 2018, Vol. 38
2018, Vol. 38文章信息
- 陈冰冰, 孙志高, 孙文广, 王伟, 王苗苗, 胡星云.
- CHEN Bingbing, SUN Zhigao, SUN Wenguang, WANG Wei, WANG Miaomiao, HU Xingyun.
- 外源氮输入对黄河口新生湿地植物-土壤系统硫分布特征的影响
- Effects of exogenous nitrogen enrichment on the distribution of total sulfur in the plant-soil system of the newly created marshes in the Yellow River estuary, China
- 生态学报. 2018, 38(16): 5698-5708
- Acta Ecologica Sinica. 2018, 38(16): 5698-5708
- http://dx.doi.org/10.5846/stxb201707071233
-
文章历史
- 收稿日期: 2017-07-07
- 网络出版日期: 2018-05-11





2. 路易斯安那州立大学植物, 环境与土壤科学系, 巴吞鲁日 700803;
3. 鲁东大学地理与规划学院, 烟台 264025;
4. 成都信息工程学院 资源环境学院, 成都 610225
2. School of Plant, Environment and Soil Science, Louisiana State University, Baton Rouge 700803, USA;
3. Department of Geography and Planning, Ludong University, Yantai 26025, China;
4. Department of Resources and Environment, College of Information and Engineering, Chengdu 610225, China
河口湿地是河口与海洋相互作用形成的重要生态类型,是海岸带地区响应全球变化和人类活动较为敏感的生态系统之一[1]。硫作为继氮、磷、钾之后的第四位营养元素是植物生长必需的中量矿质营养元素,在许多生物地球化学过程中扮演着重要角色,如参与蛋白质、氨基酸和叶绿素形成,控制光合作用中碳水化合物代谢、影响植物呼吸作用和抗逆性等[2],其对于维持河口湿地生态系统健康具有重要意义。受河口水动力和海洋潮汐的双重影响,河口湿地的硫酸盐含量丰富且硫酸盐的动态平衡及其还原产物形成了河口湿地独特的硫循环体系[3],其与碳矿化、CH4释放、水体酸化、黄铁矿形成、金属元素循环以及大气硫释放等一系列生物地球化学过程密切相关[4-6]。目前,国外关于湿地硫循环的研究已涉及土壤-水体系硫形态分布、有机硫矿化特征、SO42-异化还原及其与C、N、Fe等耦合过程、含硫气体释放以及植物硫累积与残体分解释放[7]等方面。与国外相比,国内在这方面也开展了许多工作,且现有研究已涉及盐沼[8-9]、淡水沼泽[10-11]、泥炭湿地[12]、红树林沼泽[13]和河口湿地[14-18],而关于河口湿地硫循环的相关研究主要集中于黄河口湿地[14-15]、长江口湿地[16]、闽江口湿地[17-18]以及南方主要河口(如九龙江口、漳江口)红树林沼泽[13]等。整体而言,当前关于河口湿地硫循环的研究主要集中在某一过程,而对硫元素与碳、氮、Fe等元素的交互作用研究还比较缺乏。
黄河口湿地是渤海与黄河河口相互作用形成的中国暖温带最广阔、最完整和最年轻的滨海湿地,承接上游带来的大量含氮物质。近年来,黄河口的营养盐入海通量已达1.41—4.22×104 t,并始终保持较高水平[19]。另外,该区的氮湿沉降量已从1980年的1—2 g m-2a-1增至当前的3—4.5 g m-2a-1[20],接近其氮沉降临界负荷(4.0 g m-2a-1)[21],而这均可能对湿地植物生长与养分循环产生深刻影响[22]。碱蓬(Suaeda salsa)作为黄河口新生湿地的典型盐生植被,是黄河三角洲丹顶鹤、白鹭、鹆形目鸟类等的主要栖息地,同时还具有维持湿地系统正常演替、防风固堤和调节气候等多重重要功能[23]。当前,许多学者已在黄河口碱蓬湿地硫元素分布特征[14]、硫累积与残体释放特征[24-25]、含硫气体释放[26]以及硫生物循环评估[27]等方面开展了许多工作,但这些研究多是基于野外原位采样来实施,真正关于野外条件下持续氮输入对碱蓬湿地植被-土壤系统硫生物循环影响的模拟试验研究还不多见。