 2018, Vol. 38
2018, Vol. 38文章信息
- 苏昌祥, 钟稚昉, 鲁长虎.
- SU Changxiang, ZHONG Zhifang, LU Changhu.
- 动物在大别山五针松种群天然更新中的作用
- Role of animals in the natural population regeneration of Pinus dabeshanensis
- 生态学报. 2018, 38(17): 6194-6203
- Acta Ecologica Sinica. 2018, 38(17): 6194-6203
- http://dx.doi.org/10.5846/stxb201710281933
-
文章历史
- 收稿日期: 2017-10-28
- 网络出版日期: 2018-05-30





全球大约有110余种松属(Pinus)植物[1], 绝大多数松属植物都是依靠风传播种子, 此外, 约有20余种松属植物种子球果硕大、种子饱满, 无翅或具没有功能的短翅, 球果成熟后种鳞闭合或者部分张开, 但种子不会轻易脱落, 无法借助风力传播, 主要依靠动物进行传播[2]。动物传播松属植物种子的主要传播途径是贮食传播(synzoochory), 其贮食行为十分复杂, 主要包括动物寻找、选择植物种子, 将种子搬运、埋藏在贮藏地以及再次将种子挖出取食等一系列行为[3]。被贮藏的植物种子如果未被重新取食, 便有机会萌发而得以传播。国内外对此均有报道, 美洲白皮松(Pinus albicaulis)主要依靠加州星鸦(Nucifraga columbiana)、红胸(Sitta canadensis)、红松鼠(Tamiasciuris hudsonicus)和花鼠(Eutamias sibiricus)等4种动物贮藏、传播[4-5];我国东北的红松(Pinus koraiensis)种子是多种动物的食物, 其中星鸦(Nucifraga caryocatactes)、松鼠(Sciurius europaeus)、花鼠和普通(Sitta europaea)具有贮食行为, 是其主要的种子传播者[6-7];华山松(Pinus armandii)也依赖啮齿动物和星鸦进行种子传播[8-9]。
大别山五针松(Pinus dabeshanensis), 为松科松属常绿乔木, 是我国现存的几种大种子果松之一[6], 为我国特有种, 其天然分布植株较少且分布范围狭窄, 仅分布于皖鄂豫三省交界的大别山区, 安徽省岳西县河图镇大王沟是目前已知最大的大别山五针松天然种群集中分布地, 种群更新困难, 已被列为国家二级重点保护野生植物。针对大别山五针松天然更新不力的情况, 目前已有较多研究对此进行分析:结实母株罕见, 授粉率极低[10];球果、种子松结实率较低、败育率极高, 种子活力不足, 发芽率极低[11-12];病虫害也影响到大别山五针松植株存活[13]。关于动物与大别山五针松的关系上, 很多研究认为营养丰富的种子使其球果、种子遭受鸟类、小型啮齿类等动物捕食危害, 野猪(Sus scrofa)等动物在林中取食果球和种子, 觅食时会破坏大别山五针松的幼树、幼苗, 极大地影响了大别山五针的天然更新[11, 14]。虽然这些研究取得了较多的成果, 但动物在其天然更新中的作用过程尚不清楚。
由于大别山五针松种子无种翅, 籽粒饱满可供动物食用, 具有典型的动物传播性状。本文以安徽岳西大王沟大别山五针松天然种群为研究对象, 拟回答以下问题:(1)大别山五针松结实情况以及球果、种子质量如何?是否有优质的成熟种子进入土壤种子库?(2)取食种子的动物种类有哪些?哪些动物具有潜在的传播作用?(3)研究地大别山五针松种群是否具有有效的天然幼苗更新?动物在其更新中是否起到重要作用?通过以上研究, 综合分析其天然更新不力的原因。
1 研究地与研究方法 1.1 研究地自然概况研究地位于安徽省安庆市岳西县河图镇南河村境内大王沟(116°5′27.6″—116°5′42″E, 30°49′15.6″—30°49′33.5″N), 地处大别山脉腹地, 海拔1050—1200 m, 山高坡陡, 岩层主体由花岗岩、片麻岩等构成, 主要土壤为山地黄棕壤和山地棕壤, 以山地棕壤为主, 质地疏松, pH值为5—5.5。大王沟属北亚热带大陆型温暖湿润季风气候, 光照充足, 夏季温湿, 冬季寒冷, 具有典型的山地气候特征, 年平均气温14—15℃;无霜期约为210 d;雨量充沛, 年降水量1200—1400 mm;相对湿度超过76%[15]。
研究地是我国大别山五针松最大的天然种群集中分布地, 自然种群天然分布于针阔混交林内, 伴生乔木有黄山松(Pinus taiwanensis)、短柄枹(Quercus serrata)、茅栗(Castanea seguinii)、灯台树(Bothrocaryum controversum)、映山红(Rhododendron simsii)、满山红(Rhododendron mariesii)、湖北四照花(Dendrobenthamia hupehensis)等;林下灌木层主要有伞八仙(Hydrangea umbellata)、山橿(Lindera reflexa)、菝葜(Smilax china)以及部分乔木幼苗等;草本层植物种类较少, 主要以禾本科的野青茅(Deyeuxia arundinacea)、求米草(Oplismentls undulatifolius)、荩草(Arthraxon hispidus)及莎草科的宽叶苔草(Carex siderosticta)为主[16-17]。
