 2018, Vol. 38
2018, Vol. 38文章信息
- 求锦津, 王咏雪, 李铁军, 孙鹏, 张苗苗, 聂振林, 俞松立, 田阔, 水柏年.
- QIU Jinjin, WANG Yongxue, LI Tiejun, SUN Peng, ZHANG Miaomiao, NIE Zhenlin, YU Songli, TIAN Kuo, SHUI Bonian.
- 舟山长白海域主要游泳动物生态位及其分化研究
- Study on the niche and differentiation of major nekton species in the Zhoushan Changbai sea area
- 生态学报. 2018, 38(18): 6759-6767
- Acta Ecologica Sinica. 2018, 38(18): 6759-6767
- http://dx.doi.org/10.5846/stxb201803300680
-
文章历史
- 收稿日期: 2018-03-30
- 修订日期: 2018-07-04





2. 台州市海洋与渔业局, 台州 318000;
3. 浙江省海洋水产研究所, 舟山 316000
2. Taizhou City Ocean and Fisheries Bureau, Taizhou 318000, China;
3. Marine Fisheries Research Institute of Zhejiang Province, Zhoushan 316000, China
生态位是指在自然生态系统中, 一个种群在时间、空间上的位置及其与相关种群之间的功能关系, 在研究生物群落结构、种间关系和生物多样性等方面有重要的作用[1]。生态位宽度和生态位重叠可以解释物种在丰度、分布等方面的差异, 可以用来研究物种对资源空间的利用程度和物种间的竞争能力, 还可以比较不同物种对环境的适应能力[2-3]。有关生态位的研究多集中于陆生生物群落[4-7], 近年来, 生态位的研究对象已逐步扩展到海洋生物群落, 如汤雁滨等[8]、葛宝明等[9]及彭松耀等[10]均采用生态位理论研究了大型底栖动物的生态位宽度和重叠值; 李显森等[11]、胡成业等[12]及董静瑞等[13]则对主要鱼类的生态位进行了探究; 另外, 李超男等[14]、胡成业等[15]运用生态位测度、种间联结性测度等方法研究了主要游泳动物生态位特征和种间关系。
舟山长白海域位于舟山本岛西北部的近岸, 既毗邻杭州湾口又位于岱衢洋海域, 为海洋生态敏感区, 关于该海域主要游泳动物群落生态位研究及其报道罕见, 而主要游泳动物群落的动态过程在一定程度上代表着整个游泳动物群落的变化特征。本文拟运用相对重要性指数、生态位宽度、生态位重叠以及冗余分析(RDA)等方法, 研究舟山长白海域主要游泳动物的生态位, 种间生态关系以及多个环境因子影响下的生态位分化, 以期为舟山长白及邻近海域游泳动物资源生态保护、可持续利用以及科学管理的研究提供理论依据。
1 材料与方法 1.1 数据来源依据《海洋调查规范》[16], 于2016年10月(秋季)和2017年4月(春季)在舟山长白海域(121°49′54.55″—122°18′33.98″E, 30°01′44.46″—30°18′58.27″N)开展了渔业资源调查, 2季分别设置12个站位(图 1)。渔业资源调查所获渔获物经低温保存运回实验室, 进行种类鉴定、生物学测定等, 物种鉴定依据相关文献[17-20]。各站位还同步调查了温度、盐度、悬浮物、溶解氧、化学需氧量和pH等环境因子。

|
| 图 1 舟山长白海域渔业资源调查站位 Fig. 1 Survey stations of fisheries resource in Changbai |
采用Jaccard′s种类相似性指数[21]计算春、秋2季游泳动物的相似程度, 计算公式如下:
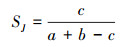
|
(1) |
