米草单优群落内, 挖取高度一致((18.28±1.65) cm)、长势健康的互花米草根茎苗, 并按1、3、5、10株/m2和20株/m2的源株密度随机移栽到固定样方内(1 m×1 m), 每个源株密度设3个重复, 共60个移栽样方, 样方与样方之间间隔2 m(图 1)。
1.2.2 植物监测于整个生长季(2016年4月10日—2016年10月10日)每月定期监测不同生境、不同源株密度处理下, 移栽互花米草根茎苗的存活率、密度及株高, 并计算种群增长率(公式1)。
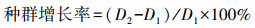
|
(1) |
式中, D1为源株密度, D2为实验结束时互花米草密度。
1.2.3 生境参数监测植物监测同期, 采用标志桩法[38]监测各生境潮滩冲淤动态。在每个生境的移栽样方旁设置一根冲淤标志桩, 每个生境15根, 共60根标志桩。标志桩长1.5 m, 垂直打入潮滩1 m左右以确保稳定, 记录标志桩暴露在滩面以上的高度(M0)作为相对高程零点, 之后每月测量标志桩顶端向东、南、西、北30 cm处, 距滩面垂直距离的平均值(Mx), M0与Mx的差值即为该时段的冲淤变化, 正值为淤积, 负值表示侵蚀[39]。
2016年4月(盐沼植物存活关键时期), 按照“S”型随机多点混合的采样方法, 使用100 m3容积的环刀在每个样方内采集3份重复表层沉积物样品, 带回实验室后利用贝克曼库尔特Coulter LS13320型激光粒度仪进行粒度分析; 利用W.E.T土壤三参仪(英国Delta-T)现场原位监测土壤电导率; 利用RTK-GPS测得样带滩面高程数据后, 通过与国家海洋局发布的平均潮位数据(芦潮港站)进行耦合后, 获得不同生境的日均淹水时间和相应的淹水深度。
1.3 数据处理应用统计软件(IBM SPSS Statistics 19)对实验数据进行统计分析, 使用单因子方差分析(One-way ANOVA)中的最小显著差异(LSD)对移栽后互花米草根茎苗的存活率、密度、株高及种群增长率进行差异显著性检验(显著性水平0.05);利用主成分分析方法对环境因子进行排序(旋转方法:最大方差法), 筛选出影响互花米草入侵力的关键环境因子。使用Origin 9.0软件作图。
2 结果 2.1 生境异质性及源株密度对互花米草存活的影响不同处理下, 互花米草存活率均表现为随着生长季的延长, 先下降后趋于平稳的变化规律(图 2), 生境异质性及源株密度对互花米草存活率均有极显著影响(P < 0.01, 表 1)。植被区(N1和S1)的互花米草存活率显著高于前沿光滩(N2和S2)(P < 0.05, 图 2)。源株密度对互花米草存活的影响在不同生境有不同的表现。互花米草在植被区的存活率随源株密度增加而减小(图 2)。其中, 在N1生境(芦苇群落), 互花米草源株密度为1株/m2和3株/m2时, 其存活率均为100%;随着源株密度的增加(5、10株/m2和20株/m2), 互花米草存活率略有下降, 但仍高于68%;在S1生境(海三棱藨草群落), 源株密度为1株/m2时, 互花米草存活率为100%;随着源株密度增加(3、5、10株/m2和20株/m2), 互花米草存活率略有下降但均维持较高水平(>70%)。在光滩区(N2和S2), 除了源株密度为20株/m2的互花米草能够一直存活在S2生境(粉砂质潮滩), 且能保留至生长季末期外, 其余源株密度的互花米草根茎苗即使最初存活, 至生长季末期也未能实现长期定居(图 2)。

|
| 图 2 异质生境及不同源株密度影响下互花米草的存活动态 Fig. 2 Survival dynamics of S.alterniflora under different ortet density in different habitats |
| 变异来源 Source of variation |
存活率 Survival rate |
密度 Density |
种群增长率 Population growth rate |
株高 Height |
|||||||
| F | P | F | P | F | P | F | P | ||||
| 生境Habitat | 784.22 | < 0.01 | 14.60 | < 0.01 | 22.75 | < 0.01 | 492.44 | < 0.01 | |||
| 源株密度Ortet density | 24.49 | < 0.01 | 134.87 | < 0.01 | 8.46 | < 0.01 | 36.50 | < 0.01 | |||
| 生境×源株密度 Habitat × Ortet density |
29.42 | < 0.01 | 11.45 | < 0.01 | 8.69 | < 0.01 | 5.51 | < 0.01 | |||
生长季末(10月), 异质生境及不同源株密度影响下互花米草的株高状况如图 3所示。生境异质性对互花米草株高有极显著影响(P < 0.01, 表 1)。北断面植被区(芦苇群落, N1)互花米草平均株高为(148.86±9.99) cm, 显著高于南断面植被区(海三棱藨草群落, S1)互花米草平均株高(74.7±8.87) cm(P < 0.05, 图 3)。此外, 在植被区, 随着源株密度增加, 互花米草的株高无显著差异(P > 0.05, 图 3)。光滩区只有在S2生境, 且当互花米草源株密度为20株/m2时, 才能一直保持存活, 生长季末的株高为(43.33±5.35) cm, 显著低于植被区(N1和S1)相同源株密度处理(P < 0.05)。

|
