 2018, Vol. 38
2018, Vol. 38文章信息
- 单文俊, 邢亚娟, 闫国永, 韩士杰, 张军辉, 王庆贵.
- SHAN Wenjun, XING Yajuan, YAN Guoyong, HAN Shijie, ZHANG Junhui, WANG Qinggui.
- 长白山天然次生林土壤微生物量对氮沉降的响应
- Response of soil microbial biomass to nitrogen deposition in natural secondary forests in Changbai Mountains
- 生态学报. 2018, 38(17): 5996-6005
- Acta Ecologica Sinica. 2018, 38(17): 5996-6005
- http://dx.doi.org/10.5846/stxb201706091048
-
文章历史
- 收稿日期: 2017-06-09
- 网络出版日期: 2018-05-30





2. 黑龙江省林业科学研究所, 哈尔滨 150081;
3. 中国科学院沈阳应用生态研究所, 沈阳 110016
2. Institute of Forestry Science of Heilongjiang Province, Harbin 150081, China;
3. Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016, China
随着人口攀升和人类活动加剧, 工业化发展迅速, 大规模使用化石燃料和肥料, 导致大气活性氮含量快速增加[1], 再通过降雨、降尘等方式重新回到地球表面形成氮沉降[2]。与19世纪工业革命起始阶段相比, 现时大气氮沉降量增加近两倍[3], 并呈逐年上涨趋势[4]。预测到2050年, 大气氮沉降量将再翻一番[3]。我国成为继欧洲和北美之后的世界三大氮沉降区之一[5], 形势不容乐观。
氮沉降增加对陆地生态系统有显著的影响[6]。森林作为陆地生态系统的重要组成部分, 对人类社会具有很高的服务价值。大气氮沉降影响森林生态系统碳、氮循环, 对其功能产生影响。研究表明, 氮沉降加剧会降低植物多样性[6]、改变土壤微生物量和群落结构[7]、减缓凋落物分解速率[8]、导致土壤酸化[9]、影响森林养分循环[10]等。土壤微生物种类多、数量大, 在森林生态系统养分循环中扮演驱动者的角色, 对土壤养分循环具有重要作用[11]。土壤微生物量碳(MBC)、氮(MBN)可作为衡量土壤微生物量大小的标准之一, 也可作为土壤肥力状况的生物学指标, 并且与其他肥力指标如土壤有机质、全氮(TN)、全磷(TP)、速效钾和碱解氮等存在正相关关系[12]。氮沉降加剧, 引起土壤酸化, Al3+、Mn2+等有害金属离子含量上升, 土壤缓冲能力下降, 造成土壤碳氮比失衡等负面影响, 导致土壤微生物量下降、功能减弱, 对森林生态系统产生不利影响[13]。也有研究指出, 氮沉降对土壤微生物量有促进作用或无影响[14]。
天然次生林是森林资源中面积最大的类型, 其面积一直在扩大。经营好次生林, 不仅可以缓解我国木材紧缺问题, 增加以森林为栖息环境的动植物资源, 而且在维持与提高环境效益方面也具有重要意义。本研究以长白山白桦(Betula platyphylla)山杨(Populus davidiana)天然次生林为研究对象, 模拟不同氮沉降量对次生林土壤微生物量的影响, 探讨温带次生林微生物量与养分循环对氮沉降的响应, 进而揭示氮沉降对温带次生林生态系统功能的影响, 为评估次生林对氮沉降的微生物响应机制提供基础数据。
1 材料与方法 1.1 样地概况及氮添加控制试验本研究控制试验样地位于吉林省白山市露水河林业局(127°29′—128°02′E, 42°24′—42°49′N), 林型为白桦山杨天然次生林。研究地区为典型的大陆性季风气候, 冬季漫长、寒冷, 夏季降水多且气温较高, 春秋两季较短。年平均气温为2.7℃, 年平均降水量为872 mm, 海拔922.5 m, 土壤为山地暗棕色森林土。本研究于2006年开始进行氮添加控制试验, 选择海拔、坡度、植被组成相对均质的天然白桦山杨林地布设9块样地, 样地面积30 m×30 m, 样地之间间隔10 m, 防止土壤氮联通。控制试验共设计3个处理, 即对照(0 kg N hm-2 a-1)、低氮(LN, 25 kg N hm-2 a-1)和高氮(HN, 50 kg N hm-2 a-1), 每个处理重复3次。每年在生长季5—9月上旬施用NH4NO3, 为消除样地间水分差异, 对照样地喷洒同量纯净水。试验样地pH(4.84±0.58)、TN((7.58±1.25) g/kg)、总有机碳(TOC)((140.3±11.6) g/kg)、TP((1.40±0.52) g/kg)[15]。
1.2 样品采集与测试方法土壤样品采集时间分别为2016年春(5月)、夏(7月)、秋(9月)3个季节。每块样地随机取5个点, 去除枯枝落叶层。取土深度分为上层(0—10 cm)和下层(10—20 cm), 将5个点的样品混合, 去除植物根茎和碎石等杂物。混合样品运输途中冰鲜保存, 带回实验室立即测定土壤MBC和MBN。
土壤MBC、MBN测定采用氯仿熏蒸浸提法。浸提样品使用德国耶拿Multi N/C® 3100总有机碳/总氮分析仪进行测定[16-17]。采用如下公式计算:
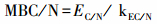
|
式中, EC/N为熏蒸土壤减未熏蒸土壤, kEC/N为转换系数, 土壤MBC和MBN均为0.45[18]。
土壤pH用酸度计测量。土壤全碳(TC)采用干烧法, 取1 g干样品放入HT1500燃烧炉中燃烧。在高纯氧(99.99%)环境中, 样品在1200℃下充分燃烧, 用Multi N/C® 3100总有机碳/总氮分析仪对释放的CO2进行测定, 每个样品3—5 min, 重复3次。数据由电脑自动记录。TN用硫酸双氧水硝煮法[19], 硝煮样品经过定容后用Multi N/C® 3100总有机碳/总氮分析仪测定。TP采用酸溶-钼锑抗比色法。土壤熏蒸前后浸提液通过Multi N/C® 3100总有机碳/总氮分析仪测定土壤可溶性有机碳(DOC)、氮(DON)。