式中, a、b分别为秋季、春季的游泳动物种类总数; c为2次调查共有的游泳动物种数。当0≤SJ<0.25时, 为极不相似; 当0.25≤SJ<0.5时, 为中等不相似; 当0.5≤SJ<0.75时, 为中等相似; 当0.75≤SJ≤1.00时, 为极相似。
1.2.2 优势种优势种判断采用Pinkas的相对重要性指数IRI[22], 计算公式如下:
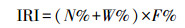
|
(2) |
式中, N为某物种尾数占总尾数的百分比, W为该物种重量占总重量的百分比, F为出现站位百分比。优势种的判定采用王雪辉等[23]的划分标准, 将IRI>1000的种类定为优势种, 1000—100为重要种, 100—10为常见种, 10—1为一般种, <1为少有种。参照相关文献[13-14], 将IRI>100的种类定为主要游泳动物。
1.2.3 生态位测度生态位宽度用于反映物种的分布范围、分布格局以及对环境的适应能力或对资源的利用程度[12, 24], 本文采用Shannon指数[25]评价, 计算公式如下:
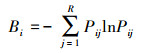
|
(3) |
生态位重叠主要反映了物种之间资源利用的相似程度和竞争关系[7], 本文采用Pianka的重叠指数[26]评价, 计算公式如下:
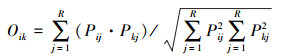
|
(4) |
式中, Pij和Pkj分别为种i和种k在j个站位中的个体数占站位总个体数的比例; R为总站位数; Bi为生态位宽度, 取值范围为[0, R], Bi值越大, 生态位宽度就越大; Oik为生态位重叠值, 取值范围为[0, 1]。根据Wnthne等的划分标准, 将生态位重叠值划分为3个等级, 即Oik>0.6, 重叠度较高; 0.3—0.6, 重叠度较低; Oik<0.3, 重叠度低[27]。生态位宽度值和生态位重叠的计算在Excel软件里进行。另外, 运用SPSS 23软件对相对重要性指数值和生态位宽度进行Pearson相关性分析。
1.2.4 生态位分化运用Canoco 4.5软件对主要游泳动物与环境因子之间的关系进行分析及排序图绘制, 以揭示主要游泳动物生态位分化的状态。先对物种数据进行除趋势对应分析(DCA), 根据每个轴的梯度长度(LGA)选择适宜的排序方法。当LGA<3时, 选择冗余分析(RDA); 当LGA>4时, 则可运用典范对应分析(CCA); 当3<LGA<4时, 2种分析均可[28]。DCA分析结果显示, LGA值均<3, 故本文采用线性模型的RDA分析。
2 结果 2.1 物种组成春、秋2季共捕获游泳动物37种, 隶属于10目23科33属。秋季28种, 隶属于9目20科24属, 其中鱼类17种, 虾类6种, 蟹类5种; 春季30种, 隶属于8目20科28属, 其中鱼类18种, 虾类8种, 蟹类4种。2个季节的共同种有21种, 其中鱼类共同种11种, 虾类6种, 蟹类4种。经计算, 游泳动物群落的Jaccard’s种类相似性指数值为0.57, 表明春、秋2季游泳动物种类组成为中等相似。
2.2 优势种与重要种由表 1的IRI指数值得出, 秋季优势种为龙头鱼与安氏白虾, 重要种为棘头梅童鱼、鮸、口虾蛄、葛氏长臂虾、三疣梭子蟹与日本蟳6种; 春季优势种有孔鰕虎鱼、脊尾白虾、细鳌虾与三疣梭子蟹, 重要种为龙头鱼、刀鲚、棘头梅童鱼、鮸、蓝点马鲛、葛氏长臂虾与日本蟳7种。秋季优势种和重要种的尾数和重量占比分别为97.19%与85.13%, 春季分别为85.94%与88.76%。本文特将春、秋2季的优势种与重要种定为主要游泳动物, 作为生态位分析和冗余分析的研究对象。
| 物种 Species |
相对重要性指数 Relative importance index |
|
| 秋季Autumn | 春季Spring | |
| 龙头鱼 Harpodon nehereus |
7760.86 | 226.84 |