| 图 3 异质生境及不同源株密度影响下互花米草的株高差异 Fig. 3 Difference of the hight of S. alterniflora under different ortet density in different habitats at the end of growing season 不同小写字母的柱状图表示相互之间存在显著差异, P < 0.05 |
生境异质性及源株密度均对互花米草种群增长率产生极显著影响(P < 0.01, 表 1)。至生长季末, 植被区(N1和S1)的互花米草种群整体呈现增长趋势(图 4), N1生境互花米草平均种群增长率((280.89±416.05)%)高于S1生境互花米草平均种群增长率((173.89±262.07)%); 而光滩(N2和S2)生境中, 互花米草种群整体则处于萎缩状态(图 4), N2生境由于移栽互花米草全部被潮水冲走, 种群增长率为-100%, S2生境仅高源株密度(20株/m2)互花米草成功定居并扩散, 其种群增长率为(150±200)%, 显著高于植被区(N1和S1)同源株密度处理(P < 0.05)。此外, 在植被区, 互花米草种群增长率与源株密度呈负相关(图 4)。其中, N1生境1株/m2源株密度处理下互花米草的种群增长率为(900±256.78)%, 显著高于其余源株密度处理(3株/m2:(311.11±254.59)%; 5株/m2:(93.33±105.26)%; 10株/m2:(66.67±47.26)%; 20株/m2:(33.33±51.32)%)(P < 0.05);S2生境互花米草种群增长率变化趋势与N1相似, 即随着源株密度增加, 种群增长率显著降低(P < 0.05, 图 4)。

|
| 图 4 异质生境及不同源株密度影响下互花米草种群增长率的差异 Fig. 4 Difference of population growth rate of S. alterniflora under different ortet density in different habitats at the end of growing season 不同小写字母的柱状图表示相互之间存在显著差异, P < 0.05 |
从表 2可以看出, 4个生境的环境因子存在明显差异。粒度分析结果表明, 北断面(N1和N2)中值粒径显著小于南断面(S1和S2, P < 0.05, 表 2)。冲淤动态监测结果显示, 南断面(S1和S2)的滩面整体以淤积状态为主, 至11月, 植被区(S1)和光滩区(S2)滩面高程分别累计抬升了(28.06±1.62) cm和(19.51±1.02) cm, 植被区的淤积速率显著高于光滩(P < 0.05);而北断面(N1和N2)处于有冲有淤、整体微冲的状态, 植被区(N1)和光滩区(N2)滩面累计侵蚀量分别为(0.55±3.28) cm和(5.99±1.72) cm, 光滩(N1)的侵蚀速率显著高于植被区(N2, P < 0.05)。此外, 南北断面, 植被区土壤电导率、淹水时间和淹水深度均显著小于光滩(P < 0.05)。
| 生境类型 Habitat types |
中值粒径 Median particle size/μm |
电导率 Conductivity/(mS/m) |
累计冲淤 Accumulative scouring and silting/cm |
淹水时间 Flooding period/min |
淹水深度 Flooding depth/m |
| N1 | 13.64±1.41c | 816.92±80.36b | -0.55±3.28c | 324.06±2.82c | 1.71±0.01d |
| N2 | 15.59±2.67c | 971.4±114.05a | -5.99±1.72d | 395.99±2.81a | 2.31±0.02b |
| S1 | 38.72±3.63b | 823.5±76.94b | 28.06±1.62a | 365.33±2.31b | 2.00±0.01c |
| S2 | 133.18±11.98a | 886.53±80.36a | 19.51±1.02b | 398.97±5.61a | 2.37±0.05a |
| 表中同一列数据标注不同小写字母表示相互之间存在显著差异(P < 0.05), N1:北断面植被区Vegetation region of the north section; N2:北断面光滩区Mudflat of the north section; S1:南断面植被区Vegetation region of the south section; S2:南断面光滩区Mudflat of the south section | |||||
环境因子主成分分析结果见表 3, 前2个特征值对应的累计方差贡献率已达总方差的86.53%(>85%), 可代表所选指标的绝大部分信息。其中, 第一主成分占总方差51.80%, 该成分中淹水时间和淹水深度载荷最高, 主要反映了高程对植物的影响。第二主成分与冲淤相关性最高, 主要表现沉积地貌对植物的影响。
| 主成分 Principal component |
重要值 Important value |
解释变异百分比 Percentage of variance explained |
累计解释变异百分比 Accumulative percentage of variance explained |
主成分矩阵Principal component matrix | ||||
| 中值粒径 Median particle size |
电导率 Conductivity |
冲淤 Scour and silt |
淹水时间 Flooding time |
淹水深度 Flooding depth |
||||
| 1 | 2.590 | 51.797 | 51.797 | 0.530 | 0.667 | 0.029 | 0.959 | 0.971 |