1.3 数据处理方法应用Excel 2010进行数据录入和简单处理。利用SPSS 20.0软件进行数据分析。利用ANOVA方差分析对土壤MBC和MBN、pH、TC、TN、TP以及DOC、DON进行季节和氮处理间差异显著性比较。用LSD多重比较分析同一处理季节间和氮处理间的土壤MBC、MBN的差异。
2 结果与分析 2.1 土壤理化性质对氮沉降的响应由表 1可见, 上层土壤pH在春、夏两季氮处理下(LN、HN)显著低于对照处理(CK), 秋季HN显著低于其他两个处理。土壤TC春季CK和LN显著高于HN, 夏季LN低于CK和HN, 秋季HN显著高于CK和LN。春、夏两季氮处理下(LN、HN)土壤TN显著高于CK, 秋季3种处理差异均不显著。HN和LN处理下土壤TP显著高于CK, 春、夏两季差异显著, 秋季差异不显著。DOC和DON在氮处理下, 只在夏季和秋季与CK存在差异。氮处理导致下层土壤pH值下降。LN对土壤TC和TN有促进作用。氮处理在夏季对土壤TP有促进作用, 秋季有抑制作用。氮处理对下层土壤DOC和DON没有显著影响(表 2)。
| 土壤理化性质 physical-chemical properties of soil |
处理 Treatment |
春季 Spring |
夏季 Summer |
秋季 Autumn |
| pH | CK | 5.54±0.01A | 5.62±0.03A | 5.57±0.04a |
| LN | 5.38±0.04B | 5.47±0.01B | 5.55±0.03a | |
| HN | 5.34±0.02B | 5.47±0.04B | 5.43±0.0.9b | |
| 全碳TC | CK | 117.00±8a | 123.00±7A | 80.00±6b |
| Total carbon/(g/kg) | LN | 103.00±10ab | 72.00±2B | 78.00±9b |
| HN | 98.00±8b | 113.00±5A | 93.00±5a | |
| 全氮TN | CK | 4.30±0.04B | 6.74±0.44b | 6.54±0.30a |
| Total nitrogen/(g/kg) | LN | 6.36±0.78Ab | 7.92±0.26a | 6.96±0.38a |
| HN | 7.86±0.56Aa | 8.00±0.76a | 7.62±0.54a | |
| 全磷TP | CK | 0.71±0.06B | 0.89±0.06Ba | 0.95±0.05a |
| Total phosphorus/(g/kg) | LN | 1.45±0.07A | 1.00±0.10ABb | 0.84±0.04a |
| HN | 1.39±0.05A | 1.20±0.06Aa | 0.97±0.09a | |
| 可溶性有机碳DOC | CK | 370.00±95a | 427.00±45Bc | 451.00±39ABb |
| Dissolved organic carbon/(mg/kg) | LN | 365.00±42a | 508.00±37Aa | 437.00±31Ba |
| HN | 321.00±43a | 479.00±45ABb | 513.00±28Aa | |
| 可溶性有机氮DON | CK | 38.00±3a | 58.00±9a | 22.00±0b |
| Dissolved organic nitrogen/(mg/kg) | LN | 39.00±6a | 51.00±5a | 23.00±2b |
| HN | 39.00±7a | 47.00±6b | 26.00±1a | |
| CK:对照处理, Stands for the control treatment;LN:低氮处理, Low nitrogen deposition treatment;HN:高氮处理, High nitrogen deposition treatment | ||||
| 土壤理化性质 Physical-chemical properties of soil |
处理 Treatment |
春季 Spring |
夏季 Summer |
秋季 Autumn |
| pH | CK | 5.79±0.14Aa | 5.81±0.12Aa | 5.67±0.06A |
| LN | 5.59±0.02ABb | 5.60±0.01ABb | 5.67±0.02A | |
| HN | 5.43±0.01Ba | 5.46±0.08Ba | 5.53±0.03B | |
| 全碳TC | CK | 55.00±13Aa | 45.00±3B | 32.00±3B |
| Total carbon/(g/kg) | LN | 34.00±2ABb | 57.00±4A | 46.00±3A |
| HN | 26.00±3Ba | 44.00±2B | 29.00±2B | |
| 全氮TN | CK | 2.60±0.22B | 3.40±0.266C | 3.82±0.46B |
| Total nitrogen/(g/kg) | LN | 3.44±0.38A | 5.84±0.18A | 5.46±0.20A |
| HN | 3.02±0.04A | 4.60±0.60B | 3.96±0.54B | |
| 全磷TP | CK | 1.02±0.30a | 1.03±0.06B | 1.13±0.04A |
| Total phosphorus/(g/kg) | LN | 1.00±0.09a | 1.29±0.06A | 0.98±0.06B |
| HN | 0.80±0.06a | 1.28±0.05A | 0.97±0.05B | |
| 可溶性有机碳DOC | CK | 208.00±39a | 488.00±55b | 501.00±31a |
| Dissolved organic carbon /(mg/kg) | LN | 205.00±50a | 490.00±35b | 502.00±59a |
| HN | 254.00±47a | 583.00±65a | 560.00±44a | |