1.2 球果和种子特征2015年11月上旬采集研究地成熟大别山五针松球果, 选取结实量较大且易采集的645、646、648、660四株母树作为目标树(为便于保护, 岳西县林业局对本地分布较大的野生大别山五针松植株进行编号, 本研究采用此编号), 所选母树均为高度10 m左右的成年结实个体, 其中母树645、646生长于阴坡处, 为东坡向, 所处地带坡度较为平缓;母树660亦生长于阴坡处, 为东北坡向, 毗邻山脊, 地势陡峭, 球果采摘难度较大。另外, 由于大别山五针松多集中生长于阴坡近山脊处, 阳坡结实母树极少, 且多处于陡峭崖壁处, 选取可采摘母树648, 为西北坡向, 但该处极为陡峭, 不利于球果采摘。
根据不同母树结实情况以及采集难度不同, 母树645采集10枚球果(n=10), 646采集18枚球果(n=18), 648采集2枚球果(n=2), 660采集5枚球果(n=5)。记录球果所属母树的编号、霉变情况, 阴干保存。测量采集球果长度、直径, 分离种子, 水沉法下沉挑选出饱满种子, 记录种子数和饱满种子数, 使用Excel计算空瘪率。
空瘪率=(种子数-饱满种子数)/种子数×100%。
1.3 种子雨与土壤种子库于2016年10—11月进行种子雨及土壤种子库调查。10月初大别山五针松球果种子成熟, 分别645、646、666、667四株母树树冠范围内(四株母树均生长于阴坡相对平缓处, 结实量较大, 易于收集), 各布置2个1 m×1 m自制种子雨收集网, 网体为尼龙材质, 网眼直径3 mm, 网面四角用结实的树枝架起, 平行于水平面, 中心部分稍向下垂呈浅兜状, 最凹处离地约1 m, 以防止鼠类破坏。10—11月中旬期间, 每隔5天观察记录种子掉落情况。
11月中旬, 进行土壤种子库调查。在四株母树所在区域距母树树干0—5、5—10、10—15 m范围随机选取5块20 cm×20 cm小样方, 共计60个小样方。每隔5 d对每个小样方上层枯枝落叶层及下方土壤层0—10 cm范围, 使用小铲翻动, 观察、计数大别山五针松种子个数, 并记录啃食情况, 共进行3轮调查。
1.4 访问动物种类选取结实量较大且利于观测的645、646、667三株母树作为观测对象, 于2015年11—12月、2016年10—11月, 使用双筒望远镜(Bosma7×50ZCY)对大别山五针松母树及其附近观察, 记录来访动物种类、数量、取食行为等信息, 期间进行10次全天候观查(6:30—17:00)。同时, 对研究地随机母株周围布置鼠笼, 鉴定、记录捕捉到的啮齿动物种类并拍照。查阅相关文献并走访当地群众, 分析大别山五针松球果及种子被捕食情况。
1.5 地面种子摆放实验及埋藏实验于2016年10—11月份, 选择研究点采集的大别山五针松种子, 进行摆放实验, 选取母树667生长处阴坡枯落物区、母树666生长处阴坡裸岩区、母树648生长处阳坡枯落物区以及母树648上坡处阳坡裸岩区4个地点作为摆放实验地点, 实验期间, 于13:00点—14:00点在4个摆放点分别摆放10、20、30、40、50、100、150、200粒种子(各组n=4), 共32次(点)实验, 隔天观察种子完整、当场取食、消失的个数并记录。
在实验后期, 模拟鼠类埋藏实验, 每次在摆放点母树667(阴坡)和648(阳坡)附近10 m范围内分别随机选取2个埋藏点, 每个埋藏点内放4粒种子, 靠近埋藏点处用白色标签做标记, 标签事先用周围环境的枯落物或树皮擦拭, 掩盖气味, 共进行10轮, 每颗母树附近进行20次埋藏(n=20), 隔天观测埋藏点种子被捕食、消失情况并做记录。
实验同时, 在距摆放点和埋藏点0.8—1 m处布设红外相机(RECON HS200 RT1017R), 观测摆放点昼夜变化情况, 设置每次触发后录制时间5—15 s不等, 次日观察视频, 记录取食动物种类、取食时间等信息。
使用SPSS 19.0处理计算摆放、埋藏实验数据, 并采用单因素方差分析(one-way ANOVA)进行不同摆放组的差异性比较, 显著性水平设定为α=0.05。
1.6 幼苗空间分布格局及鼠巢分布与之相关性大别山五针松天然种群较小, 分布零散, 在研究地地呈狭长带状分布, 特殊的空间分布特征导致其种群研究不适宜采用传统的规则样地法进行调查, 因此采取研究区域内全面调查, 于2015年11—12月对研究地点内(中心地理坐标116°05′33.94″E, 30°49′28.91″N, 研究地面积约4.2 hm2)所有大别山五针松个体测量并记录其胸径尺寸(DBH)等数据, 使用GPS(GARMIN 62 s)获取每株个体坐标。于2016年10—12月再次进行全面调查, 记录该种群数量及个体变化情况, 并对研究地范围内的鼠类洞穴进行搜寻, 使用GPS获取巢穴精确坐标并记录。