| 2 | 1.737 | 34.737 | 86.534 | 0.738 | -0.590 | 0.894 | 0.157 | 0.143 |
对克隆植物的研究发现, 当生长空间较充沛时, 高密度源株具有较强的存活能力及分蘖能力; 而当生长空间有限时, 源株较多会导致竞争, 从而降低植株存活率及分蘖能力[40-41]。本研究中, 植被区由于已有本地植物生长, 移栽互花米草的生存空间相对有限, 密度效应明显。当源株密度越高时, 受到有限环境资源的限制和种间及种内竞争的影响也越大, 进而互花米草存活率和种群增长率也越低。这主要是由于植被区环境干扰小、生境稳定, 在周围植物的遮护下, 以低源株密度定植的互花米草可以更好地利用有限资源, 生存并产生新分蘖, 相反, 高密度则会由于源株拥挤而导致竞争, 使无性分蘖及存活率下降[14, 16]。在光滩区, 由于不受生长空间的限制, 高源株密度处理下互花米草存活率显著高于低源株密度处理, 这与之前的互花米草移栽实验结果[7]也基本一致。这可能是因为光滩区域虽然有充沛的生长空间, 但同时缺乏周围植物的庇护, 高密度互花米草种群的“抱团”效应会促进邻近源株的存活, 从而促进种群对生境的占领。赵春燕和沈有信[42]在模拟源株密度对入侵植物紫茎泽兰(Eupatorium coelestinum)生长和竞争影响的研究中也同样发现, 源株密度控制着紫茎泽兰叶片和根蘖株等构件和无性系植株的形成率, 源株密度越大, 紫茎泽兰个体株高、叶片数、根蘖株数和生物量越少, 而种群死亡率越高。于国磊[43]开展的入侵克隆植物空心莲子草(Alternanthera philoxeroides)种内关系的室外实验也得出了相似的结论, 即空心莲子草的生长和表型特征也受密度效应的调控, 其叶面积、节数、总生物量、根和叶生物量与密度呈显著负相关。可见, 源株密度对克隆植物, 尤其是对以克隆繁殖为主要繁殖途径的外来物种的入侵力具有重要的影响作用。
此外, 研究也表明, 盐沼植物成功定居或扩散存在阈值效应, 包括生物阈值(生物量、根长等)[44-45]和环境阈值(高程、潮流、波浪等)[46-48]。在本研究中, 互花米草的存活时间总体与源株密度呈正相关。在植被区, 互花米草的存活和定居不受源株密度限制, 1株/m2即可成功入侵。在光滩区, 尽管该区域高程已经能够满足互花米草存活的最低高程要求, 但由于受潮流等环境因子的干扰相对较大, 互花米草存活的生物阈值也较高。仅当源株密度为20株/m2时, 互花米草可以成功在粉砂质光滩(S2)定居, 且可一直保留至当年生长季末; 而在淤泥质光滩(N2), 当互花米草源株密度为20株/m2时, 在移栽初期和中期(4—8月)互花米草可以存活, 但随着8月下旬的台风等环境条件的扰动, 最终未能保留至生长季末, 可能需要更大的源株密度, 才能实现永久定居。这也表明生境异质性也会影响互花米草入侵的生物阈值。
3.2 异质环境对互花米草入侵力的影响分析外来入侵种的形态和生理特征往往会对异质生境表现出较强的可塑性, 其在不同资源和胁迫背景下的构型趋异是入侵能力的外在特征体现[49-50]。表型可塑性被认为是预测外来种入侵力的一个特征[51]。许凯扬等[52]报道不同生境(水生、湿生、中生和旱生)中的喜旱莲子草表现出显著的表型趋异, 随土壤含水量减少, 喜旱莲子草的根冠比显著增加, 分支强度显著降低。本研究中, 高程越低、淹水胁迫越大、潮滩处于冲刷状态, 高源株密度处理下互花米草株高越低, 而种群增长率却越高, 表现出明显的表型趋异及对异质性生境的适应。这主要因为高程(或与其相关的环境因子)影响了互花米草的资源分配格局, 强胁迫条件下, 互花米草会优先扩张种群, 缓解外界干扰压力, 提高环境适应度, 进而成功拓殖异质生境。沙晨燕等[53]在研究典型滩涂环境因子对植物群落空间分布的影响时得出了相似的结论, 互花米草的盖度与密度随土壤紧实度、养分有效性增大而增加; 随着高程的升高, 互花米草的生物量、株高反而下降。
生境条件的异质性大大增加了盐沼植物吸收和利用资源的难度, 也严重影响其存活、繁殖和扩散[7-8]。本研究中, 冲淤动态、淹水时间和淹水深度是决定互花米草生存状况的主要影响因子。粉砂质光滩(S2)整体以淤积状态为主, 这在一定程度上促进了互花米草的定居, 增强了已定居植物对环境干扰的抵抗力; 而淤泥质光滩(N2)滩面处于侵蚀状态, 沉积物的大量冲刷不利于定居植物的保留。相对于沉积物淤积, 盐沼植物对侵蚀过程更为敏感, 不同量级的沉积物扰动过程会直接或间接地影响植物存活[25]。此外, 在不同淹水时间和淹水深度影响下, 互花米草的入侵力也表现出明显差异, 生长季末90%以上的存活互花米草均分布在淹水时间和淹水深度显著较低的植被区(N1和S1), 其株高也显著高于受淹水胁迫较大的光滩区存活植株。一方面可能是因为光滩区较长周期的潮水淹没会导致植物缺氧、降低光合作用, 直接抑制互花米草的生长; 另一方面土壤氧化还原电位、海水矿物质、及光强等生境资源会因淹水时间和深度增大而发生明显变化, 这可能也会间接影响互花米草的生长和分布[54]。
3.3 河口湿地互花米草入侵力及其管控入侵植物的入侵力取决于活力繁殖体的可传播性及定植机会窗口(一段可用来定植与繁殖的无干扰时间)的时空可利用性[44, 55]。已有研究表明, 互花米草有性繁殖体和无性繁殖体均可在潮流作用下进行远距离传播, 进而开拓新生境[56-57]。盐沼植物种子随潮流传播命运及实生苗定植机会窗口的研究表明:受水动力及沉积动力的影响, 盐沼植物的种子很难通过长距离传播实现成功定居[58], 且即使成功保留在种子库中的种子也并不意味着定植成功[59]。对实生苗而言, 由于受到生态瓶颈(繁殖力、传播能力等)和物理压力(水动力、盐度等)阈值效应的影响及机会窗口的限制, 实生苗定植新生境的成功率普遍较低[60-61]。但在河口强潮动力作用下, 互花米草根茎苗或根状茎易被潮水冲刷并脱离母体[62], 虽然数量较少, 但定居成功率、生长、繁殖等指标均优于有性繁殖方式[18]。