| 可溶性有机氮DON | CK | 18.00±1a | 50.00±4Aa | 16.00±2a |
| Dissolved organic nitrogen /(mg/kg) | LN | 15.00±1a | 36.00±3Bb | 14.00±2a |
| HN | 17.00±2a | 46.00±7ABa | 16.00±2a |
如图 1所示, 连续10年氮添加, 上层土壤MBC在春季氮处理下(LN和HN)显著低于CK, LN较CK下降6.2%, HN较CK下降20.8%;夏季CK和LN差异不显著, 与HN差异极显著, HN较CK降低46.9%;秋季HN较CK升高14.2%, LN较CK降低23.3%(图 1)。下层土壤MBC春季LN和CK差异不显著, 与HN差异极显著, CK与HN差异显著, LN较CK升高1.9%, HN较CK降低5.6%;夏季3种处理之间均差异极显著, LN较CK上升37.1%, HN较CK降低17.2%;秋季CK与LN和HN都表现为极显著差异, HN、LN较CK都降低24.3%(图 1)。

|
| 图 1 氮沉降对土壤微生物量碳的影响 Fig. 1 Effects of nitrogen deposition on soil microbial biomass carbon |
如图 2所示, 上层土壤MBN春季LN与CK差异极显著, HN与CK差异显著, LN较CK上升11.6%, HN较CK上升6.1%;夏季3种处理之间差异都极显著, LN较CK降低9.3%, HN较CK降低48.9%;秋季3种处理之间差异都极显著, LN较CK上升45.2%, HN较CK升高34.2%(图 2)。下层土壤MBN春季HN与CK差异极显著, LN与CK差异显著, LN较CK上升5.1%, HN较CK上升7.7%;夏季3种处理之间均差异极显著, LN较CK下降37.2%, HN较CK下降8.3%;秋季HN与CK、LN都差异极显著, HN较CK升高75.7%, LN较CK升高3.5%(图 2)。

|
| 图 2 氮沉降对土壤微生物量氮的影响 Fig. 2 Effects of nitrogen deposition on soil microbial biomass nitrogen |
由表 3可见, 春季上层土壤MBC/MBN, 3种处理之间差异极显著, LN比CK降低0.55, HN比CK降低0.88;下层土壤LN比CK降低0.12, 差异不显著, HN比CK降低0.47, 差异极显著。夏季上层土壤MBC/MBN, LN与HN差异显著, 与CK存在极显著差异, LN比CK升高0.65, HN比CK升高0.18;下层土壤, LN比CK升高5.24, 差异极显著, HN比CK降低0.48, 差异显著。秋季上层土壤MBC/MBN, 3种处理之间差异极显著, LN比CK降低2.85, HN比CK降低0.89;下层土壤3种处理之间差异都极显著, LN比CK降低5.01, HN比CK降低8.05。总体来看, 氮沉降降低了春季和秋季土壤MBC/MBN, 提高了夏季土壤MBC/MBN。季节动态表现为:CK土壤微生物量从春季到秋季表现出升高趋势, 而氮添加处理(LN和HN)都表现为先升高后降低的趋势。
| 处理 Treatment |
春季Spring | 夏季Summer | 秋季Autumn | |||||
| 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | 0—10 cm | 10—20 cm | |||
| CK | 3.47±0.03A | 3.77±0.14A | 5.37±0.05Ba | 4.56±0.13Ba | 6.13±0.26A | 14.08±0.74A | ||
| LN | 2.92±0.08B | 3.65±0.08A | 6.02±0.14Aa | 9.80±0.32A | 3.28±0.19C | 9.07±1.18B | ||
| HN | 2.59±0.05C | 3.30±0.04B | 5.55±0.27Ab | 4.07±0.07Bb | 5.24±0.16B | 6.03±0.33C | ||
由表 4可见, 氮添加(N, N addition)、季节差异(S, Season)和土层深度(D, Depth)对土壤微生物量影响显著, 且两两因素之间存在交互作用, 3种因素间互作(N×S×D)影响显著。说明氮添加处理并不是唯一影响土壤微生物量的因素, 季节差异和土层深度也是影响土壤微生物量的主要因素。氮添加、季节差异和土层深度都是影响土壤pH的主要因素, N×S对土壤pH影响极显著, N×D、S×D, 以及N×S×D对土壤pH的影响都不显著。
| 变异来源 Source of variation |
土壤微生物量碳 Microbial biomass carbon |
土壤微生物量氮 Microbial biomass nitrogen |
pH | |||
| F | P | F | P | F | P | |
| N添加N addition(N) | 548.431 | <0.01 | 22.420 | <0.01 | 38.275 | <0.01 |
| 季度Season(S) | 2235.435 | <0.01 | 5980.523 | <0.01 | 6.618 | <0.01 |
| 土壤深度Depth(D) | 18430.506 | <0.01 | 13617.934 | <0.01 | 24.607 | <0.01 |
| N添加×季度(N×S) | 560.510 | <0.01 | 141.365 | <0.01 | 4.094 | <0.01 |
| N添加×土壤深度(N×D) | 142.784 | <0.01 | 146.992 | <0.01 | 2.221 | >0.05 |
| 季度×土壤深度(S×D) | 1008.880 | <0.01 | 2279.391 | <0.01 | 3.118 | >0.05 |
| N添加×季度×土壤深度(N×S×D) | 249.619 | <0.01 | 66.364 | <0.01 | 0.402 | >0.05 |