参考了红松、华山松等松属植物的径级分类方法[18-20], 结合大别山五针松种群径级特征, 将胸径尺寸DBH≤2 cm的个体作为大别山五针松幼苗。采用Ripley′s K函数[21]对2015年采集到数据进行分析。该函数可描述在不同空间尺度下大别山五针松幼苗的空间分布格局, 反映其种群结构特点。Ripley′s K函数计算公式如下:
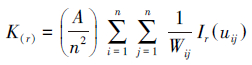
|
式中, A为样地面积, n为总点数(植物个体数), uij为点i和点j之间的距离, 当uij≤j时, Ir(uij)= 1, 当uij≥j时, Ir(uij)= 0;Wij为以点i为圆心, uij为半径的圆落在面积A中的比例。
为了使期望值线性化并保持方差稳定性, Besag提出用L(r)函数来代替K(r)函数[22], 其计算公式为:

|
当L(r)= 0时, 幼苗呈空间完全随机分布;当L(r)<0时, 幼苗呈均匀分布;当L(r)>0时, 幼苗呈集群分布。用Monte-Carlo模拟法拟合检验, 计算上下包迹线(Envelope), 即置信区间。假定空间点是随机分布, 用随机模型拟合一组点的坐标值, 对每一个r值, 计算L(r)值, 即为期望值, 重复进行这一过程直到达到事先确定的次数。99%置信水平的拟合次数为100次。实际分布数据计算得到的不同尺度下的值L(r)值即为观测值。
用r作为横坐标, 上下包迹线作为纵坐标绘图。L(r)的最大值和最小值分别为上下包迹线的坐标值, 若L(r)值在置信区间上方, 则表示呈集群分布格局;若在置信区间内, 则表示呈随机分布格局;若L(r)值在置信区间下方, 则表示呈均匀规则分布格局。
在Ripley′s K函数计算过程中, 根据不同尺度要求, 小尺度范围从0.1 m进行计算, 步长为0.1 m, 计算20次;大尺度范围从2.5 m进行计算, 步长为2.5 m, 计算11次。使用ArcGIS 10.2进行不同尺度的Ripley′s K函数分析, 并绘制观测曲线。
根据2016年调查获得的大别山五针松幼苗坐标记录, 使用ArcGIS 10.2绘制大别山五针松幼苗分布热力图, 并将调查获取研究地范围鼠类巢穴分布情况与之对比、分析。
2 研究结果 2.1 球果与种子特征四株母树球果中种子的空瘪率极高(表 1), 母树660所有球果中甚至未发现一粒含种仁的松子, 空瘪率达到100%。阴坡母树球果霉变率较高, 球果的长度、直径和外观发育完整程度均低于阳坡母树的球果, 且阴坡球果空瘪率更高。
| 母树编号 Seed tree nuber |
位置 Location |
霉变率 Decay proportion/% |
球果长 Cone length/cm |
球果直径 Cone diameter/cm |
出种量 Seed output |
空瘪率 Empty/% |
| 645(n=10) | 阴坡 | 90.0 | 10.11±1.33 | 5.08±0.13 | 31.70±3.49 | 96.3±2.5 |
| 646(n=18) | 阴坡 | 100.0 | 10.09±0.23 | 5.21±0.11 | 30.55±4.75 | 82.2±4.8 |
| 660(n=5) | 阴坡 | 93.5 | 9.14±0.58 | 4.94±0.24 | 35.60±10.61 | 100.0±0 |
| 648(n=2) | 阳坡 | 0 | 12.02±0.16 | 6.24±0.08 | 34.00±2.35 | 79.4±10 |
2016年10—11月中旬, 种子雨调查期间, 收集网共收集到掉落的球果4枚, 未发现掉落的完整种子。
在4株母树下60个小样方3轮土壤种子库调查中, 未在任何样方发现完整种子, 仅在17个样方内发现残破的种壳, 种壳位置均为地面枯落物下, 土壤内未发现残破种壳。
2.3 访问动物组成调查共记录到访问大别山五针松母树或球果的动物16种(6目11科)(表 2)。能够确定取食球果或松子的动物7种。黑尾蜡嘴雀(Eophona migratoria)停歇于大别上五针松树枝上, 偶尔记录到单只取食大别山五针松种子, 未见搬运行为;野猪主要在林下地面搜寻掉落的松果, 经常可见到被啃食过半的球果;调查期间多次于大别山五针松林下地面发现勺鸡(Pucrasia macrolopha)和环颈雉(Phasianus colchicus)活动或听到鸣叫, 已有二者取食大别山五针松种子的相关报道[14];同时, 走访调查得知狗獾(Meles meles)会在林下取食种子, 但未见到实体, 以上5种动物均为种子捕食者。
| 种类Species | 访问位置Visiting location | 访问行为Visiting behaviors |
| 野猪Sus scrofa | 林下地面 | 啃食球果 |
| 狗獾Meles meles* | 林下地面 | 啃食球果 |
