 2018, Vol. 38
2018, Vol. 38文章信息
- 叶锦玉, 张衡, 张瑛瑛, 靳少非.
- YE Jinyu, ZHANG Heng, ZHANG Yingying, JIN Shaofei.
- 长江口盐沼植被生境内斑尾刺虾虎鱼食性特征
- Feeding habits of Acanthogobius ommaturus in the salt marsh in the Yangtze Estuary
- 生态学报. 2018, 38(17): 6217-6227
- Acta Ecologica Sinica. 2018, 38(17): 6217-6227
- http://dx.doi.org/10.5846/stxb201707271356
-
文章历史
- 收稿日期: 2017-07-27
- 网络出版日期: 2018-05-30





2. 上海海洋大学海洋学院, 上海 201306;
3. 闽江学院海洋学院, 福州 350108;
4. 闽江学院海洋研究院, 福州 350108
2. College of Marine Sciences, Shanghai Ocean University, Shanghai 201306, China;
3. Department of Geography, Ocean College, Minjiang University, Fuzhou 350108, China;
4. Institute of Oceanography, Minjiang University, Fuzhou 350108, China
河口盐沼是河水径流汇入海洋的通道, 既有淡水也有咸水生境。很多河口区生长有盐沼或红树林植被, 使得河口区生物的分布格局和栖息地选择具有一定的独特性, 特别是对于潮间带生物(如鱼类、底栖动物和植物)来说[1-3]。长江口作为中国最大的河口, 也是中国渔业生产力最高的生态系统之一, 潮滩湿地类型丰富多样, 大面积的湿地生境给予了河口鱼类(特别是幼鱼)有益的育幼场功能, 支持了河口及邻近海域众多鱼类的栖息和发育等生态功能[3]。虾虎鱼类是河口鱼类的优势类群, 主要栖息于河口浅水区包括盐沼植被分布区。其中, 斑尾刺虾虎鱼(Acanthogobius ommaturus)作为长江口湿地鱼类群落的优势种之一, 经常随着潮水进入盐沼植被区栖息和摄食[4-6], 其生活史的幼体阶段会季节性利用盐沼生境, 但利用方式和原理还不甚清楚。
斑尾刺虾虎鱼隶属鲈形目(Pereiformes)虾虎鱼科(Gobiidae), 曾命名斑尾复虾虎鱼(Synechogobius ommaturus)或矛尾复虾虎鱼(Synechogobius hasta), 为暖温性近岸底层鱼类, 喜栖息于泥底浅海区, 亦可进入沿海淡水水域;中国东海、南海、黄海和渤海均有分布[2]。斑尾刺虾虎鱼具有较高的经济价值, 也是国家一级保护动物中华鲟(Acipenser sinensis)等大型鱼类的主要饵料生物, 在长江口生态系统中具有重要的作用[3]。已有少量研究表明斑尾刺虾虎鱼是一种凶猛性肉食性鱼类, 喜食各种幼鱼、虾、蟹和小型软体动物[2], 但未涉及具体的饵料组成;韩东燕等[7]对胶州湾斑尾刺虾虎鱼的摄食生态做了初步研究, 范海洋等[8]研究了黄河三角洲斑尾刺虾虎鱼的渔业生物学特征。由于鱼类食性组成具有明显的地理差异, 针对长江口斑尾刺虾虎鱼摄食习性相关的研究极少见报道[9];而涉及盐沼植被生境内的斑尾刺虾虎鱼生物学特征和食性组成尚未见报道。因此, 本研究根据盐沼植被生境内斑尾刺虾虎鱼的调查数据, 分析其生物学特征以及食性组成, 以期掌握其利用盐沼生境的内在机制和原因。
1 材料与方法 1.1 采样点布设和采集方法于2015年5月至2016年4月(其中1和2月因属于植被凋落期未作调查)在长江口东滩湿地东北水域(即东旺沙站点, 经纬度为31°35′12″N, 121°54′28″E)的盐沼植被区(海三棱藨草(Scirpus mariqueter)和岛屿型互花米草(Spartina alterniflora))进行了斑尾刺虾虎鱼的样本采集, 分析盐沼植被生境斑尾刺虾虎鱼的生物学和食性组成特征。在海三棱藨草和互花米草两种生境内各设置1顶网具, 网具类型为插网[1, 6], 网墙的方向与江堤方向平行, 网高约80 cm, 网长约20 m, 网身网目尺寸8 mm, 每顶网两端装有袖网5 m, 分别伸向大堤方向, 网具的两端各装有1网袋, 以利于提高捕捞效率, 囊网尺寸为6 mm。每月在大潮汛期进行白天和夜晚潮水的两次渔获取样, 每潮水拦截时间约12 h;考虑到进入盐沼植被的班尾刺虾虎鱼数量相对较少, 本研究将两顶网具的白天和晚上的渔获样品合为一个样本作为该月的渔获样本。退潮后在网袋处取鱼, 全部带回实验室进行分析, 同时记录温度(精确到0.1℃)和盐度。种类全部鉴定到种[2-3], 计数, 用吸水纸吸去体表多余的水后称量其湿质量(精确到0.1 g)。根据白天潮水和夜晚潮水的样品, 分析斑尾刺虾虎鱼胃含物组成的昼夜差异。
调查期间, 水温和盐度均具有明显的季节变化;水温变化范围为8.8—31.0℃, 盐度变化范围为3.4—19.5。