本研究中, 1株互花米草根茎苗即可在植被区实现稳定存活、种群扩张和成功入侵, 当源株密度足够大, 亦可成功入侵光滩区。若不加以重视, 成功定居的零星互花米草能以无性繁殖的方式充分占据空间优势, 实现肆意蔓延的强劲态势, 必然会对滩涂湿地的生态恢复与生物多样性的保护带来极大的负面影响。此外, 近年来, 我国海岸带的圈围工程, 也显著改变了潮滩的沉积环境, 导致泥沙在近岸区域持续淤积, 并向海延伸, 形成了大面积的新生滩涂[63-64], 也为随潮流传播的互花米草繁殖体提供了合适的入侵生境。因此, 基于互花米草入侵机制的研究, 在入侵早期, 通过适当人为干预, 阻截其进一步爆发, 可大大降低互花米草的入侵风险和后期控制的经济成本, 对河口湿地的保育和可持续管理具有重要意义。
| [1] | Tong C, Wang W Q, Huang J F, Gauci V, Zhang L H, Zeng C S. Invasive alien plants increase CH4 emissions from a subtropical tidal estuarine wetland. Biogeochemistry, 2012, 111(1-3): 677–693. DOI:10.1007/s10533-012-9712-5 |
| [2] | Barbier E B, Koch E W, Silliman B R, Hacker S D, Wolanski E, Primavera J, Granek E F, Polasky S, Aswani S, Cramer L A, Stoms D M, Kennedy C J, Bael D, Kappel C V, Perillo G M E, Reed D J. Coastal ecosystem-based management with nonlinear ecological functions and values. Science, 2008, 319(5861): 321–323. DOI:10.1126/science.1150349 |
| [3] | Temmerman S, Meire P, Bouma T J, Herman P M, Ysebaert T, De Vriend H J. Ecosystem-based coastal defence in the face of global change. Nature, 2013, 504(7478): 79–83. DOI:10.1038/nature12859 |
| [4] | Zhou R, Li Y Z, Wu J J, Gao M, Wu X Q, Bi X L. Need to link river management with estuarine wetland conservation:A case study in the Yellow River Delta, China. Ocean & Coastal Management, 2017, 146: 43–49. |
| [5] | 任璘婧, 李秀珍, 杨世伦, 闫中正, 黄星. 崇明东滩盐沼植被变化对滩涂湿地促淤消浪功能的影响. 生态学报, 2014, 34(12): 3350–3358. |
| [6] | 王丽荣, 赵焕庭. 中国河口湿地的一般特点. 海洋通报, 2000, 19(5): 47–54. DOI:10.3969/j.issn.1001-6392.2000.05.007 |
| [7] | Schwarz C, Ysebaert T, Zhu Z C, Zhang L Q, Bouma T J, Herman P M J. Abiotic factors governing the establishment and expansion of two salt marsh plants in the Yangtze Estuary, China. Wetlands, 2011, 31(6): 1011–1021. DOI:10.1007/s13157-011-0212-5 |
| [8] | 曹浩冰, 葛振鸣, 祝振昌, 张利权. 崇明东滩盐沼植被扩散格局及其形成机制. 生态学报, 2014, 34(14): 3944–3952. |
| [9] | Japhet W, Zhou D W, Zhang H X, Zhang H X, Yu T. Evidence of phenotypic plasticity in the response of Fagopyrum esculentum, to population density and sowing date. Journal of Plant Biology, 2009, 52(4): 303–311. DOI:10.1007/s12374-009-9037-7 |
| [10] | 黎磊, 耿宇鹏, 兰志春, 陈家宽, 宋志平. 异质生境中水生植物表型可塑性的研究进展. 生物多样性, 2016, 24(2): 216–227. |
| [11] | Cai Y, Guan B H, An S Q, Shen R L, Jiang J H, Dong L. Phenotypic plasticity of the clonal plant trapa bicornis in response to sediment phosphorus concentration and plant density. Journal of Plant Ecology, 2007, 31(4): 599–606. DOI:10.17521/cjpe.2007.0076 |