已有研究表明,氮与硫在湿地中存在着复杂的耦合关系,外源氮输入增加不但可影响植物对氮硫的吸收利用的状况[28-29]、改变土壤硫转化过程[30]、促进含硫气体(如COS、CS2)的释放[31],而且还可影响硫驱动下的自养硝态氮还原菌参与的自养反硝化作用[32],导致N2O和N2释放量的显著增加[33]和含硫气体释放规律的改变[31]。然而,当前关于外源氮输入条件下河口湿地氮与硫的耦合机制研究还鲜有报道。鉴于此,本研究以黄河口北部滨岸高潮滩的典型碱蓬湿地为研究对象,基于野外原位氮输入模拟试验,研究了不同氮输入梯度下碱蓬湿地土壤硫分布特征以及植被硫累积与分配特征的差异,并从氮与硫养分供给角度明确了二者的耦合关系。研究结果有助于揭示黄河口湿地硫生物循环对外源氮输入的响应机制,并可为该区退化湿地的恢复与重建提供重要依据。
1 研究区域与研究方法 1.1 研究区概况研究区位于山东黄河三角洲国家级自然保护区(37°40′ N—38°10′ N, 118°41′ E—119°16′ E),该保护区于1992年经国家林业局批准建立,是中国最大的河口三角洲自然保护区,在世界范围河口湿地生态系统中具有较强代表性,主要保护新生湿地生态系统和珍稀濒危鸟类。保护区总面积为15.3万hm2,其中缓冲区1.3万hm2,实验区8.2万hm2。研究区属暖温带季风气候区,具有明显的大陆性季风气候特点,四季分明,雨热同期,冷热干湿界限明显,年降水量551.6 mm,70%的降水集中在7—8月,年均温12.1℃,无霜期196 d,≥10℃积温约4300℃,年蒸发量1928.2 mm。该区的土壤类型主要为隐域性潮土和盐土,主要植被类型为芦苇(Phragmites australis)、碱蓬(S. salsa)、柽柳(Tamatix chinensis)、白茅(Imperata cylindrica)及罗布麻(Apocynum venetum)等,其中芦苇、碱蓬和柽柳的分布较广。
1.2 研究方法 1.2.1 实验样地布设试验选择今黄河入海口北部滨岸高潮滩的碱蓬湿地为研究对象,参照欧洲NITREX项目以及靳英华等在长白山地区类似研究的设计[34],开展外源氮野外原位输入模拟试验。在碱蓬湿地试验区内布设相关装置通过人工喷氮方式模拟外源氮输入。试验设4个处理,即N0[对照处理,无额外氮输入,其值为当前湿地实际氮输入量。结合该区现有资料,考虑陆源氮输入(2.5—3.5 gN m-2 a-1)和氮沉降(3—4.5 gN m-2 a-1)的综合影响,将值确定为6.0 gN m-2 a-1]、N1[低氮处理:1.5N0(9.0 gN m-2 a-1),模拟湿地未来较低的外源氮增加量]、N2[中氮处理:2.0N0(12.0 gN m-2 a-1),模拟湿地未来较高的外源氮增加量]和N3[高氮处理:3.0N0(18.0 gN m-2 a-1),模拟湿地未来更高的外源氮增加量],每个处理设3个重复样地(5 m×10 m)。试验于植物生长季(4—11月)进行,自4月下旬开始,每隔20—30 d以CO(NH2)2水溶液的形式对不同样地进行外源氮输入强度的模拟,共喷氮8次。按照4种处理在不同阶段的氮输入要求,将CO(NH2)2溶解在20 L水中,通过预布设装置均匀喷洒入各样地,对照样地喷洒等量的水以减少因外加水造成对湿地生态过程的影响。
1.2.2 样品的采集与测定用土壤采样器在上述不同样地内分别逐月采集3个土壤柱样,柱样深度为40 cm(10 cm一层)。将采集的新鲜土样及时带回实验室后自然风干,去除杂物研磨过100目筛后装袋待测。地上生物量采用收获法,自5月份按照植物生长特点每20—30 d采样1次。在不同样地内选取3—4个25 cm×25 cm的样方,沿地面剪下植物地上部分,带回实验室将其分离为茎、叶和立枯物。由于成熟期碱蓬的果实与小叶紧密相连,难以准确区分,所以实际为叶+果实。地下生物量采用挖掘法,并在地上生物量测定样方内进行测定。采样时,将样方内0—40 cm的根挖出,放在细纱网袋中将泥土冲洗干净至无。枯落物采样与生物量测定同步,将样方内的植物地上部分割除,摘除立枯物(实验室进行)。将采集的地上、地下植物及立落物样品置于80℃烘干箱中烘干至恒重。称量后将样品粉碎,过0.25 mm筛后备用。土壤和植物样品的全硫(TS)含量采用Vario EL型元素分析仪测定。
1.2.3 指标计算不同氮输入处理下植物不同器官的S累积系数AF(Accumulation factor)可用不同器官的TS含量与土壤TS含量(0—20 cm均值)比值来表征[35],计算公式如下:
