 2018, Vol. 38
2018, Vol. 38文章信息
- 付战勇, 孙景宽, 李传荣, 池源, 夏江宝, 屈凡柱, 宋爱云, 杨红军, 董林水.
- FU Zhanyong, SUN Jingkuan, LI Chuanrong, CHI Yuan, XIA Jiangbao, QU Fanzhu, SONG Aiyun, YANG Hongjun, DONG Linshui.
- 黄河三角洲贝壳堤土壤微生物生物量对不同生境因子的响应
- Responses of soil microbial biomass to different habitat factors in the chenier of the Yellow River Delta
- 生态学报. 2018, 38(18): 6594-6602
- Acta Ecologica Sinica. 2018, 38(18): 6594-6602
- http://dx.doi.org/10.5846/stxb201803220571
-
文章历史
- 收稿日期: 2018-03-22
- 修订日期: 2018-07-04





2. 滨州学院山东省黄河三角洲生态环境重点实验室, 滨州 256603;
3. 国家海洋局第一海洋研究所, 青岛 266061
2. Key Laboratory of Eco-environmental Science for Yellow River Delta, Binzhou University, Binzhou 256603, China;
3. The First Institute of Oceanography, State Oceanic Administration, Qingdao 266061, China
土壤微生物是陆地生态系统的主要分解者, 在土壤养分矿化、分解、固持和营养物质循环等方面起着重要作用, 同时也调控着碳和养分在土壤-植物之间的循环, 充当植物营养元素的源和库[1-3], 并且其对环境变化的敏感性通常被作为土壤环境质量的灵敏生物指标[4]。土壤微生物生物量是指土壤中体积小于5000 μm3的且去除活植物体以外的生物总量。它是土壤有机质的活性部分, 在土壤有机质中不足3%, 但却作为生态系统养分循环的关键因素, 其与土壤养分分布特征的联系对于我们理解生态系统养分循环十分重要[5]。土壤和植被是一个有机整体, 二者相辅相承、互相影响。土壤为植被生长发育提供必需的营养物质, 而植被生长又可通过调节区域小气候、凋落物来改善土壤系统的结构组成和肥力状况。在湿地生态系统中, 通过土壤微生物生物量来评价土壤肥力状况势必要考虑其土壤水分、pH值、土壤养分、盐分和土壤质地等生态限制因子, 研究这些因子对土壤微生物生物量的影响极其重要。
黄河三角洲贝壳堤是淤泥质或粉砂质海岸所特有的一种特殊类型的滩脊, 由海生贝壳及其碎片和细砂、粉砂、泥炭、淤泥质粘土薄层组成, 与美国圣路易斯安娜州和南美苏里南的贝壳堤并称为世界三大古贝壳堤[6]。近年来, 由于海水侵蚀、风暴潮、平堤水产养殖、挖砂烧瓷等自然和人为因素的影响, 该区土壤肥力严重下降, 植被退化严重。目前, 土壤微生物生物量已成为国际土壤生物学、土壤生态学以及森林生态学等研究领域的热点问题之一[7-9]。国内外学者对微生物生物量作了大量研究, 但多集中于对森林、高原、丘陵、草原和农田等生态系统的研究[10-14], 对于湿地土壤微生物生物量的研究较少[15-17], 特别是从微地形土壤的水平和垂直的空间分布角度对土壤微生物生物量的研究却鲜有报道。因此, 研究不同生境土壤微生物生物量对了解土壤养分水平及其养分的转化和循环具有重要意义, 以此来探讨影响土壤微生物生物量碳、氮、磷含量的主要土壤生境因子, 为黄河三角洲贝壳堤的土壤肥力管理、植被保护和恢复提供一定的理论依据。
1 研究地区与研究方法 1.1 研究地区概况黄河三角洲贝壳堤湿地位于渤海湾的西南部和山东省无棣县北部交汇处的滨海地区, 地理坐标为38°02′50.51″—38°21′06.06″N, 117°46′58.00″—118°05′42.95″E, 总面积约435.4 km2。该区气候为暖温带大陆性季风气候, 年平均气温为12.36℃, 多年平均降水量为550 mm, 降水集中在6—9月, 年蒸发量2430.6 mm, 蒸降比为4.4, 年日照时数为2849 h。黄河三角洲贝壳堤地势低平, 海拔一般在5 m以下; 地下水埋深浅、矿化度高; 土壤为滨海盐渍土, 成土母质主要是风积物和钙质贝壳土壤化, 土壤类别主要是滨海盐土类和贝壳砂土类; 植物以旱生、盐生灌木和草本植物为主。
1.2 研究方法 1.2.1 野外取样2017年6月, 根据黄河三角洲贝壳堤土壤质地、地貌特征以及植被覆盖率(表 1)和距离海岸的远近, 采用网格布点法, 在研究区内, 沿垂直海岸线方向设置4个采样地, 按照距海由近到远依次记作高潮线、向海侧、滩脊和背海侧4个样地。在贝壳堤剖面的0—5、5—10、10—20、20—40 cm和40—60 cm深度采集土壤样品, 取3组平行样。将采集到样品装入自封袋中密封, 装入便携式自制冰盒, 带回实验室。