1.2 数据分析记录每尾鱼的标准体长(吻端至尾椎骨末端;The standard length, SL)作为该尾鱼的体长(精确到0.1 mm)。鱼类发育阶段划分为幼鱼和成鱼(本研究以体长大于160 mm定义为成鱼)[2-3]。分析斑尾刺虾虎鱼的摄食等级以及胃含物种类、数量和质量组成特征等;摄食等级划分为0—4级[10]。胃含物种类参照文献[2-3, 11-12]进行种类鉴定, 对于消化程度较低的胃含物直接通过形态特征鉴定其种类, 对于消化程度较高的胃含物通过额刺、残肢等加以鉴定, 并尽量鉴定到最小分类单元。
采用质量百分比W(%)、个体数百分比N(%)、出现频率F(%)和相对重要性指数百分比IRI(%)等指标来分析斑尾刺虾虎鱼各饵料组成的重要程度[13-14]。采用饱满指数P(%)、摄食等级来描述其摄食强度[14-15]。

|
(1) |

|
(2) |

|
(3) |
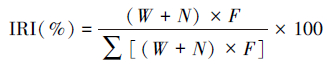
|
(4) |

|
(5) |
采用营养生态位宽度指数进行斑尾刺虾虎鱼的生态位宽度差异分析, 计算公式如下[16-17]:
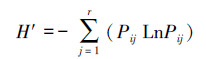
|
(6) |
式中, Pij为饵料生物i 在不同体长组j的食物组成中所占的质量百分比。
采用PRIMER 5.0软件对斑尾刺虾虎鱼不同体长组的饵料质量百分比数据进行平方根转换后进行体长组间Bray-Curtis相似性系数计算, 采用等级聚类(即组平均法)的分类方法对其个体摄食差异进行聚类分析[18]。采用SPSS 17.0统计软件分析不同采样时间(白天与夜晚)下斑尾刺虾虎鱼各饵料类别的质量百分比差异(One-way ANOVA)。
2 结果 2.1 丰度分布调查期间, 共采集到斑尾刺虾虎鱼186尾, 其中6—8月采集的数量较高, 而其他月份较低, 其中12月和4月未采集到样品(表 1)。从不同体长组的月变化来看, 5月均为41—60 mm的个体, 6—8月大个体的鱼类比例逐渐增加, 在7月和8月发现少量成体(尾数比例分别为4%和8%)。9月因风浪较大, 导致渔获尾数下降明显, 10—11月大个体的鱼类比例相对较高, 11月采集到斑尾刺虾虎鱼最大体长组(181—200 mm)个体1尾。5—11月斑尾刺虾虎鱼个体平均体质量逐渐增大, 但9月因样本数仅2尾导致平均体质量有所降低。幼鱼比例在3月、5—6月、9—10月均为100%, 而7—8月幼鱼比例仍高达90%以上, 11月幼鱼比例相对较低, 但仍高达80%。
| 时间 Time |
体长组Body length groups/mm | 尾数 Number of individuals |
平均体质量 Biomass (Mean value ±SE)/(g/尾) |
幼鱼比例 Percentage of young fishes/% |
|||||||
| 41—60 | 61—80 | 81—100 | 101—120 | 121—140 | 141—160 | 161—180 | 181—200 | ||||
| 2015-05 | 3(100) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 0(0) | 3 | 1.4±0.2 | 100 |
| 2015-06 | 7(17) | 14(34) | 9(22) | 9(22) | 1(3) | 0(0) | 0(0) | 0(0) | 40 | 9.4±1.1 | 100 |
| 2015-07 | 0(0) | 7(10) | 12(17) | 24(34) | 13(19) | 11(16) | 3(4) | 0(0) | 70 | 21.4±1.7 | 96 |
| 2015-08 | 0(0) | 4(8) | 5(9) | 14(25) | 26(46) | 2(4) | 4(8) | 0(0) | 56 | 23.7±1.8 | 93 |
| 2015-09 | 0(0) | 1(50) | 0(0) | 1(50) | 0(0) | 0(0) | 0(0) | 0(0) | 2 | 12.5±0.3 | 100 |
| 2015-10 | 0(0) | 0(0) | 0(0) | 1(33) | 1(33) | 1(33) | 0(0) | 0(0) | 3 | 25.2±0.6 | 100 |
| 2015-11 | 0(0) | 0(0) | 3(30) | 4(40) | 1(10) | 0(0) | 1(10) | 1(10) | 10 | 28.0±0.7 | 80 |
| 2015-12 | |||||||||||
| 2016-03 | 1(50) | 0(0) | 0(0) | 0(0) | 1(50) | 0(0) | 0(0) | 0(0) | 2 | 13.7±0.1 | 100 |
| 2016-04 | |||||||||||
| 括号内数据为不同体长组的尾数比例% | |||||||||||
斑尾刺虾虎鱼主要摄食虾类(IRI(%)=69.05)、鱼类(IRI(%)=17.31)、蟹类(IRI(%)=11.51)和双壳类(IRI(%)=0.95)等11大类41小类饵料生物(表 2)。虾类作为最重要的饵料类别, 其重量、数量和出现频次百分比均排列第一, 以安氏白虾(Exopalaemon annandalei)、日本沼虾(Macrobrachium nipponense)、长额刺糠虾(Acanthomysis longirostris)等虾类为主要饵料。鱼类饵料中, 以大鳍弹涂鱼(Periophthalmus magnuspinnatus)、大弹涂鱼(Boleophthalmus valenciennes)等虾虎鱼科鱼类为主。蟹类饵料中, 以长足长方蟹(Metaplax longipes)、隐秘螳臂相手蟹(Chiromantes neglectum)等为主, 其余为不可辨认蟹类饵料。双壳类饵料中, 以彩虹明樱蛤(Iridona iridescens)为主。腹足类、端足类(主要是蜾蠃蜚科sp.和钩虾类未定种)、多毛类等饵料的重要值相对较小。