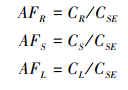
|
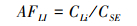
|
式中,CR、CS、CL、CLI分别为植物根、茎、叶和枯落物中S平均含量,CSE表示土壤中S平均含量。
植物体S的储量Hn(mg/m2)可用下式计算:
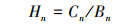
|
式中,Cn为植物体中的S含量(mg/g),Bn表示植物体的生物量(g/m2)。
土壤剖面第i层的S库储量(LSi)为相应土层S含量(Si)、土壤容重(dvi)与土壤厚度(hi)的乘积[36],即
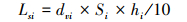
|
单位面积一定剖面深度(j到n层)的S库储量(LS)为j到n层储量之和,即
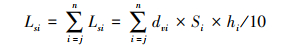
|
式中,LSi 、LS的单位为g/m2,dvi的单位为g/cm3,Si的单位为mg/g,hi的单位为cm。
1.2.4 数据处理与统计运用Origin 8软件对数据进行作图和计算,采用SPSS 23.0软件对不同氮输入处理间的土壤TS含量和植物不同部分的TS含量数据进行方差分析,显著性水平设定为P=0.05。
2 结果与分析 2.1 N输入对土壤TS含量分布的影响尽管不同氮输入处理下碱蓬湿地不同土层的TS含量均具有相似的季节变化模式,但氮输入明显改变了湿地土壤的TS含量分布状况(图 1)。随着N输入量的增加,除0—10 cm土层的TS含量增加不明显外,其他土层的TS含量均呈增加趋势,且这种趋势在20—30 cm土层表现尤为明显。相对于N0处理,N3处理下的TS平均含量增加了17.91%。在垂直方向上,除N3处理下的TS含量在10—20 cm土层最高外,其他处理下的TS含量均于0—10 cm土层取得最高值。方差分析表明,尽管N1、N2和N3处理下0—10 cm或30—40 cm土层中的TS含量与N0处理下相应土层中TS含量均存在极显著差异(P<0.01),但三者相应土层TS含量之间的差异并不显著(P>0.05)。同时,仅N3处理下10—20 cm土层的TS含量较N0处理存在显著差异(P<0.05),而3种处理下20—30 cm土层的TS含量均较N0处理存在极显著差异(P<0.01)。另外,N3与N1处理下20—30 cm土层的TS含量亦存在显著差异(P<0.05)。

|
| 图 1 不同氮输入处理下不同土层中全硫(TS)含量季节变化 Fig. 1 Seasonal variations ofthe total sulfur (TS) content in different soil layers as affected by different nitrogen import treatments N0:无氮输入处理, no N import treatment; N1:低氮输入处理, low N import treatment; N2:中氮输入处理, middle N import treatment; N3:高氮输入处理, high N import treatment |
不同氮输入处理下碱蓬各器官的TS含量变化特征存在显著差异(P<0.05)(图 2),其在整体上均表现为叶>茎>根,说明叶是硫的主要累积器官。生长初期,除N3处理下根的TS含量呈小幅增加外,其他处理下根的TS含量呈波动降低趋势,茎和叶的TS含量则呈增加趋势;生长旺期,不同处理下根和茎的TS含量总体均有不同程度降低,而叶的TS含量整体呈不规则“M”型变化,并均于6月末达到最高值;生长末期,不同处理下根和茎的TS含量总体呈增加趋势,且二者分别于N2和N3处理下的增幅最为明显。与之不同,此间叶的TS含量除在N3处理下增加明显外,其他处理下的TS含量均出现不同程度的降低,且均与N3处理存在极显著差异(P<0.01)。就立枯体而言,其TS含量在不同处理下的变化在9月上旬前与叶存在一定的相似性(图 2),即叶的TS含量较高时,立枯体的TS含量也较高。方差分析表明,5—10月不同部分的TS含量在各处理间的差异均不显著(P>0.05),但在10—11月,N3处理下叶和立枯体的TS含量均明显高于其他处理(P<0.05)。就生长季不同部分TS含量的平均状况而言(图 3),根在生长季的TS平均含量在N2处理下最低,而在N1和N3处理下均较高;茎、叶和立枯体的TS平均含量在N1和N2处理下均较低,而在N3处理下均较高。