| 样地 Plots |
植被类型 Vegetation types |
优势物种 Dominant plants |
伴生物钟 Non-dominant species |
总盖度/% Total cover density |
物种数 Spices |
| 高潮线High tide line | — | — | — | — | — |
| 向海侧Seaward side | 草本 | 砂引草 | 蒙古鸦葱 | 5 | 2 |
| 滩脊Dune crest | 草本×灌木 | 酸枣、芦苇、蒙古蒿 | 小果白刺、獐毛、砂引草、攀援天门冬 | 60 | 7 |
| 背海侧Landside | 草本 | 盐地碱蓬、碱蓬 | 獐毛、二色补血草 | 45 | 4 |
| 砂引草:Messerschmidia sibirica; 酸枣:Ziziphus jujuba var. spinosa; 芦苇:Phragmites australis; 蒙古蒿:Artemisia mongolica; 盐地碱蓬:Suaeda salsa; 碱蓬:Suaeda glauca; 蒙古鸦葱:Scorzonera mongolica; 小果白刺:Nitraria sibirica; 獐毛:Aeluropus sinensis; 攀援天门冬:Asparagus brachyphyllus; 二色补血草:Limonium bicolor | |||||
土壤样品放入4℃冰箱中保存。用于测定土壤微生物生物量碳、氮、磷的新鲜土样去除可见动、植物残体, 过2 mm孔径尼龙筛, 彻底混匀。在25℃+100%空气湿度的培养箱预培养7天后直接测定。用于测定土壤基本理化性质的样品在室温下自然风干, 碾磨并过100目孔径筛, 保存备用。
1.2.3 测定方法土壤微生物生物量碳、氮采用氯仿熏蒸K2SO4提取[18]。称取4份培养土样, 每一份土壤重25 g(烘干基), 其中2份用氯仿熏蒸24 h, 除去氯仿, 加入100 mL 0.5 mol/L K2SO4提取, 振荡30 min, 过滤。另2份不做氯仿熏蒸, 浸提方法同上。浸提液中有机碳含量采用TOC/TNb自动分析仪(Liquid TOC Ⅱ, 德国)测定。

|
浸提液中全氮含量采用过硫酸钾氧化-流动注射分析仪(AA3, 德国)法测定[19],
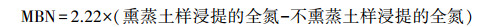
|
土壤微生物生物量磷(MBP)采用氯仿熏蒸NaHCO3提取法[20]。称取4份培养土样, 每份土壤重4.0 g(烘干基), 其中2份用氯仿熏蒸24 h, 除去氯仿, 用0.5 mol/L NaHCO3 (pH 8.5)提取, 另外2份不做氯仿熏蒸, 浸提方法同上。采用比色法测定分析提取液中的磷。同时用外加无机磷的方法测定磷的提取回收率, 以熏蒸土样与不熏蒸土样提取的P的差值并校正提取回收率后, 乘以转换系数Kp计算土壤微生物生物量磷。

|
式中, Epi:熏蒸与未熏蒸的差值; Rpi:=[(外加KH2PO4溶液土壤的测定值-未熏蒸土壤的测定值)/25]·100%;Kp:转化系数, 取值0.4。
土壤有机碳(SOC)采用重铬酸钾-浓硫酸外加热法[21]; 土壤全氮(TN)用元素分析仪(varioEL Ⅲ, Elementar, 德国)测定; 土壤全磷(TP)采用钼蓝比色法测定。
土壤含水量采用烘干法测定, 土壤含盐量采用重量法测定, pH值采用电位法测定。
1.3 数据分析数据处理采用SPSS 17.0和Microsoft Excel 2003进行统计分析, 采用单因素方差分析(One-way ANVOA)比较不同生境不同土层土壤微生物生物量碳、氮、磷的差异性, 并采用Duncan法进行多重比较并判断测定指标差异是否显著; 土壤微生物生物量碳、氮、磷与土壤生境因子的相关分析采用Pearson检验; 采用Origin 8.6软件绘图。
2 结果 2.1 不同生境的基本理化性质不同微地形之间环境因子存在差异(表 2)。不同生境的pH范围在7.88—9.07之间, 表现为向海侧>高潮线>滩脊>背海侧。含盐量的变化范围在1.63—13.21 g/kg之间, 表现为背海侧>高潮线>滩脊>向海侧。4个样地的含水量在5.37%—17.87%之间, 表现为高潮线>背海侧>滩脊>向海侧。从土壤养分含量来看, 土壤有机碳、全氮、全磷含量均表现出:滩脊>背海侧>高潮线>向海侧。
| 样地 Plots |
pH | 含盐量/(g/kg) Salt content |
含水量/% Water content |
土壤有机碳/(g/kg) Soil organic carbon |
全氮/(g/kg) Total nitrogen |
全磷/(g/kg) Total phosphorus |
| 高潮线High tide line | 7.94±0.07 | 7.09±0.39 | 17.87±2.27 | 1.11±0.18 | 0.12±0.02 | 0.31±0.02 |
| 向海侧Seaward side | 9.07±0.07 | 1.63±0.31 | 5.37±1.37 | 0.56±0.08 | 0.06±0.01 | 0.25±0.02 |