| 岩松鼠Sciurotamias davidianus | 林下地面、树枝 | 啃食球果、取食松子、种子搬运 |
| 小林姬鼠Apodemus sylvaticus | 林下地面 | 啃食球果、取食松子、种子搬运 |
| 勺鸡Pucrasia macrolopha | 林下地面 | 取食松子 |
| 环颈雉Phasianus colchicus | 林下地面 | 取食松子 |
| 大斑啄木鸟Dendrocopos major | 树枝 | 停歇 |
| 红耳鹎Pycnonotus jocosus | 树枝 | 停歇、查看松子 |
| 栗背短脚鹎Hemixos castanonotus | 树枝 | 停歇、查看松子 |
| 松鸦Garrulus glandarius | 树枝 | 停歇、查看松子 |
| 红嘴蓝鹊Urocissa erythrorhyncha | 树枝 | 停歇、查看松子 |
| 红胁蓝尾鸲Tarsiger cyanurus | 树枝 | 停歇、查看松子 |
| 棕颈钩嘴鹛Pomatorhinus ruficollis | 树枝 | 停歇、查看松子 |
| 黑尾蜡嘴雀Eophona migratoria | 树枝 | 停歇、取食松子 |
| 大山雀Parus major | 树枝 | 停歇 |
| 红头长尾山雀Aegithalos concinnus | 树枝 | 停歇 |
| *走访调查结果 | ||
另外两种取食球果或种子的动物均为啮齿动物, 岩松鼠(Sciurotamias davidianus)为调查地区唯一见到的日行性啮齿动物, 而小林姬鼠(Apodemus sylvaticus)则是唯一一种拍摄、记录并被捕捉到的夜行性鼠类, 二者均有取食、搬运散落地面上种子以及处理大别山五针松球果的行为, 为大别山五针松种子的潜在传播者。
需要提出的是, 大别山五针松群落范围内松鸦(Garrulus glandarius)的数量较为可观, 但2015—2016两年观察期未见松鸦对大别山五针松种子有取食、搬运等行为。
2.4 地面种子命运隔天回访摆放点, 回收红外相机, 发现经过啮齿动物的捕食和搬运, 不同摆放粒数的大别山五针松种子总留存率、总捕食率和总搬运率分别为(1.25±3.00)%、(6.04±5.40)%和(92.71±5.56)%, 所有摆放组留存率均不超过5%, 搬运率均大于89%, 各组95%以上的种子被捕食或搬运, 大别山五针松种子在不同的摆放粒数情况下, 啮齿动物的留存率(F7, 24=1.071, P=0.411>0.05)、捕食率(F7, 24=0.468, P=0.848>0.05)和搬运率(F7, 24=0.569, P=0.774>0.05)均无显著性差异(表 3)。
| 摆放粒数 Seed-placed number |
留存率 Remaining (Mean±SE)/% |
捕食率 Predation (Mean±SE)/% |
搬运率 Removal (Mean±SE)/% |
| 10(n=4) | 0 | 5.00±5.77 | 95.00±5.77 |
| 20(n=4) | 2.50±5.00 | 2.50±5.00 | 95.00±5.77 |
| 30(n=4) | 0 | 6.68±4.71 | 93.33±4.71 |
| 40(n=4) | 1.25±2.50 | 8.13±2.39 | 90.63±4.27 |
| 50(n=4) | 0.50±1.00 | 4.50±3.00 | 95.00±3.46 |
| 100(n=4) | 0.75±0.96 | 8.25±10.14 | 91.00±10.13 |
| 150(n=4) | 4.50±6.12 | 6.02±3.94 | 89.48±2.38 |
| 200(n=4) | 5.00±0.58 | 7.25±7.14 | 92.25±6.80 |
| 总计Total(n=32) | 1.25±3.00 | 6.04±5.40 | 92.71±5.56 |
红外相机监测显示, 岩松鼠与小林姬鼠是研究地大别山五针松种子的主要搬运者。岩松鼠的取食活动最早记录为05:43, 活动持续至11:00时, 其觅食高峰期为15:20—18:00, 其更偏好于大片、裸露、光洁的岩层上取食, 极少出现于林下枯落物处觅食。以母树666裸岩区摆放点为例, 当岩松鼠发现裸露岩石上的种子后, 会先取食部分种子。岩松鼠会牙齿打磨松子种壳进行试探, 判定其饱满程度, 取食饱满的松子, 丢弃干瘪种子, 进食少部分种子后, 在当天日落前会分批次将剩余种子全部搬运。小林姬鼠的主要活动时间为19:00—24:00, 对于枯落物摆放区的种子消耗很大, 其对种子处理方式与岩松鼠类似。小林姬鼠在进食过后也会对种子进行反复搬运, 直到摆放点种子全部被搬运走为止。
在同域啮齿类动物通常会有相互盗食行为, 盗食行为是贮藏食物损失的重要原因。在隔天检查时发现, 贮点被盗挖现象严重, 由表 4可以看出, 各贮点当夜被发现的概率均达到90%以上, 两株母树附近, 种子的总留存率和总搬运率分别为(11.25±23.99)%和(88.75±23.99)%, 在所有的埋藏实验点, 均没有发现贮藏种子被取食。另外, 红外相机监测显示, 埋藏点的盗挖现象均是小林姬鼠造成的。