|
| 图 2 不同氮输入处理下碱蓬不同部分TS含量季节变化 Fig. 2 Seasonal variations of TS content in different parts of Suaeda salsa as affected by different nitrogen import treatments |

|
| 图 3 不同氮输入处理下碱蓬不同部分TS平均含量及其变异系数 Fig. 3 Means and coefficient of variation of TS content in different organs of Suaeda salsa by different nitrogen import treatments CV:变异系数, coefficient of variation |
尽管不同氮处理下碱蓬各部分的S分配比在不同时期均存在明显差异(图 4),但各部分S分配比整体以叶最高,根最低,说明叶是主要的S累积器官。生长初期,根的S分配比除在N3处理下出现小幅增加外,其他处理下均呈明显降低趋势;生长旺期,不同处理下根的S分配比均呈小幅变化,且均于9月上旬取得最低值;生长末期,根的S分配比除在N3处理下出现小幅降低外,其他处理下均呈增加趋势,且以N2处理下的增幅最为明显。不同氮处理下茎和叶的S分配比在5—10月整体均呈先降低后增加变化,并均于生长末期骤然降低,而立枯体的S分配比在此间均骤然增加。比较而言,不同处理下各部分的S分配比在5—6月整体均表现为叶>茎>根,7—10月表现为叶>茎>枯>根;11月下旬,N0、N1、N3处理下各部分的S分配比均表现为枯>茎>叶>根,N2处理下则表现为枯>茎>根>叶。

|
| 图 4 不同氮输入处理下碱蓬不同部分硫分配比季节变化 Fig. 4 Seasonal variations of sulfur allocation proportions of different parts of Suaeda salsa as affected by different nitrogen import treatments |
尽管不同氮处理下碱蓬各部分的S累积系数(AF)存在较大差异,但在生长季整体均表现为叶>枯>茎>根,这与前述植物不同部分TS含量的变化特征相一致。就平均状况而言,随着氮输入量的增加,根、茎和叶的AF值均于N2处理下出现明显降低,而在其他处理下变化不大。与之不同,立枯体的AF值以N0处理最高,N3处理次之,而N1和N2处理较低且比较相近(图 5)。不同氮处理下植物-土壤系统硫分配的研究表明,植物地上器官是植物亚系统的主要S库(表 1),且其AF值整体随氮输入量的提高而增加。在不同氮处理下的植物亚系统中,根的平均S储量最低,叶最高,茎次之。同时,土壤S储量整体也随氮输入量的提高而增加,且其分配比明显高于植物亚系统。就植物-土壤系统而言,其S储量亦随氮输入量的提高而增加。相较N0处理,N1、N2、N3处理下植物亚系统S储量的增幅分别为18.03%、20.81%和59.18%,而土壤S储量的增幅分别为0.33%、5.62%和8.18%,均远低于氮输入的增幅(50%、100%和200%)。

|
| 图 5 不同氮输入处理下碱蓬各部分的硫累积系数 Fig. 5 Accumulation factors of sulfur in different parts of Suaeda salsa as affected by different nitrogen import treatments AF:硫累积系数, Accumulation factors of sulfur |
| 项目 Item |
根 Root |
茎 Stem |
叶 Leaf |
立枯体 Litter |
植物亚系统 Plantsubsystem |
土壤(0—40 cm) Soil (0—40 cm) |
植物-土壤体系 Plant-soil system |
| N0/(g/m2) | 0.096±0.03 | 0.995±0.38 | 1.573±0.90 | 0.716±0.70 | 3.380±0.75 | 6. 29 | 9.67 |
| 累积系数CV | 0.33 | 0.38 | 0.57 | 0.98 | 0.22 | — | — |
| 分配比/% Allocation proportion | 2.8a | 29.4a | 46.5a | 21.2a | 34.9b | 65.1b | 100.0b |