| 刀鲚 Coilia nasus |
466.88 | |
| 棘头梅童鱼 Collichthys lucidus |
954.46 | 287.34 |
| 鮸 Miichthys niiuy |
106.45 | 411.51 |
| 蓝点马鲛 Scomberomorus niphonius |
282.75 | |
| 孔鰕虎鱼 Trypauchen vagina |
1425.02 | |
| 口虾蛄 Oratosquilla oratoria |
169.66 | |
| 安氏白虾 Exopalaemon annandalei |
4429.09 | |
| 脊尾白虾 Exopalaemon carinicauda |
1046.88 | |
| 葛氏长臂虾 Palaemon gravieri |
698.67 | 243.12 |
| 细鳌虾 Leptochela gracilis |
1783.96 | |
| 三疣梭子蟹 Portunus trituberculatus |
545.07 | 5133.54 |
| 日本蟳 Charybdis japonica |
750.04 | 108.23 |
春、秋2季主要游泳动物生态位宽度值差异均较大(表 2)。秋季, 主要游泳动物生态位宽度值的变化范围为0.05—3.90, 其中生态位宽度最大的是龙头鱼(3.90), 其次是安氏白虾(3.69), 鮸的生态位宽度最小(0.05);春季, 主要游泳动物的生态位宽度值变化范围为0.13—3.71, 三疣梭子蟹的生态位宽度值最大(3.71), 其次为细鳌虾(3.43), 生态位宽度值最小的是蓝点马鲛(0.13)。
| 秋季Autumn | 春季Spring | |||||
| 种号Number | 物种 Species |
生态位宽度 Niche breadth |
种号Number | 物种 Species |
生态位宽度 Niche breadth |
|
| A1 | 龙头鱼 | 3.90 | S1 | 龙头鱼 | 0.88 | |
| A2 | 棘头梅童鱼 | 1.13 | S2 | 刀鲚 | 1.68 | |
| A3 | 鮸鱼 | 0.05 | S3 | 棘头梅童鱼 | 0.98 | |
| A4 | 口虾蛄 | 0.67 | S4 | 鮸鱼 | 0.18 | |
| A5 | 安氏白虾 | 3.69 | S5 | 蓝点马鲛 | 0.13 | |
| A6 | 葛氏长臂虾 | 1.61 | S6 | 孔鰕虎鱼 | 2.35 | |
| A7 | 三疣梭子蟹 | 0.84 | S7 | 脊尾白虾 | 1.47 | |
| A8 | 日本蟳 | 0.75 | S8 | 葛氏长臂虾 | 1.33 | |
| S9 | 细鳌虾 | 3.43 | ||||
| S10 | 三疣梭子蟹 | 3.71 | ||||
| S11 | 日本蟳 | 0.41 | ||||
根据生态位宽度值变化, 将春、秋2季主要游泳动物各分为三类:窄生态位种(0<Bi<1.00)、中生态位种(1.00—2.00)和广生态位种(>2.00)。秋季, 窄生态位种有鮸、口虾蛄、三疣梭子蟹和日本蟳; 中生态位种有棘头梅童鱼和葛氏长臂虾; 广生态位种有龙头鱼和安氏白虾。春季, 窄生态位种有龙头鱼、棘头梅童鱼、鮸、蓝点马鲛和日本蟳, 中生态位种有刀鲚、脊尾白虾和葛氏长臂虾, 广生态位种孔鰕虎鱼、细鳌虾和三疣梭子蟹。
2.4 生态位宽度值与相对重要性指数的关系运用SPSS 23软件分别对春、秋2季主要游泳动物的IRI和Bi指数值进行Pearson相关性分析得出, 秋季, 两者间存在显著正相关关系(r=0.897, r>0);春季, 两者之间也存在显著正相关关系(r=0.818, r>0)。运用Excel软件分别对春、秋2季主要游泳动物的IRI和Bi指数值进行回归分析, 经过多种曲线拟合, 取相关系数最大者, 可拟合得出春、秋季两者之间均存在线性关系(图 2), 即IRI值越大, Bi值也越大。

|