| [12] | 田耀华, 冯玉龙, 刘潮. 氮肥和种植密度对紫茎泽兰生长和竞争的影响. 生态学杂志, 2009, 28(4): 577–588. |
| [13] | Toillon J, Fichot R, Dallé E, Berthelot A, Brignolas F, Marron N. Planting density affects growth and water-use efficiency depending on site in populus deltoides×p. nigra. Forest Ecology and Management, 2013, 304: 345–354. DOI:10.1016/j.foreco.2013.05.017 |
| [14] | 王永健, 钟章成. 模拟源株密度对蝴蝶花生长和克隆繁殖的影响. 植物生态学报, 2010, 34(3): 340–347. DOI:10.3773/j.issn.1005-264x.2010.03.011 |
| [15] | Worm B, Reusch T B H. Do nutrient availability and plant density limit, seagrass colonization in the Baltic Sea?. Marine Ecology Progress, 2000, 200(1): 159–166. |
| [16] | 钟青龙, 戴文龙, 项世亮, 王天厚, 汪承焕. 密度对三种莎草科植物克隆生长的影响. 生态科学, 2016, 35(1): 1–9. DOI:10.3969/j.issn.1671-6361.2016.01.001 |
| [17] | 祝振昌, 张利权, 肖德荣. 上海崇明东滩互花米草种子产量及其萌发对温度的响应. 生态学报, 2011, 31(6): 1574–1581. |
| [18] | 徐伟伟, 王国祥, 刘金娥, 陈正勇, 杭子清, 王会. 苏北海滨湿地互花米草种群繁殖方式. 生态学报, 2014, 34(14): 3839–3847. |
| [19] | Liu W W, Maungdouglass K., Strong D R., Pennings S C, Zhang Y H. Geographical variation in vegetative growth and sexual reproduction of the invasive Spartina alterniflora in China. Journal of Ecology, 2016, 104(1): 173–181. DOI:10.1111/1365-2745.12487 |
| [20] | Li Z J, Wang W Q, Zhang Y H. Recruitment and herbivory affect spread of invasive Spartina alterniflora in China. Ecology, 2016, 95(7): 1972–1980. |
| [21] | 邓自发, 安树青, 智颖飙, 周长芳, 陈琳, 赵聪蛟, 方淑波, 李红丽. 外来种互花米草入侵模式与爆发机制. 生态学报, 2006, 26(8): 2678–2686. DOI:10.3321/j.issn:1000-0933.2006.08.034 |
| [22] | Castillo J M, Grewell B J, Pickart A, Bortolus A, Peña C, Figueroa E, Sytsma M. Phenotypic plasticity of invasive Spartina densiflora (Poaceae) along a broad latitudinal gradient on the Pacific Coast of North America. American Journal of Botany, 2014, 101(3): 448–458. DOI:10.3732/ajb.1400014 |
| [23] | Xue L, Li X Z, Yan Z Z, Zhang Q, Ding W H, Huang X, Tian B, Ge Z M, Yin Q X. Native and non-native halophytes resiliency against sea-level rise and saltwater intrusion. Hydrobiologia, 2018, 806: 47–65. DOI:10.1007/s10750-017-3333-x |
| [24] | Ge Z M, Cao H B, Cui L F, Zhao B, Zhang L Q. Future vegetation patterns and primary production in the coastal wetlands of East China under sea level rise, sediment reduction, and saltwater intrusion. Journal of Geophysical Research:Biogeosciences, 2015, 120: 1923–1940. DOI:10.1002/2015JG003014 |
| [25] | Cao H B, Zhu Z C, Balke T, Zhang L Q, Bouma T J. Effects of sediment disturbance regimes on Spartina seedling establishment:implications for salt marsh creation and restoration. Limnology & Oceanography, 2018, 63: 647–659. |