| 滩脊Dune crest | 8.37±0.27 | 5.30±3.70 | 7.66±2.46 | 5.85±5.47 | 0.62±0.57 | 0.43±0.14 |
| 背海侧Landside | 7.88±0.20 | 13.21±1.63 | 14.06±1.95 | 2.56±2.01 | 0.22±0.22 | 0.38±0.10 |
黄河三角洲贝壳堤土壤微生物生物量碳、氮、磷含量见图 1。不同生境因子影响下, 土壤MBC、MBN、MBP含量存在显著差异, 表现为滩脊>背海侧>高潮线>向海侧。4个样地土壤MBC、MBN、MBP的变化范围分别在7.19—347.21, 1.26—62.57, 2.31—17.29 mg/kg之间。

|
| 图 1 不同生境土壤微生物生物量碳、氮、磷的空间分布 Fig. 1 Spatial distribution of MBC, MBN and MBP in different habitats 小写字母表示同一生境不同土层中土壤微生物生物量碳、氮、磷的差异性; 大写字母表示同一土层不同生境中土壤微生物生物量碳、氮、磷的差异性 |
如表 3所示, 土壤微生物生物量碳、氮、磷占土壤有机碳、全氮、全磷比例变化范围分别为1.09%—3.48%、2.62%—7.27%、0.78%—2.86%;变异系数分别为5.73%—11.60%、6.56%—10.36%、3.09%—9.28%。4个样地MBC/SOC、MBN/TN、MBP/TP所占比例趋势不同, MBC/SOC均值表现为高潮线>背海侧>滩脊>向海侧; MBN/TN均值表现为背海侧>滩脊>高潮线>向海侧; MBP/TP均值表现为滩脊>背海侧>向海侧>高潮线。其中, 向海侧MBC/SOC、MBN/TN最低。
| 样地 Plots |
微生物生物量碳/土壤有机碳 MBC/SOC/% |
微生物生物量氮/全氮 MBN/TN/% |
微生物生物量磷/全磷 MBP/TP/% |
|||||||||||
| 最小值 Min |
最大值 Max |
平均值 Average |
变异系数 C.V/% |
最小值 Min |
最大值 Min |
平均值 Average |
变异系数 C.V/% |
最小值 Min |
最大值 Max |
平均值 Average |
变异系数 C.V/% |
|||
| 高潮线High tide line | 1.72 | 3.38 | 2.8 | 10.95 | 3.35 | 4.32 | 3.95 | 6.82 | 0.94 | 1.49 | 1.25 | 3.09 | ||
| 向海侧Seaward side | 1.09 | 2.00 | 1.58 | 11.60 | 2.62 | 5.00 | 3.93 | 6.56 | 0.78 | 1.78 | 1.26 | 9.28 | ||
| 滩脊Dune crest | 2.13 | 3.28 | 2.63 | 5.73 | 4.50 | 7.27 | 5.64 | 7.79 | 1.55 | 2.86 | 2.31 | 7.41 | ||
| 背海侧Landside | 1.82 | 3.48 | 2.74 | 8.90 | 3.41 | 6.74 | 5.70 | 10.36 | 1.25 | 2.79 | 2.13 | 5.86 | ||
| MBC/SOC:the ratio of microbial biomass carbon to soil organic carbon; MBN/TN:the ratio of microbial biomass nitrogen to total nitrogen; MBP/TP:the ratio of microbial biomass phosphorus to total phosphorus; C.V:coefficient of variation | ||||||||||||||
随着土壤深度的增加, 不同生境土壤微生物生物量碳、氮、磷含量变化趋势基本一致(图 1), 均呈现出垂直分布特征, 即:0—5 cm>5—10 cm>10—20 cm>20—40 cm>40—60 cm。其中, 滩脊和背海侧MBC、MBN含量在0—60 cm各土层上差异显著(P<0.05)。在土壤深度为0—20 cm时, 滩脊和背海侧处土壤MBC、MBN含量显著大于高潮线和向海侧(P<0.05)。当土壤深度为20—40 cm时, 滩脊处土壤MBC、MBN显著大于高潮线、向海侧和背海侧土壤MBC、MBN。4个样地的土壤MBP则相对稳定, 在0—20 cm各土层间均差异显著(P<0.05), 当土壤深度为20—60 cm时, 各土层间均无显著性差异(P>0.05)。
2.4 土壤微生物生物量碳、氮、磷和土壤碳、氮、磷的相关性相关分析表明, 不同生境中, 滩脊、背海侧和高潮线处土壤MBC和MBN、MBC和MBP、MBN和MBP之间均具有极显著正相关(P<0.01)。向海侧土壤MBC和MBN之间呈显著正相关(P<0.05), MBN和MBP呈极显著正相关(P<0.01)。滩脊和背海侧土壤MBC、MBN、MBP和SOC、TN、TP之间呈极显著正相关(P<0.01)。高潮线处土壤MBC、MBN、MBP和TN、TP之间呈极显著正相关(P<0.01), MBC与SOC呈显著正相关(P<0.05)。向海侧MBN、MBP与TN呈极显著正相关(P<0.01), MBC、MBN、MBP与SOC、TP无显著相关性(P>0.05)(表 4)。