| 母树编号 Seed tree nuber |
留存率 Remaining(Mean±SE)/% |
捕食率 Predation (Mean±SE)/% |
搬运率 Removal (Mean±SE)/% |
发现率 Finding% |
| 667(n=20) | 15.00±30.78 | 0.00 | 85.00±30.78 | 90 |
| 648(n=20) | 7.50±14.28 | 0.00 | 92.50±14.28 | 100 |
| 总计Total(n=40) | 11.25±23.99 | 0.00 | 88.75±23.99 |
从2015年研究地大别山五针松母树及幼苗基株分布来看(图 1), 大别山五针松母株共计22株(母树644因距离种群集中区较远, 不便作图, 排除), 幼苗42株。根据幼苗轮枝数判别, 发现研究地的大别山五针松种群分别于2015年和2016年新增一年生幼苗5株和7株。研究发现, 幼苗更偏好在背阴的东北坡着生, 调查期间常发现数株幼苗聚集在一定范围内, 但彼此分开不成簇。

|
| 图 1 大别山五针松母树与幼苗分布情况 Fig. 1 The distribution of P. dabeshanensis |
在大、小尺度上大别山五针松幼苗的点格局分析结果显示(图 2), 从小尺度上看, 大别山五针松幼苗在0.1—0.6 m范围呈随机分布, 在0.6—2.0 m范围呈聚集分布(图 2A);从大尺度上看, 大别山五针松幼苗在2.5—27.5 m范围均呈聚集分布(图 2B)。综上所述, 大别山五针松幼苗在0—0.6 m尺度范围内呈随机分布;在>0.6 m尺度范围上呈聚集分布。

|
| 图 2 大别山五针松两个尺度上的幼苗点格局 Fig. 2 Point pattern analysis for the seedings of P. dabeshanensis in the two scales |
通过红外相机监测以及动物调查, 确定在研究地搜寻到的鼠穴均为小林姬鼠巢穴, 从幼苗的热力图分布及小林姬鼠巢穴分布情况来看(图 3), 巢区分布与幼苗分布存在一定的相关性, 巢区附近往往会存在幼苗群。

|
| 图 3 大别山五针松幼苗分布热力图 Fig. 3 The thermodynamic diagram of the seedings of P. dabeshanensis |
大别山五针松球果发育质量与个体分布有关。大别山五针松种群零散, 个体间距离通常较远, 伴生植物群落密集, 花粉难以飞散, 个体之间不能进行有效的基因交流[10], 自交率极高, 空瘪粒相对较多;散生母树的种子空瘪粒则特别多[23], 本研究结果也证实了其种子空瘪率高。
有研究表明, 气候因素对大别山五针松球果的发育也有较大影响[10, 24]。本研究调查发现, 大别山五针松成体多生于阴坡或半阴坡, 湿润的气候和阴坡较少的光照使得五针松球果发育极差, 而阳坡开阔地的大别山五针松球果发育良好程度要远远高于阴坡冠层下方球果;同时, 由于大别山区雨量充沛、温暖潮湿, 大别山五针松球果受雨、受潮后极易发霉, 霉斑与松脂在种鳞外部形成白色蜡斑, 使种鳞相互粘连, 阻碍种鳞张开;此外, 阴坡球果极少受到太阳光照, 种鳞内经常被一些蜘蛛当做巢穴, 即使球果发育良好, 种子也受繁密的蛛网限制不能脱出, 对球果和种子的发育起到限制作用。
本研究中未发现种子雨, 大别山五针松球果成熟后, 大部分种子不易自行掉落, 这与红松、偃松(Pinus pumila)等部分五针松组(Sect. Strobus)植物特性相似[25], 球果整体掉落后经动物取食、埋藏可能是其进入土壤种子库的主要途径。土壤种子库调查发现, 各样方未能发现完整种子, 只有个别残破种壳留存于枯落物中, 推测一方面可能是因为自行脱落的大别山五针松种子稀少, 所选样方内均未包含, 另一方面可能由于林下高强度的动物取食造成鲜有种子能够顺利进入土壤种子库, 这是其天然更新中缺乏幼苗的主要原因。
3.2 动物对大别山五针松种子的取食与潜在传播作用通过两年的动物取食和搬运观测, 我们发现两个显著特点:(1)动物对种子的捕食率很高。岩松鼠对树上、林下掉落的球果内种子进行广泛的搜集;勺鸡、环颈雉、野猪等动物在白天对风吹落的球果及种子发现能力强;小林姬鼠通过敏锐的嗅觉夜间在林下搜索。摆放试验中, 不同的摆放粒数对啮齿动物捕食、搬运大别山种子没有显著影响, 这表明动物会尽可能多的搜集可获取的食物, 对大别山五针松种子具有极高的捕食效率。有研究报道岩松鼠等小型啮齿动物在大别山五针松球果刚成熟时便会啃食, 对大别山五针松的种群起到一定的抑制作用[14]。(2)作为种子取食者, 动物在不断演化中形成了适应性对策, 许多啮齿类都表现出贮食行为[26-28], 从地面种子摆放实验结果来看, 动物对大别山五针松种子搬运和贮藏比例极高, 约有90%左右的种子会被岩松鼠及小林姬鼠搬运、贮藏, 动物的贮藏行为对大别山五针松的更新应该有着重要的正面影响。虽然很多啮齿动物会将种子集中贮藏在巢穴中, 由于种子埋藏较深, 可能不利于种子萌发[29-30], 但由于野猪拱土觅食[31]以及其他鼠类对贮藏点盗挖造成的二次传播等行为, 对大别山五针松种群更新也可能起到一定的积极作用。