| N1/(g/m2) | 0.115±0.05 | 1.252±0.81 | 1.857±1.22 | 0.767±0.88 | 3.990±1.10 | 6.31 | 10.30 |
| 累积系数CV | 0.46 | 0.65 | 0.66 | 1.14 | 0.27 | — | — |
| 分配比/% Allocation proportion | 2.9a | 31.4a | 46.5a | 19.2a | 38.7b | 61.3b | 100.0b |
| N2/(g/m2) | 0.118±0.04 | 1.305±1.11 | 1.862±1.09 | 0.798±0.99 | 4.084±1.20 | 6.65 | 10.73 |
| 累积系数CV | 0.37 | 0.85 | 0.59 | 1.24 | 0.29 | — | — |
| 分配比/% Allocation proportion | 2.9a | 32.0a | 45.6a | 19.5a | 38.1b | 61.9b | 100.0b |
| N3/(g/m2) | 0.168±0.13 | 1.686±0.58 | 2.421±1.32 | 1.106±1.18 | 5.381±1.20 | 6.81 | 12.19 |
| 累积系数CV/% | 0.79 | 0.35 | 0.54 | 1.07 | 0.22 | — | — |
| Allocation proportion | 3.1a | 31.3a | 45.0a | 20.5a | 44.1b | 55.9b | 100.00b |
| a占植物亚系统中的S储量比例; b占植物-土壤体系中的S储量比例。N0:无氮输入处理, no N import treatment; N1:低氮输入处理, low N import treatment; N2:中氮输入处理, middle N import treatment; N3:高氮输入处理, high N import treatment | |||||||
本研究表明,不同氮输入处理下植物各器官TS含量的季节变化存在一定差异。这一方面可能与植物相应器官的结构与功能有关,另一方面可能与不同时期硫的营养功能差异有关。生长初期,除N3处理下根的TS含量呈小幅增加外,其他处理下根的TS含量总体呈波动降低趋势,而茎和叶的TS含量呈增加趋势。这主要是由于根在生长初期需输送大量硫养分以供给地上器官,特别是流向正在发育的叶[37],而茎作为营养传输器官其含量亦随之增加。生长旺期,不同氮处理下根中TS含量的降低(图 2)主要与硫的生理功能有关,即碱蓬叶的生长发育及光合生产需大量S来构成叶绿体,而根作为硫的重要输出库必须在植物生长高峰期将大量硫营养转移到地上器官,由此导致其TS含量明显降低[14]。生长末期,植物开始大量死亡,此时地上生物器官中的硫开始向地下转移,由此导致不同处理下根和茎中的TS含量总体呈增加趋势。例如,N2处理下叶中TS含量的降幅最为明显,而其根中TS含量则骤然增加(图 2)。由于茎是营养元素的传输器官,其对S元素整体起到传输作用,故不同氮处理下茎中的TS含量变化不大。与根和茎不同,不同氮处理下叶的TS含量在不同时期呈不同变化特征,其在生长初期及旺期整体表现为N0>N1>N2>N3,而在生长末期表现为N3>N1>N0>N2。生长初期及旺期叶中TS含量随氮输入增加而降低的原因可能与大量氮输入导致植物硫的生理性缺乏有关。McGrath等(1996)以及Rsmanussen和Krsge(1986)的研究也发现,高氮会诱导植物生理性缺硫,使得植物中的硫含量较低[38-39]。另外,不同氮处理下叶中的TS含量均在8月中旬出现较低值,原因可能在于8月中旬是植物生长高峰期,此间叶的生物量迅速增加导致的硫“稀释效应”明显有关(图 6)。9—10月是碱蓬果实(种子)发育和成熟的时期,此时不仅根系需要向其供给大量氮硫营养,而且茎和叶中的氮硫营养也会大量转移至果实中,故此时叶(实际为叶+果实)中的TS含量又迅速增加。生长末期,叶的TS含量在N3处理下增加明显,原因可能与N3处理下碱蓬的生长发育期较其他氮处理延缓有关。已有研究发现,高氮处理不仅促进了黄河口碱蓬地上生物量的增加,而且还使得碱蓬生长高峰期相对于无氮处理推迟20 d左右[40]。Sun等的研究亦发现,氮添加可使三江平原小叶章(Calamagrostis angustifolia)茎和叶的最大生物量相对于无氮处理取得的时间推迟了15—30 d[41]。本研究中,生长末期N2处理下叶中TS含量的显著降低可能与该处理下碱蓬的种子发育可能提前有关。胡星云等对该处理下碱蓬的相关研究表明,碱蓬的种子发育相对于其他处理可能会提前约1个月[40]。正是如此,该处理下的植被在生长末期可能提前枯死,而其地上器官中的硫营养亦极可能提前向地下转移,由此在一定程度上增加了N2处理根中的TS含量,并同时明显降低了其叶中的TS含量。