| 样地 Plots |
土壤有机碳 SOC |
总氮 TN |
总磷 TP |
微生物生物量碳 MBC |
微生物生物量氮 MBN |
微生物生物量磷 MBP |
|
| 高潮线High tide line | SOC | 1 | |||||
| TN | 0.583* | 1 | |||||
| TP | 0.336 | 0.833** | 1 | ||||
| MBC | 0.564* | 0.757** | 0.688** | 1 | |||
| MBN | 0.462 | 0.943** | 0.782** | 0.743** | 1 | ||
| MBP | 0.420 | 0.702** | 0.718** | 0.932** | 0.680** | 1 | |
| 向海侧Seaward side | SOC | 1 | |||||
| TN | -0.336 | 1 | |||||
| TP | 0.063 | 0.356 | 1 | ||||
| MBC | 0.354 | 0.466 | 0.005 | 1 | |||
| MBN | 0.041 | 0.647** | -0.112 | 0.769* | 1 | ||
| MBP | -0.485 | 0.818** | 0.294 | -0.003 | 0.807** | 1 | |
| 滩脊Dune crest | SOC | 1 | |||||
| TN | 0.997** | 1 | |||||
| TP | 0.958** | 0.966** | 1 | ||||
| MBC | 0.980** | 0.983** | 0.987** | 1 | |||
| MBN | 0.975** | 0.976** | 0.987** | 0.995** | 1 | ||
| MBP | 0.941** | 0.944** | 0.981** | 0.974** | 0.949** | 1 | |
| 背海侧Landside | SOC | 1 | |||||
| TN | 0.996** | 1 | |||||
| TP | 0.897** | 0.906** | 1 | ||||
| MBC | 0.992** | 0.993** | 0.901** | 1 | |||
| MBN | 0.989** | 0.994** | 0.893** | 0.991** | 1 | ||
| MBP | 0.924** | 0.934** | 0.972** | 0.945** | 0.929** | 1 | |
| n为样本数, **表示相关性极显著(P<0.01), *代表相关性显著(P<0.05);SOC:soil organic carbon; TN:total nitrogen; TP:total phosphorus; MBC:microbial biomass carbon; MBN:microbial biomass nitrogen; MBP:microbial biomass phosphorus | |||||||
由表 5所知, 除向海侧微生物生物量碳外, 滩脊、高潮线和背海侧土壤MBC、MBN、MBP均与pH值呈极显著负相关(P<0.01)。滩脊和背海侧土壤MBC、MBN、MBP与含盐量、含水量呈显著或极显著正相关。高潮线土壤MBC、MBN、MBP与土壤含水量呈极显著负相关(P<0.01), 土壤MBC与土壤含盐量呈显著正相关(P<0.05)。向海侧土壤MBN、MBP与土壤含水量呈极显著正相关(P<0.01)。
| 样地 Plots |
土壤微生物生物量 Soil microbial biomass |
含盐量 Salt content |
pH | 含水量 Water content |
| 高潮线High tide line | MBC | 0.607* | -0.688** | -0.901** |
| MBN | 0.593* | - 0.727** | -0.754** | |
| MBP | 0.473 | -0.699** | -0.939** | |
| 向海侧Seaward side | MBC | 0.253 | -0.214 | 0.470 |
| MBN | 0.320 | -0.622** | 0.645** | |
| MBP | 0.434 | -0.811** | 0.794** | |
| 滩脊Dune crest | MBC | 0.983** | -0.919** | 0.533* |
| MBN | 0.976** | -0.925** | 0.525* | |
| MBP | 0.962** | -0.943** | 0.525* | |
| 背海侧Landside | MBC | 0.733** | -0.915** | 0.752** |