| 食物组成Food Species | W/% | N/% | F/% | IRI/% |
| 鱼类Fishes | 37.59 | 8.79 | 21.97 | 17.31 |
| 大弹涂鱼Boleophthalmus valenciennes | 13.64 | 1.40 | 2.62 | 4.22 |
| 大鳍弹涂鱼Periophthalmus magnuspinnatus | 11.08 | 1.40 | 3.66 | 4.91 |
| 阿部鲻虾虎鱼Mugilogobius smitt | 0.60 | 0.18 | 0.52 | 0.04 |
| 长体刺虾虎鱼Acanthogobius elongata | 0.43 | 0.18 | 0.52 | 0.03 |
| 棱鮻Liza carinatus | 1.88 | 0.18 | 0.52 | 0.12 |
| 贝氏Hemiculter bleeker | 1.79 | 0.18 | 0.52 | 0.11 |
| 弹涂鱼属Periophthalmus sp. | 0.94 | 0.18 | 0.52 | 0.06 |
| 虾虎鱼科Gobiidae sp. | 6.22 | 2.63 | 5.76 | 5.46 |
| 银鱼科Salangidae sp. | 0.09 | 0.18 | 0.52 | 0.02 |
| 不可辨鱼种Unidentified Fish | 0.92 | 2.28 | 6.81 | 2.34 |
| 虾类Shrimps | 46.9 | 78.13 | 39.82 | 69.05 |
| 安氏白虾Exopalaemon annandalei | 18.42 | 5.49 | 9.23 | 24.16 |
| 脊尾白虾Exopalaemon carinicauda | 0.37 | 0.11 | 0.18 | 0.49 |
| 白虾属Exopalaemon sp. | 11.61 | 5.26 | 5.29 | 17.97 |
| 日本沼虾Macrobrachium nipponense | 4.29 | 2.45 | 5.24 | 3.78 |
| 中国毛虾Acetes chinensis | 2.47 | 0.18 | 0.52 | 0.15 |
| 长额刺糠虾Acanthomysis longirostris | 1.71 | 43.78 | 0.52 | 2.55 |
| 长臂虾科Palaemonidae sp. | < 0.01 | 0.18 | 0.52 | < 0.01 |
| 糠虾目Mysidacea sp. | 0.02 | 8.23 | 3.66 | 3.24 |
| 糠虾属Acanthomysis sp. | 0.34 | 6.13 | 0.52 | 0.36 |
| 沼虾属Macrobrachium sp. | 0.10 | 0.18 | 0.52 | 0.02 |
| 不可辨虾类Unidentified shrimps | 7.56 | 6.14 | 13.61 | 16.32 |
| 蟹类Crabs | 11.79 | 5.1 | 14.12 | 11.51 |
| 长足长方蟹Metaplax longipes | 4.18 | 0.53 | 1.57 | 0.79 |
| 隐秘螳臂相手蟹Chiromantes neglectum | 1.71 | 0.18 | 0.52 | 0.11 |
| 厚蟹属Helice sp. | 0.13 | 0.35 | 0.52 | 0.03 |
| 螳臂相手蟹属(残体)Chiromantes sp. | 0.04 | 0.18 | 0.52 | 0.01 |
| 螳臂相手蟹属(卵)Egg of Chiromantes sp. | < 0.01 | 0.10 | < 0.01 | 0.01 |
| 沙蟹科Ocypodidae sp. | < 0.01 | 0.18 | 0.52 | 0.01 |
| 不可辨蟹类Unidentified crabs | 5.73 | 3.68 | 10.47 | 10.56 |
| 等足类Isopoda | < 0.01 | 0.35 | 1.05 | < 0.01 |
| 水虱Bosmina | < 0.01 | 0.35 | 1.05 | < 0.01 |
| 端足类Amphipoda | 1.20 | 2.28 | 1.57 | 0.32 |
| 端足类Amphipoda | 1.20 | 2.28 | 1.57 | 0.32 |
| 多毛类Polychaeta | 0.04 | 0.18 | 0.52 | 0.01 |
| 沙蚕目Nereidida sp. | 0.04 | 0.18 | 0.52 | 0.01 |
| 双壳类Bivalves | 1.26 | 2.29 | 5.76 | 0.95 |