| 图 2 生态位宽度值与相对重要性指数值关系图 Fig. 2 The relationship between niche breath and relative importance index |
由表 3可知, 春、秋2季主要游泳动物物种间的生态位重叠不均衡。秋季, 主要游泳动物种对有28对, 生态位重叠值范围为0.08—0.88。其中, Oik>0.6的种对12对, 占总种对数的42.86%, 口虾蛄(A4)和葛氏长臂虾(A6)的重叠值最大(0.88);0.3≤Oik≤0.6的种对11对, 占39.29%;Oik<0.3的种对仅5对, 占17.86%, 棘头梅童鱼(A2)和安氏白虾(A5)的重叠值最小(0.08)。总体而言, 秋季主要游泳动物种对间的生态位重叠程度较高。春季, 主要游泳动物种对共55对, 生态位重叠值范围为0—0.85。其中Oik>0.6的种对仅12对, 占春季总种对数的21.82%, 棘头梅童鱼(S3)和蓝点马鲛(S5)的重叠值最高(0.85);0.3≤Oik≤0.6的种对有22对, 占40.00%;Oik<0.3的种对有21对, 占38.18%, 蓝点马鲛(S5)与龙头鱼(S1)、鮸(S4)、脊尾白虾(S7)和葛氏长臂虾(S8), 以及鮸(S4)与脊尾白虾(S7)的重叠值为0。总体而言, 春季主要游泳动物生态位重叠程度较低。
| 种号 Number |
A1 | A2 | A3 | A4 | A5 | A6 | A7 | 种号 Number |
S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | |
| A2 | 0.37 | S2 | 0.59 | ||||||||||||||||
| A3 | 0.13 | 0.72 | S3 | 0.26 | 0.49 | ||||||||||||||
| A4 | 0.31 | 0.73 | 0.58 | S4 | 0.04 | 0.65 | 0.10 | ||||||||||||
| A5 | 0.76 | 0.08 | 0.13 | 0.46 | S5 | 0 | 0.27 | 0.85 | 0 | ||||||||||
| A6 | 0.51 | 0.52 | 0.45 | 0.88 | 0.68 | S6 | 0.40 | 0.44 | 0.56 | 0.26 | 0.21 | ||||||||
| A7 | 0.30 | 0.20 | 0.36 | 0.61 | 0.46 | 0.59 | S7 | 0.50 | 0.13 | 0.07 | 0 | 0 | 0.21 | ||||||
| A8 | 0.62 | 0.62 | 0.77 | 0.77 | 0.54 | 0.71 | 0.74 | S8 | 0.66 | 0.74 | 0.12 | 0.61 | 0 | 0.33 | 0.26 | ||||
| S9 | 0.65 | 0.73 | 0.39 | 0.57 | 0.17 | 0.43 | 0.46 | 0.78 | |||||||||||
| S10 | 0.65 | 0.51 | 0.50 | 0.37 | 0.33 | 0.45 | 0.34 | 0.55 | 0.73 | ||||||||||
| S11 | 0.43 | 0.60 | 0.13 | 0.46 | 0 | 0.18 | 0.04 | 0.74 | 0.77 | 0.34 | |||||||||
| 该表种号对应的物种同表 2 | |||||||||||||||||||
RDA分析结果显示, 秋季, 第一排序轴的特征值为0.304, 物种与环境因子的相关系数为0.772;第二排序轴的特征值为0.119, 物种与环境因子的相关系数为0.851。蒙特卡洛显著性检验结果显示, 第一排序轴和所有排序轴均呈现不显著差异(第一轴:F= 2.181, P= 0.588;所有轴:F= 0.858, P= 0.638)。如图 3所示, 秋季主要游泳动物受盐度、温度、溶解氧和pH等环境因子影响存在生态分化现象, 第一排序轴与溶解氧及盐度均呈较大正相关, 而与温度呈最大负相关; 第二排序轴与溶解氧呈最大正相关, 而与pH呈最大负相关。2个排序轴共解释了主要游泳动物组成42.3%的变异。