|
| 图 6 不同氮输入处理下碱蓬生物量季节动态 Fig. 6 Seasonal dynamics of the biomasses of different organs of Suaeda salsa as affected by different nitrogen import treatments |
研究还发现,不同氮处理下立枯体中的TS含量变化在9月上旬前与叶存在一致性(图 2),即当叶中的TS含量较高时,立枯体的TS含量也较高。原因在于,9月上旬前产生的立枯体主要为叶,而叶的TS含量在此期间又相对较高,由此使得叶在枯死前虽存在部分硫转移,但滞留在叶枯体中的硫含量依然较高。9月中旬至10月上旬产生的立枯体主要为茎和叶,而茎和叶中的硫营养又逐渐转移至碱蓬的果实中,由此导致此间立枯体中的TS含量出现明显降低。10月中旬立枯体中TS含量(特别N3处理)的骤然增加可能主要与生长末期碱蓬种子发育成熟后未完全脱落以及茎、叶大量枯死后的硫滞留有关。比较而言,生长末期N3处理下立枯体的TS含量整体高于N0处理,充足的氮养分供给可能使得N3处理下碱蓬地上器官的硫累积增强(图 5),“稀释效应”不明显,进而导致该处理下产生立枯体的TS含量也较高。与之不同,N1和N2处理下立枯体的TS含量整体均低于N0处理。原因在于,碱蓬在N1和N2处理下对硫养分的吸收较N0多,但由于二者的地上生物量远大于N0处理(图 6),“稀释效应”明显,故导致两种处理下产生立枯体的TS含量均低于N0处理。
本研究还表明,尽管氮输入处理并未改变碱蓬的硫分配格局以及其地上与地下之间的硫养分供给关系,但可能改变了植被的硫养分吸收与利用策略,而这一策略在N2处理下表现的尤为明显。本研究中,根的S分配比除在生长末期N3处理下出现小幅降低外,其他处理下均呈增加趋势,且以N2处理下的增幅最为明显。原因可能与植物体为适应不同养分环境而进行的自身生长特性及养分分配的调整有关,而这已为我们前期的研究所证实,即生长末期N2处理下碱蓬的地上生物量与N1处理相当,但其地下生物量则与N3处理相当,且N2处理下植物地上器官中的硫养分极有可能提前向地下转移[26],由此导致其根中的S分配比较高,而叶的分配比较低。实际上,相关研究也得到类似结论。如Xie等的研究表明,养分的输入刺激了水葫芦(Eichhornia crassipes)的生长,但当其生长环境中的养分含量增加后,植物体自身会适时调整生物量的分配以达到最获得资源的最佳利用[42]。
3.2 N输入对土壤-植物系统S分配特征的影响本研究表明,N0、N1和N2处理下表层土壤的TS含量最高,虽然N3处理下10—20 cm土层的TS含量最高(1.33 mg/g),但其在0—10 cm的含量也不低(1.30 mg/g)。表层土壤中的较高硫含量主要与大量输入至表层土壤中的氮促进了地表残体分解及硫养分归还有关。Akira等对日本Sasakami泥炭地有机物质分解的研究表明,有机物质的失重率与泥炭中的N、P浓度成正比[43]。Steven的研究也发现,Cladium jamaicense和Typha domingensis枯落物在贫养分条件下分解最慢,而在适度养分条件下分解最快[44]。研究还表明,随着氮输入量的增加,除表层土壤的TS含量增加不明显外,其他土层的TS含量均呈增加趋势。前述可知,外源氮输入可促进地表残体分解及硫养分归还,使得不同氮处理下表层土壤的硫含量均较高。由于碱蓬根系主要集中于表层土壤(0—10 cm),而根系从表层土壤中吸收的硫养分不断输送至地上以促进地上器官的发育,故不同处理下表层土壤中的TS含量变化往往受制于植物生长节律和地上生物量变化的影响。随着氮输入量的增加,土壤中的硫含量也随之增高,但因植物生物量也随之增大(图 6),故对S的吸收量相对就越多,土壤S的降低也较大,从而使得不同处理下表层土壤中S的增加并不明显。相比之下,根系很少从其他土层吸收硫养分,而且可能由于外源氮输入至较深土层而对有机物的分解及硫养分归还产生促进作用有关,由此导致较深土层中的TS含量增加明显。