由表 5可见, 各因素与土壤MBC相关分析得出, 氮添加处理的上、下两层土壤MBC与pH存在负相关关系。CK的上层土壤MBC与TC正相关(P < 0.01), 与TN和TP负相关(P < 0.05)。而LN的TP与MBC正相关(P < 0.01)。DOC在上层土壤HN处理下与MBC负相关(P < 0.01)。上层土壤DON与MBC在LN处理下正相关(P < 0.01), 下层土壤DON与MBC在CK和LN处理下均存在正相关关系(P < 0.01)。
| 微生物量碳Microbial biomass carbon | |||||||
| 0—10 cm | 10—20 cm | ||||||
| CK | LN | HN | CK | LN | HN | ||
| pH | 0.021 | -0.859** | -0.715 | 0.465 | -0.741 | -0.694* | |
| 全碳TC | 0.860** | 0.421 | -0.661 | 0.441 | 0.519 | 0.254 | |
| 全氮TN | -0.668* | -0.043 | -0.224 | -0.315 | 0.236 | -0.100 | |
| 全磷TP | -0.809** | 0.776* | 0.455 | -0.142 | 0.957** | 0.099 | |
| 可溶性有机碳DOC | -0.439 | -0.194 | -0.946** | -0.032 | 0.008 | -0.640 | |
| 可溶性有机氮DON | 0.374 | 0.860** | -0.499 | 0.866** | 0.894** | 0.443 | |
| 微生物量氮MBN | 0.934** | 0.689* | 0.933** | 0.806** | 0.316 | 0.958** | |
| * *表示极显著相关(P<0.01);*表示显著相关(P<0.05) | |||||||
由表 6可见, 氮处理的上层土壤LN的MBN与pH负相关(P<0.01), 下层土壤HN的pH与之正相关(P<0.01)。上层土壤CK的TP与MBN存在负相关(P<0.01), LN的TP与之存在正相关(P<0.05)。氮处理的上、下层土壤DOC与MBN均负相关(P<0.01), 上、下层土壤CK的TN均与MBN负相关(P<0.05), 下层土壤LN的TN与MBN负相关(P<0.01)。总体来看, MBC与MBN之间存在显著的正相关关系。
| 微生物量碳Microbial biomass carbon | |||||||
| 0—10 cm | 10—20 cm | ||||||
| CK | LN | HN | CK | LN | HN | ||
| pH | -0.218 | -0.878** | -0.740* | 0.444 | 0.777* | 0.804** | |
| 全碳TC | 0.651 | 0.885** | -0.384 | 0.791* | -0.595 | 0.007 | |
| 全氮TN | -0.865** | -0.605 | -0.033 | -0.720* | -0.807** | -0.3522 | |
| 全磷TP | -0.900** | 0.952** | 0.720* | -0.335 | 0.411 | -0.156 | |
| 可溶性有机碳DOC | -0.524 | -0.771* | -0.950** | -0.595 | -0.894** | -0.815** | |
| 可溶性有机氮DON | 0.053 | 0.292 | -0.216 | 0.476 | -0.081 | 0.210 | |
| 微生物量碳MBC | 0.934** | 0.689* | 0.933** | 0.806** | 0.316 | 0.958** | |
| * *表示极显著相关(P<0.01);*表示显著相关(P<0.05) | |||||||
氮沉降对土壤理化性质的影响主要表现为土壤酸化, 显著降低土壤pH。产生的NH4+和NO3-导致土壤酸化, 在硝化过程中1 mol NH4+被转化为NO3-产生2 mol H+, 故NH4+沉降比NO3-沉降更能促进土壤酸化, 导致土壤pH降低[9, 13]。本研究氮沉降使土壤pH降低, 且氮沉降对土壤pH的影响并没有随着土层深度而减弱, 这可能是由于长期模拟氮添加导致NH4+、NO3-淋溶, 对下层土壤影响也很明显[20]。本研究土壤pH存在季节性变化, 这可能和该地区季节性气候变化以及植物生长对养分吸收有关[21]。
土壤TC和DOC都可作为表征土壤碳库的重要指标[22], Silveira研究显示氮沉降对土壤TC的影响并不显著, 这可能是因为土壤总有机碳库变化是长期过程, 而且施氮在提高植物初级生产力的同时, 也限制了有机质的分解[23]。周纪东对内蒙草原氮添加频度和强度研究也得出了相同结论[24]。土壤DOC是土壤活性有机质, 可以被土壤微生物或植物根系吸收利用, 有研究显示土壤DOC并不随氮沉降发生规律性变化[25]。有研究则指出氮沉降对土壤DOC含量有抑制作用, 且随着氮沉降量的增加而加强[26]。Nadelhoffer试验则得出了相反的结论, 氮沉降导致土壤DOC含量增加[27]。本研究土壤DOC含量随季节变化表现不同, 可能受到不同季节土壤温度和水分、微生物、根分泌物、凋落物量等因素的影响。
土壤氮素是植物生长的重要元素之一, 土壤TN和DON也是衡量土壤氮肥的重要指标。有研究显示, 氮沉降并没有增加土壤TN, 可能是土壤微生物和植物对外源氮的吸收和氮的挥发, 从而导致TN维持在稳定水平[28-29]。本研究氮沉降对上层土壤TN影响不显著, 但LN促进了下层土壤TN, 这可能是因为氮素的长期淋溶和下层土壤生物作用减弱导致土壤下层氮积累。闫聪微等研究表明土壤DON随氮沉降增加而增加[30], 但元晓春等研究指出, 氮沉降对土壤DON没有显著影响[31]。本研究与元晓春等的研究结论相似, 这可能是由于土壤中氮的矿化速率随氮沉降增大而增加所导致。
土壤磷被认为是森林生态系统除氮限制外的第二大限制因子[32]。氮沉降主要通过改变土壤碳氮比、微生物组成、与磷循环有关酶类和土壤铁、铝离子流动性而改变土壤磷循环[33]。张继舟研究指出, 氮沉降并没有改变土壤TP[34], 也有研究指出氮沉降增加了土壤TP[35]或降低[25]。本研究氮沉降在春、夏两季增加TP, 但秋季对下层土壤TP有抑制作用。