| 彩虹明樱蛤Iridona iridescens | 0.44 | 1.23 | 2.62 | 0.47 |
| 河蚬Corbicula fluminea | 0.02 | 0.18 | 0.52 | 0.01 |
| 不可辨双壳类Unidentified bivalvia | 0.80 | 0.88 | 2.62 | 0.47 |
| 腹足类Gastropoda | 0.16 | 1.06 | 3.14 | 0.15 |
| 光滑狭口螺Stenothyra glabra | < 0.01 | 0.35 | 1.05 | 0.04 |
| 优雅河纹螺Fluviocingula elegantula | 0.16 | 0.71 | 2.09 | 0.11 |
| 头足类Cephalopod | 0.68 | 0.18 | 0.52 | 0.05 |
| 不可辨头足类Unidentified cephalopod | 0.68 | 0.18 | 0.52 | 0.05 |
| 昆虫类Insects | < 0.01 | 0.18 | 0.52 | < 0.01 |
| 蝽科Pentatomidae sp. | < 0.01 | 0.18 | 0.52 | < 0.01 |
| 其他Others | 0.37 | 1.77 | 5.22 | 0.59 |
| 泥沙Sand | 0.14 | 0.18 | 0.52 | 0.02 |
| 腐殖质Humus | 0.03 | 0.18 | 0.52 | 0.01 |
| 未知植物Plant | < 0.01 | 0.18 | 0.52 | < 0.01 |
| 消化物Digestion residue | 0.20 | 1.23 | 3.66 | 0.56 |
| W:质量百分比, Weight percentage; N:个体数百分比, Number percentage; F:出现频率, Frequency of occurrence; IRI:相对重要性指数百分比, Index of relativity important | ||||
各月平均饱满指数P(%)波动较大, 3月和5月仅为0.63和0.76, 6—11月则呈逐渐增加趋势(图 1)。从摄食等级来看, 5—11月斑尾刺虾虎鱼平均摄食等级基本呈逐渐增加趋势;5—7月摄食等级为0的个体比例可达10%以上, 8—10月相对较低。摄食等级为4的个体比例从5月的0%逐渐增加到8月的16%, 10月和11月则分别为33%和10%(图 1, 2)。

|
| 图 1 不同月份斑尾刺虾虎鱼的平均胃饱满指数和摄食等级变化(平均值±标准误差) Fig. 1 The monthly changes in the mean stomachs fullness index P (%) and feeding levels of Acanthogobius ommaturus (Mean value ±SE) |

|
| 图 2 不同月份斑尾刺虾虎鱼的摄食等级变化 Fig. 2 The monthly changes in the differential feeding levels of Acanthogobius ommaturus 摄食等级:0级, 空胃;1级, 胃内食物不足胃腔的1/2;2级, 胃内食物占胃腔的1/2;3级, 胃内充满食物且胃壁不膨胀;4级, 胃内充满食物且胃壁膨胀变薄 |
体长组(41—140 mm)斑尾刺虾虎鱼的平均胃饱满指数P (%)随体长的增大而增大, 但在141—180 mm体长组P (%)值有所下降。整体来看, 斑尾刺虾虎鱼的平均胃饱满指数P (%)和摄食等级均随体长的增大而增大(图 3)。从不同体长组来看, 幼鱼(体长 < 160 mm)摄食等级为0的个体比例较高(平均值为21%), 而成鱼则为0;而41—100 mm体长组未发现摄食等级为4的个体。可知, 41—100 mm体长组的个体摄食能力较低, 而101—200 mm体长组的摄食能力相对较高(图 4)。

|
| 图 3 不同体长组斑尾刺虾虎鱼的平均胃饱满指数和摄食等级变化(平均值±标准误差) Fig. 3 Changes of the mean stomachs fullness index P(%) and feeding levels for the differential body length groups in Acanthogobius ommaturus (Mean value ±SE) |

|
| 图 4 不同体长组斑尾刺虾虎鱼摄食等级的百分比 Fig. 4 Changes of feeding levels for the differential body length groups in Acanthogobius ommaturus |
41—60 mm体长组的个体主要以虾类作为饵料, 其次为蟹类, 而鱼类的生物量比例极低(< 1%)。而随着体长的增加, 饵料中鱼类的比例呈逐渐增高趋势, 成体的饵料组成中鱼类质量比例达65%以上, 虾类的比例则下降明显(表 3)。
| 类别 Items |
41—60 mm | 61—80 mm | 81—100 mm | 101—120 mm | 121—140 mm | 141—160 mm | 161—180 mm | 181—200 mm | ||||||||