| [26] | 石福臣, 鲍芳. 盐和温度胁迫对外来种互花米草(Spartina alterniflora)生理生态特性的影响. 生态学报, 2007, 27(7): 2733–2741. DOI:10.3321/j.issn:1000-0933.2007.07.009 |
| [27] | 何军, 赵聪蛟, 清华, 甘琳, 安树青. 土壤水分条件对克隆植物互花米草表型可塑性的影响. 生态学报, 2009, 29(7): 3518–3524. DOI:10.3321/j.issn:1000-0933.2009.07.009 |
| [28] | Ma Z J, Melville D S, Liu J G, Chen Y, Yang H Y, Ren W W, Zhang Z W, Piersma T, Li B. Rethinking China's new great wall:massive seawall construction in coastal wetlands threatens biodiversity. Science, 2014, 346(6212): 912–914. DOI:10.1126/science.1257258 |
| [29] | 崔利芳, 王宁, 葛振鸣, 张利权. 海平面上升影响下长江口滨海湿地脆弱性评价. 应用生态学报, 2014, 25(2): 553–561. |
| [30] | Yuan L, Ge Z M, Fan X Z, Zhang L Q. Ecosystem-based coastal zone management:A comprehensive assessment of coastal ecosystems in the Yangtze Estuary coastal zone. Ocean & Coastal Management, 2014, 95(95): 63–71. |
| [31] | 黄华梅.上海滩涂盐沼植被的分布格局和时空动态研究[D].上海: 华东师范大学, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D317058 |
| [32] | Xiao D R, Zhang L Q, Zhu Z C. The range expansion patterns of Spartina alterniflora on salt marshes in the Yangtze Estuary, China. Estuarine Coastal and Shelf Science, 2010, 88(1): 99–104. DOI:10.1016/j.ecss.2010.03.015 |
| [33] | Yuan L, Zhang L Q, Xiao D R, Huang H M. The application of cutting plus waterlogging to control Spartina alterniflora on saltmarshes in the Yangtze Estuary, China. Estuarine Coastal and Shelf Science, 2011, 92(1): 103–110. DOI:10.1016/j.ecss.2010.12.019 |
| [34] | Niu J Y, Zou Y A, Yuan X, Zhang B, Wang T H. Waterbird distribution patterns and environmentally impacted factors in reclaimed coastal wetlands of the eastern end of Nanhui county, Shanghai, China. Acta Zoologica Academiae Scientiarum Hungaricae, 2013, 59(2): 171–185. |
| [35] | 陶燕东, 于克锋, 何培民, 孙彬, 历成伟, 赵萌, 方淑波. 围垦后南汇东滩海三棱藨草的空间分布及其影响因子研究. 长江流域资源与环境, 2017, 26(7): 1032–1041. DOI:10.11870/cjlyzyyhj201707009 |
| [36] | 冯凌旋, 李占海, 李九发, 周云轩, 陈炜, 徐敏. 基于机制分解法长江口南汇潮滩悬移质泥沙通量研究. 长江流域资源与环境, 2011, 20(8): 944–950. |
| [37] | 黄华梅, 张利权, 袁琳. 崇明东滩自然保护区盐沼植被的时空动态. 生态学报, 2007, 27(10): 4166–4172. DOI:10.3321/j.issn:1000-0933.2007.10.025 |
| [38] | 赵敏. 潮滩冲淤监测方法与实时监测系统设计. 地球科学与环境学报, 2009, 31(4): 437–441. DOI:10.3969/j.issn.1672-6561.2009.04.015 |
| [39] | 李蕙, 袁琳, 张利权, 李伟, 李诗华, 赵志远. 长江口滨海湿地潮间带生态系统的多稳态特征. 应用生态学报, 2017, 28(1): 327–336. |