3.2 氮沉降对土壤MBC、MBN的影响氮沉降主要通过改变土壤理化性质、酶活性、微生物群落结构和凋落物分解速率来改变土壤微生物量。本研究土壤微生物量随土层深度而减少, 这与周嘉聪结论一致[36]。随着土壤层加深, 土壤中有机质含量降低、土壤微生物可利用物质含量下降、气体交换减少等导致微生物含量下降。
本研究土壤微生物量具有明显的季节动态, 且MBC和MBN表现不一致。MBC和MBN春季最高、秋季最低, 与赵玉涛等[37]在该试验样地的结论相似。可能是因为春季植物刚开始生长, 对土壤中营养物质的需求并不强烈, 到夏季生长旺盛阶段, 对营养物质的吸收加强, 微生物可利用营养物质含量下降导致微生物量下降。秋季下降可能是由于温度限制了土壤微生物生长。在中国南方地区, 由于秋季温度不是主要限制因子, 周世兴等[38]得出土壤微生物量反而升高, 原因是秋季凋落物增加了土壤中微生物可利用营养物质。
本研究中, 氮沉降量对土壤微生物量影响也有显著作用。LN使上层土壤MBC降低, 但对下层土壤在春、夏两季有促进作用, HN则抑制MBC, 这与郭依秋等[39]得出的结论相似。Jana等[40]在哈佛森林试验表明, 氮沉降降低MBC可能与土壤微生物群落结构发生变化有关。MBC降低可能是由于氮添加使得土壤酸化[41], 随着添加氮的向下淋溶, 底层土壤中可利用氮含量增加, 进而对下层土壤微生物产生促进作用[42]。氮添加对MBN在春、秋两季有促进作用, 但是在夏季有抑制作用。涂利华等[43]研究指出, 氮添加增加了不同季节土壤MBN, 本研究中夏季的研究结果与其存在差异。氮沉降导致土壤MBC和MBN反映不一致可能是由于土壤微生物群落结构发生变化[44]。氮添加具有营养富集效应, 可使喜氮微生物或者利用小分子物质的微生物优先生长, 氮沉降通过对土壤碳、氮转换酶的抑制或促进而改变土壤微生物量, 特别是与木质素与纤维素分解有关的酶[45-46]。氮沉降还可以通过改变凋落物中碳氮比和木质素/氮, 进而影响微生物对凋落物的分解。氮沉降早期, 土壤酶、凋落物长期处于氮限制状态, 氮添加可能对微生物分解凋落物产生促进作用, 导致微生物量增加[47-48];而在氮饱和地区则会出现抑制作用。
3.3 氮沉降背景下影响土壤微生物量的其他因素森林土壤微生物量对氮沉降的响应还受到林型、土壤采集时间和深度等因素的影响。郭萍萍等研究指出, 氮沉降降低了0—10 cm土壤微生物量, 但对下层土壤没有影响[49]。Grayston指出不同植物种类具有不同的微生物群落, 由于不同植物分泌酶等物质的不同, 根系周围的微生物存在差异[50]。土壤微生物量存在明显的季节变化, 微生物的生长与土壤的干湿交替、植物生长对养分的利用等有关。有研究指出, 在生长季土壤微生物量的峰值出现在夏季, 因为夏季具有较好的水热条件, 有利于微生物生长[41, 51]。有研究则得出不同结论, 夏季由于植物生长旺盛, 对土壤养分利用率较高, 微生物可利用养分受到限制, 导致生物量降低;秋季植物生长速度下降, 对养分需求减弱, 土壤微生物可利用养分增加, 导致生物量增加[52]。本研究由春季到秋季, 土壤微生物量呈递减趋势, 可能是因为在春季土壤中可利用的氮素经过冬季的积累, 可充分满足土壤微生物生长所需, 且土壤湿度在冬季融雪后也得到改善;夏季由于干湿交替加剧, 植物生长对养分消耗增加, 土壤微生物量下降;秋季由于植物生长减慢、气温降低, 进一步抑制了土壤微生物生长。有研究也表明长白山天然次生林土壤有效氮含量随生长季而呈递减趋势[15]。氮添加强度对土壤微生物量也会产生不同影响, 表现出LN促进和HN抑制[49, 24], 也有研究发现LN和HN处理同时促进或同时抑制[53]。
4 结论长白山天然次生林土壤pH在氮沉降的作用下降低。上层土壤TC、TN在氮沉降下变化较小, 却显著增加了下层土壤TC、TN的含量。氮沉降增加了土壤春、夏两季TP含量, 但在秋季LN处理对TP有抑制作用, 且下层土壤作用显著。氮沉降对土壤DOC、DON的影响不显著。
上层土壤MBC春季到秋季为递减趋势, 下层土壤春季到秋季为先升后降趋势, 且上、下层土壤差异显著。氮处理在春季对上层土壤MBC有抑制作用, HN在夏季表现出抑制作用, 在秋季有促进作用, LN在秋季有抑制作用。在春、夏两季LN对下层土壤MBC有促进作用, 秋季有抑制作用, HN在3个季节都有抑制作用。土壤MBN表现为上、下层土壤由春季到秋季递减的趋势, 且上、下层土壤MBN差异显著。氮处理促进了春、秋两季土壤MBN, 但是在夏季有抑制作用。氮沉降降低了春季和秋季土壤MBC/MBN, 提高了夏季土壤MBC/MBN。
氮添加强度、季节变化和土层深度对土壤微生物量的影响都很显著, 且三者之间的互作明显, 说明氮沉降对长白山天然次生林土壤微生物量的影响并非单独作用。在同一区域研究中, 氮沉降对土壤微生物量的影响表现出时空动态, 即时间动态表现为不同季节, 空间动态表现为不同土层。未来研究在重视年际变化的同时, 也要注重时空动态对氮沉降作用表现出的差异。
| [1] | Pinder R W, Davidson E A, Goodale C L, Greaver T L, Herrick J D, Liu L L. Climate change impacts of US reactive nitrogen. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(20): 7671–7675. DOI:10.1073/pnas.1114243109 |
| [2] | Liu X J, Zhang Y, Han W X, Tang A H, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459–462. DOI:10.1038/nature11917 |
| [3] | Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle:recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889–892. DOI:10.1126/science.1136674 |