| W/% | F/% | W/% | F/% | W/% | F/% | W/% | F/% | W/% | F/% | W/% | F/% | W/% | F/% | W/% | F/% | |
| 鱼类Fishes | 0.22 | 11.11 | 45.62 | 38.10 | 31.66 | 17.39 | 29.95 | 29.63 | 34.51 | 13.04 | 31.66 | 20.00 | 65.06 | 36.36 | 100 | 100 |
| 虾类Shrimps | 93.09 | 55.56 | 41.00 | 33.33 | 49.30 | 43.48 | 55.12 | 53.70 | 46.14 | 56.52 | 66.02 | 53.33 | 12.39 | 27.27 | 0 | 0 |
| 蟹类Crabs | 6.48 | 22.22 | 0.81 | 4.76 | 14.76 | 26.09 | 9.73 | 9.26 | 18.11 | 23.91 | 0.27 | 6.67 | 21.76 | 18.18 | 0 | 0 |
| 等足类Isopoda | 0 | 0 | 0 | 0 | 0.01 | 4.35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 端足类Amphipoda | 0 | 0 | 0 | 0 | 0 | 0 | 1.44 | 1.85 | 0.49 | 2.17 | 0 | 0 | 0 | 0 | 0 | 0 |
| 多毛类Polychaeta | 0 | 0 | 2.03 | 4.76 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 双壳类Bivalves | 0 | 0 | 8.51 | 12.29 | 4.26 | 8.70 | 1.44 | 3.70 | 0 | 0 | 2.02 | 6.67 | 0.70 | 9.09 | 0 | 0 |
| 腹足类Gastropoda | 0 | 0 | 2.03 | 4.76 | 0 | 0 | 0 | 0 | 0.25 | 2.17 | 0.03 | 13.33 | 0 | 0 | 0 | 0 |
| 头足类Gastropoda | 0 | 0 | 0 | 0 | 0 | 0 | 2.31 | 1.85 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 昆虫类Insects | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.01 | 9.09 | 0 | 0 |
| 其他Others | 0.22 | 11.11 | 0 | 0 | 0 | 0 | 0 | 0 | 0.49 | 2.17 | 0 | 0 | 0 | 0 | 0 | 0 |
研究发现, 白天潮水采集的斑尾刺虾虎鱼丰度要高于晚上, 特别是夏季(6—8月);但也发现其平均体长、平均体质量和摄食等级均要略低于晚上潮水(表 4)。考虑到部分月份的样本数较低, 仅根据6—8月斑尾刺虾虎鱼主要饵料类别分析其昼夜差异;发现白天样本中鱼类饵料的质量百分比显著高于夜晚(P < 0.01), 约是夜晚的8倍(图 5)。但虾类的质量百分比则是夜晚显著高于白天(P < 0.01), 约是白天的2倍, 而蟹类饵料的昼夜差异不明显(P>0.05)。
| 月份 Month |
白天Day | 夜晚Night | ||||||
| 丰度 Abundance/(尾/网) |
ML | MW | MF | 丰度 Abundance/(尾/网) |
ML | MW | MF | |
| 2015.05 | 3 | 4.9±0.2 | 1.6±0.1 | 0.5±0.1 | ||||
| 2015.06 | 23 | 82.6±0.4 | 10.1±0.6 | 1.5±0.2 | 17 | 80.6±0.3 | 8.5±0.4 | 1.5±0.1 |
| 2015.07 | 42 | 109.9±0.4 | 18.8±0.7 | 0.8±0.2 | 28 | 124.1±0.5 | 25.4±0.8 | 2.0±0.2 |
| 2015.08 | 39 | 117.8±0.3 | 22.5±0.8 | 1.5±0.1 | 17 | 125.6±0.4 | 26.4±0.8 | 1.9±0.2 |
| 2015.09 | 1 | 114.0±0.0 | 21.0±0.0 | 3.0±0.0 | 2 | 108.0±0.8 | 17.9±0.2 | 0.8±0.1 |
| 2015.10 | 3 | 125.3±1.1 | 25.2±5.5 | 2.3±0.8 | ||||
| 2015.11 | 7 | 127.5±1.8 | 32.5±8.9 | 2.1±0.4 | 3 | 109.3±0.6 | 17.5±2.2 | 2.3±0.3 |
| 2015.12 | ||||||||
| 2016.03 | 1 | 5.0±0.0 | 2.3±0.0 | 0.3±0.0 | 1 | 130.0±0.0 | 25.1±0.0 | 2.0±0.0 |
| 2016.04 | ||||||||
| 月平均值Mean | 11.3±5.6 | 92.8±0.3 | 17.9±1.5 | 1.5±0.2 | 7.4±3.3 | 100.9±0.3 | 18.5±1.6 | 1.7±0.2 |