较深土层中TS含量的增加还可能与潮滩较强沉积作用对植物残体的掩埋以及无脊椎动物的活动有关。受调水调沙工程和潮汐扰动沉积的共同影响,黄河口碱蓬盐沼的全年沉积可增加约90—100 mm/a[45],较强的沉积可掩埋植物残体。当外源氮输入至较深土层,有利于沉积掩埋下植物残体的分解,从而使得较深土层中TS含量增加明显。另外,由于螃蟹[日本大眼蟹(Macrophthalmus japonicus)和伍氏厚蟹(Helice wuana)为优势种]为黄河口碱蓬盐沼最为典型的小型无脊椎动物,其洞穴密度比较大,垂直深度为15—20 cm,并在20 cm深度左右建造水平或倾斜栖息洞穴[46]。一方面,螃蟹觅食可能会把表层一部分植物残体带到洞穴中,使得较深土层植物残体增加;另一方面,其分泌物也会增加土壤中的有机物。当外源氮输入至该土层,这部分残体和分泌物会增加该层土壤中的养分,进而使得较深土壤中的TS含量较高。
本研究发现,生长季碱蓬各部分的S累积系数在不同氮处理下整体均表现为叶>枯>茎>根,这与叶是硫的主要累积器官有关。另外,随着N输入量的增加,植物亚系统的S储量也随之增加,说明N输入有利于植物对S的累积。然而,随着N输入量的增加,土壤S储量的增幅远低于植物亚系统S储量的增幅以及氮供给的增幅,即尽管土壤中的S养分呈增加趋势,但在N供给大幅增加的前提下,N、S之间的养分供给存在不同步性。当N输入量很高时,土壤中的S养分增幅却不大,而此间植物对S养分的需求量又很高,说明如果没有外源S养分及时补充的前提下土壤的供S能力远不能满足植被对硫养分的大量需求,而外源硫养分的补给主要有两个途径:一是与海水中的硫酸盐含量丰富[3],其随潮汐不断补给碱蓬湿地系统有关;二是与上述提及的氮输入增加对地表残体分解及硫养分归还的促进作用有关。不管是何种S补给途径,不同处理下上述植物—土壤系统所表现出的N、S养分供给不同步的现象表明,随着氮输入量的增加,碱蓬湿地系统的S生物循环速率可能会加速。近年来,黄河口的营养盐入海通量尽管年际变化较大,但一直保持在较高水平[19],这就使得黄河口新生湿地的养分状况可能会不断被改善。在未来黄河口N养分负荷增加情况下,碱蓬湿地植物-土壤系统的硫生物循环速率不但可能会加速,而且N、S养分之间也可能形成一个正反馈机制。由于硫养分供给对植物叶绿体形成和光合生产具有重要影响[47],故从硫养分供给角度,硫生物循环加速将有利于维持新生湿地的稳定与健康。
4 结论(1) 随着N输入量的增加,除表层TS含量变化不明显外,其他土层均呈增加趋势。不同氮处理下植物各器官的TS含量整体表现为叶>茎>根,说明叶是硫的主要累积器官;
(2) 尽管氮输入处理并未改变碱蓬的硫分配格局以及其地上与地下之间的硫养分供给关系,但其为适应不同养分环境可进行自身生长特性及养分分配的调整;
(3) 随着N输入量的增加,土壤S储量的增幅远低于植物亚系统S储量的增幅以及氮供给的增幅,说明N、S之间的养分供给存在不同步性;
(4) 未来黄河口N养分负荷增加的情况下,N、S养分之间可能形成一个正反馈机制,而这将有利于维持新生湿地的稳定与健康。
| [1] | Simas T, Nunes J P, Ferreira J G. Effects of global climate change on coastal salt marshes. Ecological Modelling, 2001, 139(1): 1–15. DOI:10.1016/S0304-3800(01)00226-5 |
| [2] | 陆景陵. 植物营养学(上).2版. 北京: 中国农业大学出版社, 2003: 72–76. |
| [3] | 幸颖, 刘常宏, 安树青. 海岸盐沼湿地土壤硫循环中的微生物及其作用. 生态学杂志, 2007, 26(4): 577–581. DOI:10.3321/j.issn:1000-4890.2007.04.023 |
| [4] | Nedwell D B, Watson A. CH4 production, oxidation and emission in a U. K. ombrotrophic peat bog:influence of SO42- from acid rain. Soil Biology and Biochemistry, 1995, 27(7): 893–903. DOI:10.1016/0038-0717(95)00018-A |