| [4] | Hůnová I, Kurfürst P, Stráník V, Modlík M. Nitrogen deposition to forest ecosystems with focus on its different forms. Science of the Total Environment, 2017, 575: 791–798. DOI:10.1016/j.scitotenv.2016.09.140 |
| [5] | 樊后保, 黄玉梓, 袁颖红, 李燕燕, 黄荣珍, 樊海燕. 森林生态系统碳循环对全球氮沉降的响应. 生态学报, 2007, 27(7): 2997–3009. DOI:10.3321/j.issn:1000-0933.2007.07.042 |
| [6] | Botez F, Postolache C. Nitrogen deposition impact on terrestrial ecosystems. Romanian Biotechnological Letters, 2013, 18(6): 8723–8742. |
| [7] | Li J, Li Z A, Wang F M, Zou B, Chen Y, Zhao J, Mo Q F. Effects of nitrogen and phosphorus addition on soil microbial community in a secondary tropical forest of China. Biology and Fertility of Soils, 2015, 51(2): 207–215. DOI:10.1007/s00374-014-0964-1 |
| [8] | Mo J M, Fang H, Zhu W X, Zhou G Y, Lu X K, Fang Y T. Decomposition responses of pine (Pinus massoniana) needles with two different nutrient-status to N deposition in a tropical pine plantation in southern China. Annals of Forest Science, 2008, 65(4): 405–405. DOI:10.1051/forest:2008017 |
| [9] | Lu X K, Mao Q G, Gilliam F S, Luo Y Q, Mo J M. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Global Change Biology, 2014, 20(12): 3790–3801. DOI:10.1111/gcb.12665 |
| [10] | Menge D N L, Hedin L O, Pacala S W. Nitrogen and phosphorus limitation over long-term ecosystem development in terrestrial ecosystems. PLoS One, 2012, 7(8): e42045. DOI:10.1371/journal.pone.0042045 |
| [11] | Pignataro A, Moscatelli M C, Mocali S, Grego S, Benedetti A. Assessment of soil microbial functional diversity in a coppiced forest system. Applied Soil Ecology, 2012, 62: 115–123. DOI:10.1016/j.apsoil.2012.07.007 |
| [12] | 张海燕, 肖延华, 张旭东, 李军, 席联敏. 土壤微生物量作为土壤肥力指标的探讨. 土壤通报, 2006, 37(3): 422–425. DOI:10.3321/j.issn:0564-3945.2006.03.002 |
| [13] | 单文俊, 王庆贵, 闫国永, 邢亚娟. 基于土壤微生物的碳氮互作效应综述. 中国农学通报, 2016, 32(23): 65–71. DOI:10.11924/j.issn.1000-6850.casb16010056 |
| [14] | 赵超, 彭赛, 阮宏华, 张雅坤. 氮沉降对土壤微生物影响的研究进展. 南京林业大学学报:自然科学版, 2015, 39(3): 149–155. |
| [15] | 胡艳玲, 韩士杰, 李雪峰, 赵玉涛, 李东. 长白山原始林和次生林土壤有效氮含量对模拟氮沉降的响应. 东北林业大学学报, 2009, 37(5): 36-38, 42-42. DOI:10.3969/j.issn.1000-5382.2009.05.013 |
| [16] | Brookes P C, Kragt J F, Powlson D S, Jenkinson D S. Chloroform fumigation and the release of soil nitrogen:the effects of fumigation time and temperature. Soil Biology and Biochemistry, 1985, 17(6): 831–835. DOI:10.1016/0038-0717(85)90143-9 |
| [17] | 林先贵. 土壤微生物研究原理与方法. 北京: 高等教育出版社, 2014: 73–77. |
| [18] | Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703–707. DOI:10.1016/0038-0717(87)90052-6 |
| [19] | 李宇庆, 陈玲, 赵建夫. 土壤全氮测定方法的比较. 广州环境科学, 2006, 21(3): 28–29. |