| ML:平均体长, Mean body length;MW:平均体质量, Mean weight;MF:平均摄食等级, Mean feeding levels。表中数据为平均值±标准误差 | ||||||||

|
| 图 5 6—8月斑尾刺虾虎鱼不同饵料类别质量百分比的昼夜差异 Fig. 5 The day and night changes in the percentage of weight in the differential food items from June to August in Acanthogobius ommaturus 不同大写字母表示同一采样时间内不同月份间差异显著, 不同小写字母表示同一月份内不同采样时间差异显著(P < 0.05) |
从等级聚类(即组平均法)的分类方法可知, 斑尾刺虾虎鱼各体长组食物组成在10%相似性水平上可分为3类, 即41—60、61—180 mm和181—200 mm(图 6)。

|
| 图 6 斑尾刺虾虎鱼各体长组食物组成的Bray-Curtis similarity聚类分析图 Fig. 6 Analyze on Bray-Curtis similarity CLUSTER on the dietary composition of Acanthogobius ommaturus in body length groups |
从营养生态位宽度来看, 181—200 mm体长组斑尾刺虾虎鱼由于只采集到1尾且只摄食大弹涂鱼1种食物造成其生态位宽度为0;41—60 mm体长组生态位宽度值为0.86, 61—180 mm体长组的生态位相对较高, 平均值为2.05。其中, 61—120 mm体长组随着体长的增大而增大, 但121—160 mm体长组生态位有所降低(图 7)。

|
| 图 7 不同体长组斑尾刺虾虎鱼的营养生态位宽度 Fig. 7 Trophic niche of Acanthogobius ommaturus in each body length groups |
斑尾刺虾虎鱼作为长江口鱼类的优势种和定居种, 主要栖息于泥底浅海区, 其幼体会经常性随着潮水的涨落而出入潮间带, 有时会进入到高潮滩的植被区[6, 19], 但人们对盐沼生境下的丰度变化和利用盐沼生境的机理性研究极其缺乏。童春富[19]对长江口东滩湿地的海三棱藨草和光滩生境斑尾刺虾虎鱼的体长分布范围做了对比研究, 认为前者生境内斑尾刺虾虎鱼体长范围(20—200 mm)较后者分布更广, 且是数量最多的鱼种, 说明斑尾刺虾虎鱼喜好利用盐沼植被生境。本研究也很好地证实了这一点, 大多数月份在盐沼植被区均采集到斑尾刺虾虎鱼样本, 丰度以6—8月和11月相对较高, 约96%个体为幼体(平均体长109 mm), 平均摄食等级为1.5, 说明斑尾刺虾虎鱼(主要是幼体)会季节性进入长江口盐沼生境进行栖息和摄食, 不同体长的个体摄食强度和饵料种类也差异较大。由于长江口斑尾刺虾虎鱼的产卵期在3—5月, 在邻近潮下带海域产卵且卵产于巢穴内, 亲鱼有护卵习性, 且其寿命为1年[3], 因此在3—5月盐沼生境斑尾刺虾虎鱼丰度很低, 且幼体比例达100%。6—8月是其利用盐沼生境的主要月份, 9—11月丰度逐渐降低, 但个体生物量有所增加, 至冬初(12月)则未在盐沼生境发现斑尾刺虾虎鱼个体出现。Zhang等[20]在长江口淡水区光滩生境同样发现斑尾刺虾虎鱼具有类似的丰度变化规律。综合来说, 盐沼生境内斑尾刺虾虎鱼以幼鱼比例极高, 且夏季是其利用盐沼生境的主要时期, 其次为秋季, 春季和冬季均丰度很低, 其进入盐沼生境的主要目的是摄食。
3.2 饵料组成的月份和体长差异有研究表明, 长江口斑尾刺虾虎鱼(光滩生境)全年主要以虾类(白虾属和磷虾类为主)、鱼类(虾虎鱼类和鮻Liza haematocheila)等饵料生物为食, 属底栖动物食性鱼类[9];渤海的斑尾刺虾虎鱼主要摄食日本鼓虾(Alpheus japonicus)和六丝钝尾虾虎鱼(Amblychaeturichthys hexanema)[21], 莱州湾的斑尾刺虾虎鱼主要摄食仔幼鱼、长尾类和多毛类[8, 15];胶州湾的斑尾刺虾虎鱼主要以虾类、多毛类和鱼类为食, 同时还摄食少量的钩虾和蟹类, 优势饵料生物是鲜明鼓虾(Alpheus distinguendus)、沙蚕和六丝钝尾虾虎鱼[7]。因此, 斑尾刺虾虎鱼食性以虾类、鱼类为主, 本研究也证明了这一点。在饵料组成上, 长江口东北水域盐沼生境斑尾刺虾虎鱼各个月份均以白虾属(或沼虾属)等虾类及大鳍弹涂鱼(或大弹涂鱼)等虾虎鱼科鱼类为优势饵料, 潮间带大型底栖动物优势种(如弓蟹科或相手蟹科等蟹类幼体、双壳类的彩虹明樱蛤及端足类的蜾蠃蜚科sp.和钩虾类未定种)[12, 22]和水体中的浮游动物优势种类(糠虾类)也是其较为喜好的食物。调查发现, 约50%的饵料种类(饵料重量约占31%)属于仅生活于潮间带盐沼植被区或邻近光滩区, 特别是在鱼类饵料类别中, 大弹涂鱼和大鳍弹涂鱼的质量比重高达69%, 且这2种鱼类的生活史主要分布于盐沼植被区[2, 4]。因此本研究推测, 斑尾刺虾虎鱼进入盐沼生境的主要原因是追随白虾属或沼虾属等虾类而进入潮间带植被区, 同时也为了能捕食到适口性更好的、仅生活于潮间带的大弹涂鱼和大鳍弹涂鱼等鱼类以及其他底栖生物。