| [40] | Oborny B, Ádám Kun, Czárán T, Bokros S. The effect of clonal integration on plant competition for mosaic habitat space. Ecology, 2000, 81(12): 3291–3304. DOI:10.1890/0012-9658(2000)081[3291:TEOCIO]2.0.CO;2 |
| [41] | Kleunen M V, Fischer M, Schmid B. Effects of intraspecific competition on size variation and reproductive allocation in a clonal plant. Oikos, 2001, 94(3): 515–524. DOI:10.1034/j.1600-0706.2001.940313.x |
| [42] | 赵春燕, 沈有信. 紫茎泽兰种内竞争试验. 生态与农村环境学报, 2008, 24(2): 27–31. DOI:10.3969/j.issn.1673-4831.2008.02.006 |
| [43] | 于国磊. 水淹对克隆植物空心莲子草种内关系的影响. 植物生态学报, 2011, 35(9): 973–980. |
| [44] | Balke T, Bouma T J, Horstman E M, Webb E L, Erftemeijer P L A, Herman P M J. Windows of opportunity:thresholds to mangrove seedling establishment on tidal flats. Marine Ecology Progress, 2011, 440(1): 1–9. |
| [45] | Angelini C, Silliman B R. Patch size-dependent community recovery after massive disturbance. Ecology, 2012, 93(1): 101–110. DOI:10.1890/11-0557.1 |
| [46] | Wang C, Temmerman S. Does bio-geomorphic feedback lead to abrupt shifts between alternative landscape states? An empirical study on intertidal flats and marshes. Journal of Geophysical Research Earth Surface, 2013, 118(1): 229–240. DOI:10.1029/2012JF002474 |
| [47] | Hu Z, van Belzen J, van der Wal D, Balke T, Wang Z B, Stive M, Bouma T J. Windows of opportunity for salt marsh vegetation establishment on bare tidal flats:The importance of temporal and spatial variability in hydrodynamic forcing. Journal of Geophysical Research:Biogeosciences, 2015, 120(7): 1450–1469. DOI:10.1002/2014JG002870 |
| [48] | Silinski A, Heuner M, Schoelynck J, Puijalon S, Schröder U, Fuchs E, Troch P, Bouma T J, Meire P, Temmerman S. Effects of wind waves versus ship waves on tidal marsh plants:a flume study on different life stages of Scirpus maritimus. Plos One, 2015, 10(3): e0118687. DOI:10.1371/journal.pone.0118687 |
| [49] | Meekins J F, Mccarthy B C. Effect of environmental variation on the invasive success of a nonindigenous forest herb. Ecological Applications, 2001, 11(5): 1336–1348. DOI:10.1890/1051-0761(2001)011[1336:EOEVOT]2.0.CO;2 |
| [50] | Claridge K, Franklin S B. Compensation and plasticity in an invasive plant species. Biological Invasions, 2002, 4(4): 339–347. DOI:10.1023/A:1023671006342 |
| [51] | Rejmánek M, Richardson D M. What attributes make some plant species more invasive?. Ecology, 1996, 77(6): 1655–1661. DOI:10.2307/2265768 |
| [52] | 许凯扬, 叶万辉, 李静, 黄忠良, 李国民. 入侵种喜旱莲子草对土壤水分的表型可塑性反应. 华中师范大学学报(自然科学版), 2005, 39(1): 100–103. DOI:10.3321/j.issn:1000-1190.2005.01.021 |