| [20] | Pietri J C A, Brookes P C. Relationships between soil pH and microbial properties in a UK arable soil. Soil Biology and Biochemistry, 2008, 40(7): 1856–1861. DOI:10.1016/j.soilbio.2008.03.020 |
| [21] | 檀满枝, 詹其厚, 陈杰. 基于信息熵原理的土壤pH影响因素空间相关性分析. 土壤, 2007, 39(6): 953–957. DOI:10.3321/j.issn:0253-9829.2007.06.022 |
| [22] | 魏春兰, 马红亮, 高人, 尹云锋, 陈仕东, 杨柳明. 模拟氮沉降对森林土壤可溶性有机碳的影响. 亚热带资源与环境学报, 2013, 8(4): 16–24. DOI:10.3969/j.issn.1673-7105.2013.04.003 |
| [23] | Silveira M L, Liu K S, Sollenberger L E, Follett R F, Vendramini J M B. Short-term effects of grazing intensity and nitrogen fertilization on soil organic carbon pools under perennial grass pastures in the southeastern USA. Soil Biology and Biochemistry, 2013, 58: 42–49. DOI:10.1016/j.soilbio.2012.11.003 |
| [24] | 周纪东, 史荣久, 赵峰, 韩斯琴, 张颖. 施氮频率和强度对内蒙古温带草原土壤pH及碳、氮、磷含量的影响. 应用生态学报, 2016, 27(8): 2467–2476. |
| [25] | Zhang N L, Wan S Q, Li L H, Bi J, Zhao M M, Ma K P. Impacts of urea N addition on soil microbial community in a semi-arid temperate steppe in northern China. Plant and Soil, 2008, 311(1/2): 19–28. |
| [26] | 汪金松, 赵秀海, 张春雨, 李化山, 王娜, 赵博. 模拟氮沉降对油松林土壤有机碳和全氮的影响. 北京林业大学学报, 2016, 38(10): 88–94. |
| [27] | Nadelhoffer K J, Emmett B A, Gundersen P, Kjnaas O J, Koopmans C J, Schleppi P, Tietema A, Wright R F. Nitrogen deposition makes a minor contribution to carbon sequestration in temperate forests. Nature, 1999, 398(6723): 145–148. DOI:10.1038/18205 |
| [28] | 李秋玲, 肖辉林, 曾晓舵, 冯乙晴, 莫江明. 模拟氮沉降对森林土壤化学性质的影响. 生态环境学报, 2013, 22(12): 1872–1878. DOI:10.3969/j.issn.1674-5906.2013.12.003 |
| [29] | Zhang Y H, Han X, He N P, Long M, Huang J H, Zhang G M, Wang Q B, Han X G. Increase in ammonia volatilization from soil in response to N deposition in Inner Mongolia grasslands. Atmospheric Environment, 2014, 84: 156–162. DOI:10.1016/j.atmosenv.2013.11.052 |
| [30] | 闫聪微, 马红亮, 高人, 尹云锋, 陈仕东. 模拟氮沉降对中亚热带森林土壤中可溶性氮含量的影响. 环境科学研究, 2012, 25(6): 678–684. |
| [31] | 元晓春, 陈岳民, 袁硕, 郑蔚, 司友涛, 元志鹏, 林伟盛, 杨玉盛. 氮沉降对杉木人工幼林土壤溶液可溶性有机物质浓度及光谱学特征的影响. 应用生态学报, 2017, 28(1): 1–11. |
| [32] | Ippolito J A, Blecker S W, Freeman C L, McCulley R L, Blair J M, Kelly E F. Phosphorus biogeochemistry across a precipitation gradient in grasslands of central North America. Journal of Arid Environments, 2010, 74(8): 954–961. DOI:10.1016/j.jaridenv.2010.01.003 |
| [33] | 陈美领, 陈浩, 毛庆功, 朱晓敏, 莫江明. 氮沉降对森林土壤磷循环的影响. 生态学报, 2016, 36(16): 4965–4976. |
| [34] | 张继舟, 倪红伟, 王建波, 王立民, 王宏韬. 氮沉降和CO2浓度增加对土壤磷素含量的影响. 中国农学通报, 2013, 29(17): 182–187. DOI:10.11924/j.issn.1000-6850.2013-0379 |
| [35] | 李银, 曾曙才, 黄文娟. 模拟氮沉降对鼎湖山森林土壤酸性磷酸单酯酶活性和有效磷含量的影响. 应用生态学报, 2011, 22(3): 631–636. |
| [36] | 周嘉聪, 刘小飞, 郑永, 纪宇皝, 李先锋, 徐鹏程, 陈岳民, 杨玉盛. 氮沉降对中亚热带米槠天然林微生物生物量及酶活性的影响. 生态学报, 2017, 37(1): 127–135. |