此外, 国内外许多研究显示, 鸦科鸟类是重要的松属植物种子传播者[7, 32], 能够在树上直接从球果里拔出种子, 利用嗉囊携带搬运种子, 并且由于飞行距离较远, 能够大范围地扩散传播种子[33]。本研究中, 虽然大别山五针松群落范围内松鸦的数量较为可观, 但在2015—2016两年研究期间却未能观察到松鸦对大别山五针松种子有取食、贮藏行为, 这也是未来研究关注的重点。
3.3 啮齿动物对大别山五针松种群更新的作用2015年和2016年期间, 研究地大别山五针松种群分别新增一年生幼苗5株和7株, 这表明大别山五针松在当地存在天然更新。通过对大别山五针松幼苗空间分布格局分析, 我们发现, 与大多数果松不同, 研究地大别山五针松的幼苗多为单生, 调查时偶见两株幼苗萌发于一处。幼苗多单独生长在母树周围, 在0—0.6 m的尺度范围内呈随机分布;在>0.6 m尺度呈聚集分布。这种一定尺度范围的聚集分布与鼠类贮食传播的松树类似[34-35], 但单生苗数量众多又与其他动物传播松有着明显的不同, 对此我们认为可能是以下情况造成的:
(1) 最直接的因素是大别山五针松种子产量较低, 落至地面的饱满种子极易被林下动物取食, 只有极少数进入土壤种子库, 并在适宜萌发的环境中建成幼苗;
(2) 大别山五针松种子作为稀缺的高质量食物, 啮齿动物搬运、贮藏后, 对贮点的重取率较高, 偶有未被重取的种子得以萌发建成幼苗, 而贮食动物为避免集中贮藏被盗食动物一次性盗食, 采用分散贮藏方式贮食[36-37], 但大别山五针松种子产量较低, 使得每个贮点内放置的种子量极少, 因此每个贮点能成功萌发的幼苗仅1—2株;
(3) 由于大别山五针松着生地海拔较高, 多为阴坡, 年均气温低, 光、水分等幼苗生存必须的资源较匮乏, 贮点内同时萌发的幼苗对资源的竞争十分激烈, 同一个贮点内萌发的幼苗间强烈的自疏作用造成了这种现象[38]。
大别山五针松种群幼苗主要的聚集区域为大别山五针松母树下坡位附近范围, 东北坡为主, 西南方向幼苗数量极少, 幼苗的分布格局与小林姬鼠巢穴位置关系密切。小林姬鼠同样面临着种内盗食的威胁, 因此对于小林姬鼠来说, 将觅食发现和盗取获得的食物埋藏于巢穴附近或直接搬运至巢穴可能是其最好的选择。通过大别山五针松幼苗分布热力图与研究地鼠类巢穴分布情况对比发现, 小林姬鼠巢穴周边的分布有大量幼苗, 且其多位于北坡的特点又与大别山五针松幼苗乃至种群的着生特点高度重合。前人研究发现, 为提高觅食效率, 鼠类喜爱在巢穴周围设置贮点[39], 且贮点多设在洞穴与种源的连线位置[40], 贮点内的种子如果未被取食, 在适宜的条件下便会萌发建成幼苗, 这一特征与热力图显示的的幼苗在小范围离散分布、大范围聚集分布这一结果相符。因此, 小林姬鼠极有很可能为大别山五针松的传播者, 对大别山五针松种群更新有着积极作用。
而研究期间未发现大别山五针松有密集的种子雨, 曾数次见到多个泛青色未成熟球果和成熟球果被堆置在母树上坡位, 观察可知, 由于林下映山红等灌木遮挡, 重力或风力较难将球果搬运带至这些地方, 这极有可能是岩松鼠取食、搬运造成, 岩松鼠对球果的破坏以及贮食行为使得种子落到地面或进入土壤。同期进行埋藏试验并使用红外相机监测显示, 小林姬鼠存在严重的盗食现象, 由此推测, 小林姬鼠通过盗挖造成对大别山五针松种子的二次传播可能是大别山五针松种群天然更新的一个重要手段。
| [1] | Richardson D M, Rundel P W. Ecology and biogeography of Pinus: an introduction//Richardson D M, ed. Ecology and Biogeography of Pinus. Cambridge, UK: Cambridge University Press, 1998: 3-48. |
| [2] | 鲁长虎. 动物对松属植物种子的传播作用研究进展. 生态学杂志, 2006, 25(5): 557–562. DOI:10.3321/j.issn:1000-4890.2006.05.018 |
| [3] | Vander Wall S B. Food Hoarding in Animals. Chicago: The University of Chicago Press, 1990: 1–445. |
| [4] | Tomback D F, Kramer K A. Limber pine seed harvest by Clark's nutcracker in the sierra Nevada:Timing and foraging behavior. The Condor, 1980, 82(4): 467–468. DOI:10.2307/1367579 |