春季, 第一排序轴的特征值为0.209, 物种与环境因子的相关系数为0.949。第二排序轴的特征值为0.165, 物种与环境因子的相关系数为0.872。蒙特卡洛显著性检验结果显示, 第一排序轴和所有排序轴均为不显著差异(第一轴:F= 2.042, P= 0.096;所有轴:F= 1.496, P= 0.100)。如图 3所示, 春季该海域主要游泳动物受盐度、温度、悬浮物和pH等环境因子影响存在生态分化现象。第一排序轴与盐度及悬浮物呈较大正相关, 与温度呈最大负相关。第二排序轴与悬浮物呈最强正相关, 与pH呈最强负相关。2个排序轴共解释了主要游泳动物组成45.5%的变异。
3 讨论 3.1 春、秋季物种相似性及优势种春、秋2季共捕获虾类仅为8种, 蟹类仅5种; 但2季共同种较多, 虾类6种, 而蟹类4种。2季虾类和蟹类的Jaccard’s种类相似性指数分别高达0.75和0.80, 表明2季虾、蟹类种类均为极相似, 物种更替率低, 这可能与所获虾、蟹类的生态习性有较大关系, 即舟山长白海域位于杭州湾口外, 盐度年变化幅度大, 海底泥沙量淤积厚, 为葛氏长臂虾、细巧仿对虾、三疣梭子蟹、日本蟳等广盐性甲壳类提供了适宜的栖息条件, 这与宋海棠等[29]有关这些虾蟹类适宜栖息环境条件的描述相一致。鱼类群落的2季种类数多达24种, 共同种仅11种, Jaccard’s种类相似性指数值为0.44, 表明鱼类群落为中等不相似, 这可能是鱼类群落存在明显的季节更替所致。综上所述, 2季虾蟹类和鱼类种类相似性耦合导致游泳动物群落种类总体呈现中等相似水平(Jaccard’s种类相似性指数为0.57)。
游泳动物群落春、秋季的动态变化, 导致2个季节游泳动物的优势种组成存在较大差异。秋季优势种为龙头鱼与安氏白虾, 春季优势种为孔鰕虎鱼、脊尾白虾、细鳌虾及三疣梭子蟹, 2季没有共同的优势种, 体现了群落优势种的更替, 表明游泳动物群落种类组成变化大。夏、秋季龙头鱼、安氏白虾在沿岸水域产卵、索饵育肥[29-30], 因此在尾数组成上有绝对优势, 尾数百分比分别为42.99%和41.62%, 并且龙头鱼在每个站位均有出现, 安氏白虾除了10号站位也均有出现。春、夏季为脊尾白虾、三疣梭子蟹的产卵盛期, 成熟大个体喜于河口沿岸地区产卵[29, 31], 因此春季航次脊尾白虾、三疣梭子蟹的尾数组成和出现频次均有较大优势。鮸、葛氏长臂虾和日本蟳为2季共同的重要种, 这可能与以上种类都是调查海域长期以来的春、秋季重要的经济种类, 且群体数量较大, 为近岸海域作业的重要捕捞对象有关。本文研究的主要游泳动物同张洪亮等[32]于2007年在舟山长白海域毗邻的岱衢洋海域捕获研究的种类组成相似, 但种类数却明显减少。这可能是近些年渔业资源捕捞过度, 长白岛及附近海岛船舶修造业、化工业、港口物流业等造成的海域污染在一定程度上给游泳动物群落带来负面影响。
3.2 生态位宽度春、秋2季主要游泳动物可分为窄生态位种(0—1.00)(图 4中a段)、中生态位种(1.00—2.00)(图 4中b段)和广生态位种(>2.00)(图 4中c段)三类。生态位宽度呈现较显著的分段现象(图 4)。春、秋2季每个广生态位种出现的站位数占总站位数的百分比均高达91.7%, 个体数量多且分布均匀; 每个中生态位种出现的站位数占总站位数的百分比均达50%, 数量较少; 窄生态位种总体出现的站位数占总站位数约45%, 在每个站位出现的数量很少。秋季的窄生态种口虾蛄, 其出现的站位数占总站位数的50%, 但每个站位口虾蛄出现的尾数占该站位总尾数的百分比不高于7.95%。因此, 生态位宽度与物种的种群数量和分布均有关, 这与霍红等[7]、董静瑞等[13]的研究结果也相一致。

|