| MBN | 0.749** | -0.901** | 0.748** | |
| MBP | 0.731** | -0.973** | 0.586** | |
| n为样本数, **表示相关性极显著(P<0.01), *代表相关性显著(P<0.05) | ||||
在湿地生态系统中, 不同微地形中土壤水分、pH值、土壤含盐量等环境因子的变化都可能引起土壤微生物生物量的变化, 研究这些因子对土壤微生物生物量的影响极其重要[22-23]。微生物生物量与土壤水分关系的研究报道较多, 但至今仍无确定性结论。本研究结果表明, 向海侧、滩脊、背海侧区域土壤微生物生物量碳、氮、磷与土壤含水量呈显著或极显著正相关(表 5), 这与Devi等[10]和赵彤等[24-25]的研究结果一致, 说明适宜的水分条件既满足了微生物生长的需求, 也没有对土壤通气性造成阻碍, 因此适合于微生物的生长繁殖。但高潮线区域土壤微生物生物量碳、氮、磷与土壤含水量呈负相关关系, 这可能是由于高潮线距离海洋较近, 且地下水位很浅(<0.6 m), 受到不规则半日潮的影响, 交替性受到海水冲刷, 潮汐作用减少了土壤氧气含量, 从而影响微生物活性, 这一结论也得到证实[26-28]。
在黄河三角洲贝壳堤, 土壤含盐量和pH是反映土壤状况的重要参数。本研究中, 除向海侧微生物碳外, 其余生境中的微生物生物量碳和4个生境中的微生物生物量氮、磷含量均与pH值呈显著或极显著负相关, 说明了土壤pH值的升高不利于土壤微生物的活性, 从而导致微生物数量减少。目前, 盐分对土壤微生物的影响仍然存在争议, Wong等[23]研究表明高盐条件下土壤微生物生物量是低盐条件下的3倍多; Tripathi等、李玲等和操庆等[29-31]研究表明高的含盐量反而抑制土壤微生物生物量。本研究中, 盐度最低(1.63 g/kg)的向海侧区域土壤微生物生物量碳、氮、磷含量最低, 含盐量较高(5.30—13.21 g/kg)的背海侧、滩脊区域土壤微生物生物量碳、氮、磷含量达到较高水平, 且该地的微生物生物量和含盐量呈显著正相关, 这可能是由于滩脊和背海侧区域表层土壤堆积丰富的有机物质, 0—60 cm土层的有机碳均值分别为5.58 g/kg、2.56 g/kg, 且0—5 cm表层土壤有机碳含量分别为14.94 g/kg、5.82 g/kg, 土壤通过增加水解有机质以此来降低高盐生境对微生物种群的胁迫。因此, 土壤含水量、pH值和土壤含盐量均是影响贝壳堤土壤微生物生物量的限制性因子。
3.2 不同生境养分对微生物生物量的影响土壤微生物生物量是活的土壤有机质组分, 在土壤有机质中不足3%, 但却为土壤中能量循环和养分转化提供了一定动力, 其含量高而周转率较低时作为养分“库”, 含量低而周转率较高时作为养分“源”[32]。植被类型和枯落物覆盖是影响土壤微生物的主要因素。土壤微生物生物量作为重要的调控土壤源和库的因子, 其易受植被类型的影响。植被类型主要通过覆盖在土壤表面的枯落物层来改善微地形小气候, 影响土壤养分含量, 促进生态系统的养分循环速率[33-34]。除此物理作用外, 枯落物分解提供的碳源也是微生物生存和繁殖需要的能量。植被类型的变化导致凋落物产量和积累量不同, 从而影响归还入土壤的养分含量。本研究中, 植被覆盖度较高的滩脊(60%)和背海侧(45%)地表有一定的枯落物层, 枯落物层有利于蓄积雨水, 减少土壤表层水分蒸发, 为微生物的生长、繁殖提供了大量的碳源。微生物可从土壤中同化有机碳、氮、磷等养分作为自身营养需要, 最终将它们同化为微生物体碳、氮、磷, 从而提高了滩脊和背海侧土壤微生物生物量碳、氮、磷含量, 表明滩脊和背海侧对土壤微生物生物量的维持能力较强。
研究表明, 土壤微生物生物量所占土壤养分的比例关系能够反映土壤养分向土壤微生物生物量的转化效率和固碳能力大小[35]。本研究中, 土壤微生物生物量碳、氮、磷占土壤有机碳、全氮、全磷百分比变化范围分别为1.09%—3.48%、2.62%—7.27%、0.78%—2.86%, 与已有研究结果0.65%—7.24%、0.93%—7.41%、0.16%—7.6%相一致[10, 17, 25]。本研究结果表明滩脊、背海侧和高潮线处MBC/SOC无显著差异, 但显著高于向海侧MBC/SOC。土壤MBN/TN、MBP/TP的变化趋势为滩脊、背海侧>向海侧、高潮线, 这可能与植被覆盖率(表 1)以及输入有机物质和养分的数量有关, 造成不同生境中微生物种类和数量差异, 导致不同生境中微生物生物量之间存在一定差异。同时, 不同生境间的有机质积累和分解速率也会影响土壤中氮磷含量。滩脊、背海侧植被覆盖率较高, 占45%以上, 主要分布着地上生物量较大的酸枣、芦苇、蒙古蒿、碱蓬、盐地碱蓬等植被, 而向海侧则植被覆盖率较低, 仅占5%左右, 且生长的砂引草和蒙古鸦葱地上生物量较小, 这也直接导致了滩脊和背海侧输入土壤有机物质的数量和质量要优于向海侧和高潮线区域, 且滩脊和背海侧土壤微生物活性和土壤微生物生物量较高, 促进了土壤有机质的分解和转化, 因此MBN/TN和MBP/TP高于向海侧和高潮线。