| [5] | Johnson M, Vander Wall S B, Borchert M. A comparative analysis of seed and cone characteristics and seed-dispersal strategies of three pines in the subsection Sabinianae. Plant Ecology, 2003, 165(1): 69–84. DOI:10.1023/A:1021598712920 |
| [6] | 鲁长虎. 动物与红松天然更新关系的研究综述. 生态学杂志, 2003, 22(1): 49–53. |
| [7] | 粟海军, 马建章, 宗诚. 四种昼行性动物取食和贮藏红松种子的行为比较. 动物学杂志, 2007, 42(2): 10–16. DOI:10.3969/j.issn.0250-3263.2007.02.003 |
| [8] | Chen F, Chen J. Dispersal syndrome differentiation of Pinus armandii in Southwest China:Key elements of a potential selection mosaic. Acta Oecologica, 2011, 37(6): 587–593. DOI:10.1016/j.actao.2011.05.009 |
| [9] | 常罡, 王开锋, 王智. 秦岭森林鼠类对华山松种子捕食及其扩散的影响. 生态学报, 2012, 32(10): 3177–3181. |
| [10] | 项小燕, 张小平, 段仁燕, 徐延年, 吴甘霖. 濒危植物大别山五针松母树林花粉传播规律. 广西植物, 2014(3): 333–337. |
| [11] | 徐观名. 岳西大王沟大别山五针松种群保护面临的问题与对策. 安徽林业科技, 2016, 42(S1): 69–72. |
| [12] | 韩建伟, 张智勇, 王恩茂, 李颖岳. 大别山五针松种子特性及促进种子萌发的研究. 中国农学通报, 2014, 30(1): 5–10. |
| [13] | 侯成林, 操丛林, 汪文革, 尹湘豫, 高健, 韩久同. 大别山五针松病害调查(简报). 安徽农业大学学报, 1995, 22(1): 53–56. |
| [14] | 彭镇华, 江泽慧. 大别山五针松及起源. 北京: 中国林业出版社, 1999: 1–86. |
| [15] | 祝霞灿. 大别山五针松生长发育情况调查. 安徽林业科技, 2005(4): 16–17. |
| [16] | 徐燕英.大别山五针松群落生态学初步研究[D].淮北: 淮北师范大学, 2012. |
| [17] | 项小燕, 吴甘霖, 段仁燕, 闫玉梅, 张小平. 大别山五针松种内和种间竞争强度. 生态学报, 2015, 35(2): 389–395. |
| [18] | 苏志尧, 吴大荣, 陈北光. 粤北天然林优势种群结构与空间格局动态. 应用生态学报, 2000, 11(3): 337–341. DOI:10.3321/j.issn:1001-9332.2000.03.004 |
| [19] | 胡元洁, 巨天珍, 米彩燕, 康晓亚, 张宋智, 马建伟. 小陇山国家级自然保护区华山松种群更新动态分析. 广西植物, 2013, 33(2): 247–252. DOI:10.3969/j.issn.1000-3142.2013.02.021 |
| [20] | 陈列.长白山阔叶红松林主要树种种群结构及其林木径向生长对气候响应[D].北京: 北京林业大学, 2014. http://cdmd.cnki.com.cn/article/cdmd-10022-1015515146.htm |
| [21] | Ripley B D. Modelling spatial patterns (with discussion). Journal of the Royal Statistical Society:Series B (Methodological), 1977, 39: 172–212. |
| [22] | Besag J E. Comments on Ripley's paper. Journal of Rural Statistic:Series B, 1977, 39: 193–195. |
| [23] | 汪为民. 大别山五针松生物学特性观察研究. 安徽林业科技, 2007(2): 20–20. |
| [24] | 吴甘霖, 羊礼敏, 段仁燕, 王志高, 张中信. 大别山五针松林林窗、林缘和林下的微气候特征. 生物学杂志, 2017, 34(4): 64–66. |
| [25] | 中国科学院中国植物志编辑委员会. 中国植物志:第七卷裸子植物门. 北京: 科学出版社, 1978: 211–221. |