| [37] | 赵玉涛, 韩士杰, 李雪峰, 胡艳玲. 模拟氮沉降增加对土壤微生物量的影响. 东北林业大学学报, 2009, 37(1): 49–51. DOI:10.3969/j.issn.1000-5382.2009.01.018 |
| [38] | 周世兴, 邹秤, 肖永翔, 向元彬, 韩博涵, 唐剑东, 罗超, 黄从德. 模拟氮沉降对华西雨屏区天然常绿阔叶林土壤微生物生物量碳和氮的影响. 应用生态学报, 2017, 28(1): 12–18. |
| [39] | 郭依秋, 范秀华, 汪金松, 金冠一. 太岳山油松林土壤微生物量对模拟氮沉降的响应. 应用与环境生物学报, 2013, 19(4): 605–610. |
| [40] | Compton Jana EC, Watrud Lidia S W, Porteous L Arlene P, DeGrood S. Response of soil microbial biomass and community composition to chronic nitrogen additions at Harvard forest. Forest Ecology and Management, 2004, 196(1): 143–158. DOI:10.1016/j.foreco.2004.03.017 |
| [41] | 袁颖红, 樊后保, 李燕燕, 刘文飞, 廖迎春, 沈芳芳, 黄荣珍. 模拟氮沉降对土壤酸化和土壤盐基离子含量的影响. 应用与环境生物学报, 2011, 17(4): 461–466. |
| [42] | Manning P, Saunders M, Bardgett R D, Bonkowski M, Bradford M A, Ellis R J, Kandeler E, Marhan S, Tscherko D. Direct and indirect effects of nitrogen deposition on litter decomposition. Soil Biology and Biochemistry, 2008, 40(3): 688–698. DOI:10.1016/j.soilbio.2007.08.023 |
| [43] | 涂利华, 戴洪忠, 胡庭兴, 张健, 雒守华. 模拟氮沉降对华西雨屏区撑绿杂交竹林土壤呼吸的影响. 应用生态学报, 2011, 22(4): 829–836. |
| [44] | Högberg M N, Briones M J I, Keel S G, Metcalfe D B, Campbell C, Midwood A J, Thornton B, Hurry V, Linder S, Näsholm T, Högberg P. Quantification of effects of season and nitrogen supply on tree below -ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest. New Phytologist, 2010, 187(2): 485–493. DOI:10.1111/j.1469-8137.2010.03274.x |
| [45] | He H B, Zhang W, Zhang X D, Xie H T, Zhuang J Z. Temporal responses of soil microorganisms to substrate addition as indicated by amino sugar differentiation. Soil Biology and Biochemistry, 2011, 43(6): 1155–1161. DOI:10.1016/j.soilbio.2011.02.002 |
| [46] | DeForest J L, Zak D R, Pregitzer K S, Burton A J. Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest. Soil Biology and Biochemistry, 2004, 36(6): 965–971. DOI:10.1016/j.soilbio.2004.02.011 |
| [47] | 莫江明, 薛璟花, 方运霆. 鼎湖山主要森林植物凋落物分解及其对N沉降的响应. 生态学报, 2004, 24(7): 1413–1420. DOI:10.3321/j.issn:1000-0933.2004.07.015 |
| [48] | 邓小文, 张岩, 韩士杰, 李晓峰, 李考学, 李雪峰. 外源氮输入对长白山红松凋落物早期分解的影响. 北京林业大学学报, 2007, 29(6): 16–22. DOI:10.3321/j.issn:1000-1522.2007.06.003 |
| [49] | 郭萍萍, 郑丽丽, 黄幸然, 吴旺旺, 卓振华, 易志刚. 模拟大气氮沉降对不同树种土壤微生物生物量的影响. 生态环境学报, 2015, 24(5): 772–777. |
| [50] | Grayston S J, Prescott C E. Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Biology and Biochemistry, 2005, 37(6): 1157–1167. DOI:10.1016/j.soilbio.2004.11.014 |
| [51] | 宋蕾, 林尤伟, 金光泽. 模拟氮沉降对典型阔叶红松林土壤微生物生物量碳氮的影响. 南京林业大学学报:(自然科学版), 2017, 41(5): 7–12. |
| [52] | 刘爽, 王传宽. 五种温带森林土壤微生物生物量碳氮的时空格局. 生态学报, 2010, 30(12): 3135–3143. |
| [53] | 许可, 王春梅, 张艺, 杨欣桐, 刘卫敏. 模拟大气氮沉降对温带森林土壤微生物群落结构的影响. 生态学杂志, 2016, 35(10): 2676–2683. |