鱼类食性的月间变化可以反映出鱼体代谢强度、摄食行为和水域环境中饵料生物的月变化, 是鱼类对环境的适应性行为。目前, 针对斑尾刺虾虎鱼食物组成月间或季节变化的报道还极为少见[9]。少量研究表明, 斑尾刺虾虎鱼的食物组成存在明显的季节变化, 在4个季节中主要摄食虾类, 冬季和春季还摄食糠虾类, 对鮻的摄食主要集中在夏季[9]。本研究结果与此结果较为类似, 但发现6—11月针对盐沼生境的底栖生物(如大弹涂鱼和大鳍弹涂鱼等虾虎鱼科以及蟹类)的摄食比例明显高于前人研究, 这反映了盐沼对于斑尾刺虾虎鱼具有重要的生境价值。
众多研究表明, 随着鱼类的生长发育和体长增大, 捕食能力增强, 摄食饵料生物的种类和个体大小也会随之而发生变化, 意味着其摄食饵料生物的范围扩大、数量增加, 这对于鱼类的生存和繁殖后代都有积极的意义[13-14, 23]。朱美贵等[9]认为, 体长小于70 mm的斑尾刺虾虎鱼主要以幼鱼为食;随着体长的增加, 脊尾白虾、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)所占比例逐渐上升;当体长大于189 mm时, 鱼类饵料的比例超过虾类, 成为最主要的饵料生物。本研究与此结果有一定的差异, 发现41—60 mm体长组的个体主要以虾类作为饵料, 其次为蟹类, 而鱼类的生物量比例极低(< 1%)。而随着体长的增加, 饵料中鱼类的比例呈逐渐增高趋势, 成体个体的饵料中鱼类的质量比例达65%以上, 虾类比例则下降明显, 说明幼体发育至成体时其存在明显的食性转换现象。同时, 本研究也发现随着斑尾刺虾虎鱼体长的增加, 其捕食饵料(白虾类和虾虎鱼类)的个体大小和重量也相应增加, 与朱美贵等[9]研究结果较为一致。
3.3 摄食强度和生态位宽度长江口斑尾刺虾虎鱼摄食强度随季节变化显著, 且随体长变化而变化[9]。本研究发现斑尾刺虾虎鱼(体长组41—200 mm)的平均胃饱满指数P (%)和摄食等级基本随体长的增大而增大, 但在141—160 mm体长组其胃饱满指数和摄食等级略有下降, 而朱美贵等[9]也发现150—189 mm体长组个体其摄食强度有所下降, 推测与较大个体的样本数较少有关。一般来说, 较大个体鱼类的摄食等级要明显高于其幼体阶段, 摄食等级3和4的个体比例明显高于幼体(如智利竹筴鱼Trachurus murphyi[13]), 斑尾刺虾虎鱼同样有类似规律。从月变化来看, 5—11月斑尾刺虾虎鱼平均胃饱满指数和摄食等级均呈逐渐增加趋势, 这与其个体大小随月份的增大而增大有关。与大洋性中上层鱼类相比, 斑尾刺虾虎鱼的P (%)月平均值为3.3, 明显高于东南太平洋智利竹筴鱼的0.32[13]和大西洋西部大眼金枪鱼(Thunnus obesus)的0.37[14], 间接说明河口区鱼类的摄食能力和饵料可获得性要相对高于大洋性鱼类。
通过对不同体长组个体的饵料类别进行聚类分析发现, 41—60 mm体长组的饵料组成与61—180 mm的个体差异较大;其营养生态位宽度也仅为0.86, 明显低于后者的2.05。由于181—200 mm体长组个体只采集到1尾且只摄食大弹涂鱼1种食物造成其生态位宽度为0, 有待于以后进一步研究。韩东燕等[7]针对胶州湾浅海区斑尾刺虾虎鱼(平均体长为189 mm)成体的营养生态位宽度进行了研究, 发现其值为2.13, 略高于本研究。本研究还发现, 121—160 mm体长组生态位宽度比81—120 mm体长组有所降低, 这是因为接近成体的斑尾刺虾虎鱼的食物组成中鱼类和虾蟹类比例较高造成, 也可能是样品数相对较少产生的实验误差造成。
由于营养生态位宽度反映了鱼类摄食饵料生物的种类数和均匀度, 也反映了鱼类的摄食能力大小[16]。较小个体(41—60 mm体长组)的斑尾刺虾虎鱼由于摄食能力较弱, 其生态位宽度很低, 而随着个体的增大, 其生态位宽度明显增大(图 7), 这也说明盐沼生境丰富的饵料环境为斑尾刺虾虎鱼提供了良好的摄食场所, 同时春季至秋季盐沼植被茂密的植株也可为幼体提供躲避捕食者的最佳庇护所, 有利于斑尾刺虾虎鱼在秋末变为成体后进入较深水域越冬。
3.4 昼夜差异目前, 尚未有研究报道斑尾刺虾虎鱼丰度和食性组成的昼夜变化。本研究发现, 白天潮水采集的斑尾刺虾虎鱼丰度要高于晚上, 特别是夏季(6—8月);但也发现其平均体长、平均体质量和摄食等级均要略低于晚上。白天样本的饵料类别中鱼类的重量百分比显著高于夜晚, 说明斑尾刺虾虎鱼在白天比夜晚更容易摄食到盐沼生境活动的虾虎鱼类[2], 作者在渔获中也发现白天潮水采集的大鳍弹涂鱼数量明显高于夜晚。而虾类饵料的质量百分比则是夜晚显著高于白天, 这也与白虾属等虾类在夜晚出现的数量更高有关。蟹类饵料的质量百分比昼夜差异极小, 与斑尾刺虾虎鱼较少摄食潮间带蟹类饵料有关。因此可知, 夏季斑尾刺虾虎鱼的丰度和摄食习性具有明显的昼夜节律变化, 白天比晚上有更多的个体进入盐沼生境进行摄食活动, 且白天更趋向于摄食鱼类, 而夜晚趋向于摄食虾类。
| [1] | 张衡, 何文珊, 童春富, 陆健健. 长江口低盐淡水区潮间带鱼类群落结构季节及半月相变化. 应用生态学报, 2008, 19(5): 1110–1116. |
| [2] | 倪勇, 王幼槐, 许成玉, 熊国强. 上海鱼类志. 上海: 上海科学与技术出版社, 1990. |
| [3] | 庄平, 王幼槐, 李圣法, 邓思明, 李长松, 倪勇. 长江口鱼类. 上海: 上海科学与技术出版社, 2006. |