| [26] | Howe H F, Smallwood J. Ecology of seed dispersal. Annual Review of Ecology and Systematics, 1982, 13(1): 201–228. DOI:10.1146/annurev.es.13.110182.001221 |
| [27] | Steele M A, Hadjchikh L Z, Hazeltine J. Caching and feeding decisions by Sciurus carolinensis:responses to weevil-infested acorns. Journal of Mammalogy, 1996, 77(2): 305–314. DOI:10.2307/1382802 |
| [28] | Hopewell L J, Leaver L A, Lea S E G. Effects of competition and food availability on travel time in scatter-hoarding gray squirrels (Sciurus carolinensis). Behavioral Ecology, 2008, 19(6): 1143–1149. DOI:10.1093/beheco/arn095 |
| [29] | 肖治术, 张知彬. 啮齿动物的贮藏行为与植物种子的扩散. 兽类学报, 2004, 24(1): 61–70. DOI:10.3969/j.issn.1000-1050.2004.01.011 |
| [30] | Vander Wall S B, Enders M S, Waitman B A. Asymmetrical cache pilfering between yellow pine chipmunks and golden-mantled ground squirrels. Animal Behaviour, 2009, 78(2): 555–561. DOI:10.1016/j.anbehav.2009.05.030 |
| [31] | 刘鹤, 李乐, 马强, 万冬梅, 张树清, 祝业平. 野猪研究进展. 四川动物, 2011, 30(2): 310–314. DOI:10.3969/j.issn.1000-7083.2011.02.018 |
| [32] | Hutchins H E, Lanner R M. The central role of Clark's nutcracker in the dispersal and establishment of whitebark pine. Oecologia, 1982, 55(2): 192–201. DOI:10.1007/BF00384487 |
| [33] | 王巍, 马克平. 岩松鼠和松鸦对辽东栎坚果的捕食和传播. 植物学报(英文版), 1999, 41(10): 1142–1144. DOI:10.3321/j.issn:1672-9072.1999.10.022 |
| [34] | 聂志文, 布仁仓, 李冰伦. 红松存在/不存在数据的多尺度空间分布格局. 生态学报, 2017, 37(3): 810–818. |
| [35] | 刘洋洋, 巨天珍, 王振乾, 高凤如, 赵欣鑫, 王菁. 宝天高速路域华山松种群空间分布格局研究. 湖北农业科学, 2017, 56(4): 691–696. |
| [36] | 路纪琪, 肖治术, 程瑾瑞, 张知彬. 啮齿动物的分散贮食行为. 兽类学报, 2004, 24(3): 267–272. DOI:10.3969/j.issn.1000-1050.2004.03.014 |
| [37] | Grodzinski U, Watanabe A, Clayton N S. Peep to pilfer:what scrub-jays like to watch when observing others. Animal Behaviour, 2012, 83(5): 1253–1260. DOI:10.1016/j.anbehav.2012.02.018 |
| [38] | 许远, 吴甘霖, 羊礼敏, 段仁燕, 项小燕, 王志高. 斑块生境下大别山五针松幼苗的光合生理特征. 生物学杂志, 2017, 34(5): 70–73. DOI:10.3969/j.issn.2095-1736.2017.05.070 |
| [39] | 李宏俊, 张知彬. 动物与植物种子更新的关系Ⅱ.动物对种子的捕食、扩散、贮藏及与幼苗建成的关系. 生物多样性, 2001, 9(1): 25–37. DOI:10.3321/j.issn:1005-0094.2001.01.004 |
| [40] | Vander Wall S B. Dispersal of singleleaf piñon pine (Pinus monophylla) by seed-caching rodents. Journal of Mammalogy, 1997, 78(1): 181–191. DOI:10.2307/1382651 |