本研究中, 土壤含水量、pH值、植被类型均影响了土壤微生物生物量, 随着土壤深度的增加, 微生物生物量碳、氮、磷均呈现下降趋势, 这一结论与张静等[16]、吴建平等[36]的研究结论相符。除了向海侧微生物生物量碳、磷外, 不同生境间土壤微生物生物量碳、氮、磷之间均呈现显著或极其显著相关, 这与赵彤等[25]的研究结果一致, 同时也说明了土壤微生物本身的生物量大小决定土壤微生物对氮素、磷素的转化和固持能力[37]。滩脊和背海侧土壤微生物生物量碳、氮、磷含量均与土壤有机碳、全氮、全磷之间具有显著相关性, 且协同性和稳定性高, 这与吴建平等[36]和张海燕等[38]的结果一致, 表明土壤微生物生物量碳、氮、磷含量可以作为判断不同微地形间土壤肥力状况的生物学指标。
4 结论不同生境因子对土壤微生物生物量碳、氮、磷分布有一定的影响, 即滩脊>背海侧>高潮线>向海侧, 且具有垂直分布特征:0—5 cm>5—10 cm>10—20 cm>20—40 cm>40—60 cm。滩脊和背海侧土壤微生物碳、氮、磷的非生物限制性因子为含水量、pH值、含盐量; 向海侧和高潮线区域土壤微生物碳、氮、磷的非生物限制性因子为含水量和pH值。滩脊、背海侧和高潮线土壤微生物生物量碳、氮、磷及土壤养分间的相关显著或者极其显著, 表明土壤微生物生物量碳、氮、磷可以作为判断土壤肥力状况的生物学指标。
| [1] | Ruan H H, Zou X M, Scatena F N, Zimmerman J K. Asynchronous fluctuation of soil microbial biomass and plant litterfall in a tropical wet forest. Plant and Soil, 2004, 260(1/2): 147–154. DOI:10.1023/B:PLSO.0000030177.20951.94 |
| [2] | 周正虎, 王传宽. 生态系统演替过程中土壤与微生物碳氮磷化学计量关系的变化. 植物生态学报, 2016, 40(12): 1257–1266. DOI:10.17521/cjpe.2016.0218 |
| [3] | Leff J W, Jones S E, Prober S M, Barberán A, Borer E T, Firn J L, Harpole W S, Hobbie S E, Hofmockel K S, Knops J M, McCulley R L, La Pierre K, Risch A C, Seabloom E W, Schütz M, Steenbock C, Stevens C J, Fierer N. Consistent responses of soil microbial communities to elevated nutrient inputs in grasslands across the globe. Proceedings of the National Academy of Sciences of the United States of America, 2015, 112(35): 10967–10972. DOI:10.1073/pnas.1508382112 |
| [4] | An S S, Mentler A, Acosta-Martínez V, Blum W E H. Soil microbial parameters and stability of soil aggregate fractions under different grassland communities on the Loess Plateau, China. Biologia, 2009, 64(3): 424–427. |
| [5] | Hall E K, Maixner F, Franklin O, Daims H, Richter A, Battin T. Linking microbial and ecosystem ecology using ecological stoichiometry:a synthesis of conceptual and empirical approaches. Ecosystems, 2011, 14(2): 261–273. DOI:10.1007/s10021-010-9408-4 |
| [6] | 田家怡, 夏江宝, 孙景宽. 黄河三角洲贝壳堤岛生态保护与恢复. 北京: 化学工业出版社, 2011. |
| [7] | Edwards K A, McCulloch J, Kershaw G P, Jefferies R L. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biology and Biochemistry, 2006, 38(9): 2843–2851. DOI:10.1016/j.soilbio.2006.04.042 |