| [53] | 沙晨燕, 李玲, 唐浩, 王卿, 王敏, 熊丽君, 吴健. 典型滩涂环境因子对植物群落空间分布的影响. 人民长江, 2016, 47(22): 10–15. |
| [54] | Hofmann H, Lorke A, Peeters F. Temporal scales of water-level fluctuations in lakes and their ecological implications. Hydrobiologia, 2008, 613(1): 85–96. DOI:10.1007/s10750-008-9474-1 |
| [55] | Gabler C A, Siemann E. Environmental variability and ontogenetic niche shifts in exotic plants may govern reinvasion pressure in restorations of invaded ecosystems. Restoration Ecology, 2012, 20(5): 545–550. DOI:10.1111/rec.2012.20.issue-5 |
| [56] | Feist B E, Simenstad C A. Expansion rates and recruitment frequency of exotic smooth cordgrass, Spartina alterniflora (Loisel), colonizing unvegetated littoral flats in Willapa Bay, Washington. Estuaries, 2000, 23(2): 267–274. DOI:10.2307/1352832 |
| [57] | Taylor C M, Hastings A. Finding optimal control strategies for invasive species:a density-structured model for Spartina alterniflora. Journal of Applied Ecology, 2004, 41(6): 1049–1057. DOI:10.1111/j.0021-8901.2004.00979.x |
| [58] | Zhu Z C, Bouma T J, Ysebaert T, Zhang L Q, Herman P M J. Seed arrival and persistence at the tidal mudflat:Identifying key processes for pioneer seedling establishment in salt marshes. Marine Ecology Progress, 2014, 513(513): 97–109. |
| [59] | Nathan R, Mullerlandau H C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in ecology & evolution, 2000, 15(7): 278–285. |
| [60] | Bouma T J, Friedrichs M, Klaassen P, Van Wesenbeeck B K, Brun F G, Temmerman S, Van Wesenbeeck M M, Graf G, Herman P M J. Effects of shoot stiffness, shoot size and current velocity on scouring sediment from around seedlings and propagules. Marine Ecology Progress, 2009, 388(12): 293–297. |
| [61] | Friess D A, Krauss K W, Horstman E M, Balke T, Bouma T J, Galli D, Webb E L. Are all intertidal wetlands naturally created equal? Bottlenecks, thresholds and knowledge gaps to mangrove and saltmarsh ecosystems. Biological Reviews of the Cambridge Philosophical Society, 2012, 87(2): 346–366. DOI:10.1111/brv.2012.87.issue-2 |
| [62] | Daehler C C, Strong D R. Variable reproductive output among clones of Spartina alterniflora (Poaceae) invading San Francisco Bay, California:the influence of herbivory, pollination, and establishment site. American Journal of Botany, 1994, 81(3): 307–313. DOI:10.1002/j.1537-2197.1994.tb15448.x |
| [63] | 李九发, 戴志军, 刘新成, 赵建春, 冯凌旋. 长江河口南汇嘴潮滩圈围工程前后水沙运动和冲淤演变研究. 泥沙研究, 2010(3): 31–37. |
| [64] | 路兵, 蒋雪中. 滩涂围垦对崇明东滩演化影响的遥感研究. 遥感学报, 2013, 17(2): 335–349. |