| 图 4 舟山长白海域主要游泳动物生态位宽度值变化(Bi) Fig. 4 Variation of niche breadth of major nekton in Changbai |
春、秋2季主要游泳动物生态位宽度值还呈现一定的季节变化。棘头梅童鱼、葛氏长臂虾、日本蟳, 特别是龙头鱼等种类的生态位宽度在春季较秋季显著下降, 而三疣梭子蟹等种类的生态位宽度值在春季较秋季显著上升。Pearson相关性分析得到IRI与Bi指数值之间存在显著正相关, 回归分析中可得两者线性回归的相关系数较高, 拟合效果最佳, 能较客观地反映两者之间的关系, 但两者也无绝对的正向线性关系, 如春季的脊尾白虾虽为优势种, 但生态位宽度却不如一些重要种, 仅为中生态位种。这与胡成业等[15]的研究结果类似。这可能与两者的计算方法和考虑的生态特征方向不同有关, 即IRI与物种尾数、生物量及出现站位百分比等相关, 忽视了物种斑块状分布的特征, 而Bi不仅考虑了物种尾数、出现站位百分比等因素, 还综合考虑了不同站位间的分布差异性。综上所述, 在物种均衡分布的情况下, IRI值越大, Bi值也越大。
3.3 生态位重叠由于物种对同一资源有共同需要, 造成了不同物种间的生态位重叠, 因此生态位重叠在一定程度上能解释物种对资源利用的相似性和竞争性。秋季, Oik>0.6的种对占总种对数的42.86%, 生态位重叠程度总体较高。种对口虾蛄(A4)—葛氏长臂虾(A6)的生态位重叠值最大(0.88), 这与其生活习性相近, 均在沿岸、浅海的泥沙底质区域分布[30]有关, 这也表明了2个物种对资源利用相似程度高, 种间竞争大。春季, Oik<0.3的种对占38.18%, 生态位重叠程度总体较低, 表明春季物种间的生态位分化程度较高, 对资源利用的相似程度较低, 种间竞争较弱。这可能是大多数游泳动物产卵盛期在春季, 很多洄游性种类在调查海域进行产卵或产卵洄游, 与土著种类在生活、繁殖、摄食习性等方面存在差异。
生态位宽度和生态位重叠之间没有绝对正相关, 如口虾蛄(A4)—日本蟳(A8)生态位宽度均较小, 分别仅为0.67和0.75, 但种对间的重叠值较大(0.77), 这可能与其对资源利用相似程度(生活习性、摄食食性等)有关; 有些种类的生态位宽度值小, 如棘头梅童鱼(A2)—日本蟳(A8)(1.13—0.75)、龙头鱼(S1)—葛氏长臂虾(S8)(0.88—1.33), 并且物种间的资源利用相似程度也较低, 但生态位重叠值却较大, 这可能是两者间的站位重合率高所致。Wnthne等[27]研究表明生态位重叠显著的种对(Oik>0.6), 站位点的重合率也较高。另外, 春季一些种对如蓝点马鲛(S5)与龙头鱼(S1)的重叠值为0, 这并不能说明物种间为完全独立, 这可能是调查范围小, 捕获数量少且其出现的站位点不同所致, 也可能是物种对资源利用的相似度或环境适应性存在差异所致。
3.4 生态位分化冗余分析显示, 春、秋2季第一排序轴均反映了盐度、温度变化对主要游泳动物分布的直接影响, 这与史赟荣等[33]的研究结果相类似, 即盐度和温度是影响物种分布, 特别是河口沿岸区域物种分布的最主要的环境因子; 秋季的第二排序轴主要反映了溶解氧及pH的变化, 是影响秋季物种分布的重要因素, 春季的第二排序轴反映的是悬浮物及pH的变化。总体而言, 在春、秋2季的调查中也可得到pH值的变化较小(分别为8.00±0.03和8.04±0.03), 但pH是水体中一些化学性的综合体现, 与温度、盐度等环境因素耦合, 对物种产生综合影响。悬浮物是春季影响物种分布的重要因素, 这可能与春季台湾暖流的北上西进与长江冲淡水系及浙江沿岸流等交汇配置时空变化从而影响水体中的悬浮物含量, 水体的透明度存在明显的差异及不同物种生态习性有关, 如细鳌虾(S9)和日本蟳(S11)分布在高悬浮物的区域, 而刀鲚(S2)则分布在低悬浮物、低盐的区域。