| [8] | Baldrian P, Merhautová V, Petránková M, Cajthaml T, Šnajdr J. Distribution of microbial biomass and activity of extracellular enzymes in a hardwood forest soil reflect soil moisture content. Applied Soil Ecology, 2010, 46(2): 177–182. DOI:10.1016/j.apsoil.2010.08.013 |
| [9] | Foote J A, Boutton T W, Scott D A. Soil C and N storage and microbial biomass in US southern pine forests:influence of forest management. Forest Ecology and Management, 2015, 355: 48–57. DOI:10.1016/j.foreco.2015.03.036 |
| [10] | Devi N B, Yadava P S. Seasonal dynamics in soil microbial biomass C, N and P in a mixed-oak forest ecosystem of Manipur, North-east India. Applied Soil Ecology, 2006, 31(3): 220–227. DOI:10.1016/j.apsoil.2005.05.005 |
| [11] | 李国辉, 陈庆芳, 黄懿梅, 安韶山, 郑粉莉, 陈利顶. 黄土高原典型植物根际对土壤微生物生物量碳、氮、磷和基础呼吸的影响. 生态学报, 2010, 30(4): 976–983. |
| [12] | 吴永胜, 马万里, 李浩, 吕桂芬, 卢萍. 内蒙古退化荒漠草原土壤有机碳和微生物生物量碳含量的季节变化. 应用生态学报, 2010, 21(2): 312–316. |
| [13] | 臧逸飞, 郝明德, 张丽琼, 张昊青. 26年长期施肥对土壤微生物量碳、氮及土壤呼吸的影响. 生态学报, 2015, 35(5): 1445–1451. |
| [14] | 王宝荣, 杨佳佳, 安韶山, 张海鑫, 白雪娟. 黄土丘陵区植被与地形特征对土壤和土壤微生物生物量生态化学计量特征的影响. 应用生态学报, 2018, 29(1): 247–259. |
| [15] | 裴希超, 许艳丽, 魏巍. 湿地生态系统土壤微生物研究进展. 湿地科学, 2009, 7(2): 181–186. |
| [16] | 张静, 马玲, 丁新华, 陈旭日, 马伟. 扎龙湿地不同生境土壤微生物生物量碳氮的季节变化. 生态学报, 2014, 34(13): 3712–3719. |
| [17] | 彭佩钦, 吴金水, 黄道友, 汪汉林, 唐国勇, 黄伟生, 朱奇宏. 洞庭湖区不同利用方式对土壤微生物生物量碳氮磷的影响. 生态学报, 2006, 26(7): 2261–2267. DOI:10.3321/j.issn:1000-0933.2006.07.027 |
| [18] | 吴金水, 林启美, 黄巧云. 土壤微生物生物量测定方法及其应用. 北京: 气象出版社, 2006. |
| [19] | 姚槐应, 黄昌勇. 土壤微生物生态学及其实验技术. 北京: 科学出版社, 2006. |
| [20] | Brookes P C, Powlson D S, Jenkinson D S. Measurement of microbial biomass phosphorus in soil. Soil Biology and Biochemistry, 1982, 14(4): 319–329. DOI:10.1016/0038-0717(82)90001-3 |
| [21] | 中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科学出版社, 1978. |
| [22] | Bastida F, Barberá G G, García C, Hernández T. Influence of orientation, vegetation and season on soil microbial and biochemical characteristics under semiarid conditions. Applied Soil Ecology, 2008, 38(1): 62–70. DOI:10.1016/j.apsoil.2007.09.002 |
| [23] | Wong V N L, Dalal R C, Greene R S B. Salinity and sodicity effects on respiration and microbial biomass of soil. Biology and Fertility of Soils, 2008, 44(7): 943–953. DOI:10.1007/s00374-008-0279-1 |