| [4] | 张衡, 朱国平. 长江河口潮间带鱼类群落的时空变化. 应用生态学报, 2009, 20(10): 2519–2526. |
| [5] | 张衡, 全为民, 陈渊戈, 崔雪森. 长江口口门区潮下带水域鱼类群落组成的季节变化. 长江流域资源与环境, 2014, 23(11): 1534–1539. |
| [6] | 张衡, 何文珊, 童春富, 陆健健. 崇西湿地冬季潮滩鱼类种类组成及多样性分析. 长江流域资源与环境, 2007, 16(3): 308–313. DOI:10.3969/j.issn.1004-8227.2007.03.009 |
| [7] | 韩东燕, 薛莹, 纪毓鹏, 徐宾铎, 刘贺, 麻秋云. 胶州湾5种虾虎鱼类的营养和空间生态位. 中国水产科学, 2013, 20(1): 148–156. |
| [8] | 范海洋, 纪毓鹏, 张士华, 苑春亭, 高天翔. 黄河三角洲斑尾复虾虎鱼渔业生物学的研究. 中国海洋大学学报, 2005, 35(5): 733–736. |
| [9] | 朱美贵, 杨刚, 张涛, 赵峰, 彭欣悦, 闫光松, 庄平. 长江口斑尾刺虾虎鱼的摄食习性. 中国水产科学, 2016, 23(4): 914–923. |
| [10] | 国家海洋局. GB/T 12763.6-2007海洋调查规范第6部分:海洋生物调查. 北京: 中国标准出版社, 2008. |
| [11] | 刘文亮, 何文珊. 长江河口大型底栖无脊椎动物. 上海: 上海科学技术出版社, 2007. |
| [12] | 张衡, 叶锦玉, 梁晓莉, 朱小静, 靳少非, 陈渊戈, 张佳蕊, 戴阳. 长江口东滩湿地芦苇生境大型底栖无脊椎动物群落结构的月动态. 应用生态学报, 2017, 28(4): 1360–1369. |
| [13] | 张衡, 张胜茂, 王雪辉, 唐峰华, 杨胜龙, 化成君, 何宗会, 马海有. 东南太平洋秋冬季智利竹筴鱼摄食习性的初步分析. 海洋渔业, 2013, 35(2): 161–167. DOI:10.3969/j.issn.1004-2490.2013.02.006 |
| [14] | 朱国平, 周应祺, 许柳雄, 姜文新. 大西洋西部大眼金枪鱼摄食生态的初步研究. 水产学报, 2007, 31(1): 23–30. |
| [15] | 孟田湘. 莱州湾黄河口近岸幼鱼食物结构与饵料重叠. 海洋水产研究, 2000, 21(2): 1–6. |
| [16] | Bearhop S, Adams C E, Waldron S, Fuller R A, Macleod H. Determining trophic niche width:a novel approach using stable isotope analysis. Journal of Animal Ecology, 2004, 73(5): 1007–1012. DOI:10.1111/j.0021-8790.2004.00861.x |
| [17] | Marshall S, Elliott M. A comparison of univariate and multivariate numerical and graphical techniques for determining inter-and intraspecific feeding relationships in estuarine fish. Journal of Fish Biology, 1997, 51(3): 526–545. DOI:10.1111/jfb.1997.51.issue-3 |
| [18] | Clarke K R, Warwick R M. A further biodiversity index applicable to species lists:variation in taxonomic distinctness. Marine Ecology Progress Series, 2001, 216: 265–278. DOI:10.3354/meps216265 |
| [19] | 童春富. 长江河口潮间带盐沼植被分布区及邻近光滩鱼类组成特征. 生态学报, 2012, 32(20): 6501–6510. |
| [20] | Zhang H. Diel, semi-lunar and seasonal patterns in the fish community of an intertidal zone of the Yangtze estuary. Journal of Applied Ichthyology, 2013, 29(6): 1252–1258. DOI:10.1111/jai.2013.29.issue-6 |
| [21] | 杨纪明. 渤海鱼类的食性和营养级研究. 现代渔业信息, 2001, 16(10): 10–19. DOI:10.3969/j.issn.1004-8340.2001.10.002 |
| [22] | 全为民, 赵云龙, 朱江兴, 施利燕, 陈亚瞿. 上海市潮滩湿地大型底栖动物的空间分布格局. 生态学报, 2008, 28(10): 5179–5187. DOI:10.3321/j.issn:1000-0933.2008.10.067 |
| [23] | 陈大刚. 渔业资源生物学. 北京: 中国农业出版社, 1997. |