通过冗余分析, 还可以对生态位宽度和生态位重叠难以解释的部分结果进行补充, 如秋季的棘头梅童鱼(A2)、日本蟳(A8)的资源利用相似程度低, 但2个物种均分布在pH值较高的区域, 具有相似的环境需求, 这就较好地解释了两者生态位宽度小而重叠值高的结果。类似地, 鮸(A3)的生态位宽度值仅为0.05, 与同在高pH区域分布的棘头梅童鱼(A2)和日本蟳(A8)有高的生态位重叠值; 春季的棘头梅童鱼(S3)和蓝点马鲛(S5)的生态位宽度值小, 两者的资源利用相似程度低, 但重叠值高(0.85), 物种出现的站位频率也不高, 但2个物种有相似的环境需求, 即均在高溶解氧区域分布。孔鰕虎鱼(S6)为广生态位种, 但与其他种类的重叠值都不高。RDA排序图显示其他种类有低溶解氧分布的特征, 而孔鰕虎鱼在高溶解氧区分布, 因此生态适应性的差异使得其与其他种类存在空间分布的差异, 从而显示出生态位分化的特征。另外, 鮸(S4)、蓝点马鲛(S5)和脊尾白虾(S7)两两之间的生态位重叠值均为0, 这与其栖息水层的差异有关, 鮸为中下层, 蓝点马鲛为中上层, 脊尾白虾则为底层, 另外鮸处在排序图的右下方, 与各环境因子的相关度均较低, 蓝点马鲛、脊尾白虾在化学需氧量、溶解氧及温度资源轴上分布差异大, 这就导致了生态位重叠值低。
| [1] | 覃林. 统计生态学. 北京: 中国林业出版社, 2009. |
| [2] | Polechová J, Storch D. Ecological Niche//Jørgensen S E, Fath B D. Encyclopedia of Ecology. Oxford: Academic Press, 2008: 1088-1097. |
| [3] | 张孝然, 蒲真, 黄治昊, 周鑫, 邢韶华. 大黄花虾脊兰生境特征及生存群落物种生态位分析. 植物科学学报, 2017, 35(6): 799–806. |
| [4] | 宗世祥, 姚国龙, 骆有庆, 许志春, 王涛. 沙棘主要蛀干害虫种群生态位. 生态学报, 2005, 25(12): 3264–3270. DOI:10.3321/j.issn:1000-0933.2005.12.022 |
| [5] | 张晶, 赵成章, 任悦, 李雪萍, 雷蕾. 张掖国家湿地公园优势鸟类种群生态位研究. 生态学报, 2018, 38(6): 2213–2220. |
| [6] | 钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561–568. |
| [7] | 霍红, 冯起, 苏永红, 司建华, 席海洋, 鱼腾飞. 额济纳绿洲植物群落种间关系和生态位研究. 中国沙漠, 2013, 33(4): 1027–1033. |
| [8] | 汤雁滨, 廖一波, 寿鹿, 曾江宁, 高爱根, 陈全震. 南麂列岛潮间带大型底栖动物群落优势种生态位. 生态学报, 2016, 36(2): 489–498. |
| [9] | 葛宝明, 郑祥, 程宏毅, 鲍毅新, 陈良. 灵昆岛围垦滩涂潮沟大型底栖动物群落和物种生态位分析. 水生生物学报, 2007, 31(5): 675–681. DOI:10.3321/j.issn:1000-3207.2007.05.011 |
| [10] | 彭松耀, 李新正, 王洪法, 张宝琳. 南黄海春季大型底栖动物优势种生态位. 生态学报, 2015, 35(6): 1917–1928. |
| [11] | 李显森, 于振海, 孙珊, 金显仕. 长江口及其毗邻海域鱼类群落优势种的生态位宽度与重叠. 应用生态学报, 2013, 24(8): 2353–2359. |
| [12] | 胡成业, 水玉跃, 田阔, 李良, 覃胡林, 张春草, 冀萌萌, 水柏年. 浙江七星列岛海洋特别保护区主要鱼类功能群划分及生态位分析. 生物多样性, 2016, 24(2): 175–184. |
| [13] | 董静瑞, 水柏年, 胡成业, 水玉跃, 杜肖, 田阔. 温州南部沿岸海域主要鱼类的生态位及种间联结性. 应用生态学报, 2017, 28(5): 1699–1706. |