| [24] | 何容, 王国兵, 汪家社, 许波峰, 汪科继, 方燕鸿, 施政, 阮宏华. 武夷山不同海拔植被土壤微生物量的季节动态及主要影响因子. 生态学杂志, 2009, 28(3): 394–399. |
| [25] | 赵彤, 闫浩, 蒋跃利, 黄懿梅, 安韶山. 黄土丘陵区植被类型对土壤微生物量碳氮磷的影响. 生态学报, 2013, 33(18): 5615–5622. |
| [26] | Wang J J, Vollrath S, Behrends T, Bodelier P L E, Muyzer G, Meima-Franke M, Den Oudsten F, Van Cappellen P, Laanbroek H J. Distribution and diversity of Gallionella-like neutrophilic iron oxidizers in a tidal freshwater marsh. Applied and Environmental Microbiology, 2011, 77(7): 2337–2344. DOI:10.1128/AEM.02448-10 |
| [27] | 赵先丽, 周广胜, 周莉, 吕国红, 贾庆宇, 谢艳兵. 盘锦芦苇湿地凋落物土壤微生物量碳研究. 农业环境科学学报, 2007, 26(增刊): 127–131. |
| [28] | 刘银银, 李峰, 孙庆业, 谢永宏. 湿地生态系统土壤微生物研究进展. 应用与环境生物学报, 2013, 19(3): 547–552. |
| [29] | Tripathi S, Kumari S, Chakraborty A, Gupta A, Chakrabarti K, Bandyapadhyay B K. Microbial biomass and its activities in salt-affected coastal soils. Biology and Fertility of Soils, 2006, 42(3): 273–277. DOI:10.1007/s00374-005-0037-6 |
| [30] | 王帅, 李玲, 付战勇, 赵珍珍, 郭锐. 施肥对黄河三角洲区盐碱化土壤活性碳、氮的影响. 农业现代化研究, 2014, 35(6): 804–809. |
| [31] | 操庆, 曹海生, 魏晓兰, 孙玮, 吴彩姣, 谷勋刚. 盐胁迫对设施土壤微生物量碳氮和酶活性的影响. 水土保持学报, 2015, 29(4): 300–304. |
| [32] | Perelo L W, Munch J C. Microbial immobilisation and turnover of 13C labelled substrates in two arable soils under field and laboratory conditions. Soil Biology and Biochemistry, 2005, 37(12): 2263–2272. DOI:10.1016/j.soilbio.2005.02.039 |
| [33] | Fanin N, Fromin N, Buatois B, Hättenschwiler S. An experimental test of the hypothesis of non-homeostatic consumer stoichiometry in a plant litter-microbe system. Ecology Letters, 2013, 16(6): 764–772. DOI:10.1111/ele.12108 |
| [34] | Sayer E J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biological Reviews, 2006, 81(1): 1–31. DOI:10.1086/504022 |
| [35] | Marschner P, Kandeler E, Marschner B. Structure and function of the soil microbial community in a long-term fertilizer experiment. Soil Biology and Biochemistry, 2003, 35(3): 453–461. DOI:10.1016/S0038-0717(02)00297-3 |
| [36] | 吴建平, 韩新辉, 许亚东, 任成杰, 杨改河, 任广鑫. 黄土丘陵区不同植被类型下土壤与微生物C, N, P化学计量特征研究. 草地学报, 2016, 24(4): 783–792. |
| [37] | 何振立. 土壤微生物量及其在养分循环和环境质量评价中的意义. 土壤, 1997, 29(2): 61–69. |
| [38] | 张海燕, 肖延华, 张旭东, 李军, 席联敏. 土壤微生物量作为土壤肥力指标的探讨. 土壤通报, 2006, 37(3): 422–425. DOI:10.3321/j.issn:0564-3945.2006.03.002 |