| [14] | 李超男, 水玉跃, 田阔, 杜肖, 孙鹏, 董静瑞, 水柏年. 温州湾春季主要游泳动物生态位与种间联结性. 生态学报, 2017, 37(16): 5522–5530. |
| [15] | 胡成业, 水玉跃, 杜肖, 水柏年. 浙江苍南南部沿岸海域主要游泳动物生态位及种间联结性. 应用生态学报, 2015, 26(10): 3195–3201. |
| [16] | 国家海洋局第三海洋研究所, 国家海洋局第二海洋研究所. GB/T 12763. 6-2007海洋调查规范第6部分: 海洋生物调查. 北京: 中国标准出版社, 2008. |
| [17] | 陈大刚, 张美昭. 中国海洋鱼类. 青岛: 中国海洋大学出版社, 2015. |
| [18] | 朱元鼎, 张春霖, 成庆泰. 东海鱼类志. 北京: 科学出版社, 1963. |
| [19] | 赵盛龙, 徐汉祥, 钟俊生, 陈键. 浙江海洋鱼类志. 杭州: 浙江科学技术出版社, 2016. |
| [20] | 庄平, 王幼魁, 李圣法, 邓思明, 李长松, 倪勇. 长江口鱼类. 上海: 上海科学技术出版社, 2006. |
| [21] | Simpson E H. Measurement of diversity. Nature, 1949, 163(4148): 688. DOI:10.1038/163688a0 |
| [22] | Pinka E R. Ecology of the agamid lizard Amphibolurus isolepis in Western Australia. Copeia, 1971, 1971(3): 527–536. DOI:10.2307/1442450 |
| [23] | 王雪辉, 杜飞雁, 邱永松, 李纯厚, 孙典荣, 贾晓平. 1980-2007年大亚湾鱼类物种多样性、区系特征和数量变化. 应用生态学报, 2010, 21(9): 2403–2410. |
| [24] | 余世孝. 数学生态学导论. 北京: 科学技术文献出版社, 1995. |
| [25] | Shannon C E, Weaver E. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1963. |
| [26] | Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53–74. DOI:10.1146/annurev.es.04.110173.000413 |
| [27] | Wathne J A, Haug T, Lydersen C. Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress Series, 2000, 194: 233–239. DOI:10.3354/meps194233 |
| [28] | Jan L, Petr Š. Multivariate Analysis of Ecological Data Using CANOCO. New York: Cambridge University Press, 2003. |
| [29] | 宋海棠, 俞存根, 薛利建, 姚光展. 东海经济虾蟹类. 北京: 海洋出版社, 2006. |
| [30] | 郑元甲, 陈雪忠, 程家骅, 王云龙, 沈新强, 陈卫忠, 李长松. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 2003: 12–12. |
| [31] | 浙江动物志编辑委员会. 浙江动物志:甲壳类. 杭州: 浙江科学技术出版社, 1991. |
| [32] | 张洪亮, 梁君, 贺舟挺, 王伟定, 周永东. 岱衢洋渔业动物多样性分析. 水生生物学报, 2012, 36(5): 922–931. |
| [33] | 史赟荣. 长江口鱼类群落多样性及基于多元排序方法群落动态的研究[D]. 上海: 上海海洋大学, 2012. |