| [5] | Mandernack K W, Lynch L, Krouse H R, Morgan M D. Sulfur cycling in wetland peat of the New Jersey Pinelands and its effect on stream water chemistry. Geochimica et Cosmochimica Acta, 2000, 64(23): 3949–3964. DOI:10.1016/S0016-7037(00)00491-9 |
| [6] | Roberta A, Gianmarco G, Marco B, David T W, Pierluigi V. Iron, sulphur and phosphorus cycling in the rhizosphere sediments of a eutrophic Ruppia cirrhosa meadow (Valle Smarlacca, Italy). Journal of Sea Research, 2001, 45(1): 15–26. DOI:10.1016/S1385-1101(00)00056-3 |
| [7] | Lamers L P M, Govers L L, Janssen I C J M, Geurts J J M, Van der Welle M E W, Van Katwijk M M, Van der Heide T, Roelofs J G M, Smolders A J P. Sulfide as a soil phytotoxin-a review. Frontiers in Plant Science, 2013, 4(9): 268–268. |
| [8] | Bo T, Fossing H, Bo B J. Manganese, iron and sulfur cycling in a coastal marine sediment, Aarhus bay, Denmark. Geochimica et Cosmochimica Acta, 1994, 58(23): 5115–5129. DOI:10.1016/0016-7037(94)90298-4 |
| [9] | Zhou J L, Wu Y, Kang Q S, Zhang J. Spatial variations of carbon, nitrogen, phosphorous and sulfur in the salt marsh sediments of the Yangtze Estuary in China. Estuarine, Coastal and Shelf Science, 2007, 71(1/2): 47–59. |
| [10] | Spratt H G J, Morgan M D. Sulfur cycling in a cedar-dominated, freshwater wetland. Limnology and Oceanography, 1990, 35(7): 1586–1593. DOI:10.4319/lo.1990.35.7.1586 |
| [11] | Küsel K, Roth U, Trinkwalter T, Peiffer S. Effect of pH on the anaerobic microbial cycling of sulfur in mining-impacted freshwater lake sediments. Environmental and Experimental Botany, 2001, 46(3): 213–223. DOI:10.1016/S0098-8472(01)00103-4 |
| [12] | Wieder R K, Lang G E. Cycling of inorganic and organic sulfur in peat from big run bog, West Virginia. Biogeochemistry, 1988, 5(2): 221–242. DOI:10.1007/BF02180229 |
| [13] | 林慧娜, 傅娇艳, 吴浩, 刘金玲, 李柳强, 虎贞贞, 丁振华. 中国主要红树林湿地沉积物中硫的分布特征及影响因素. 海洋科学, 2009, 33(12): 79–82. |
| [14] | 孙万龙, 孙志高, 林光辉, 牟晓杰, 王玲玲. 黄河口滨岸潮滩不同生境下翅碱蓬硫元素的季节变化. 植物生态学报, 2011, 35(3): 303–310. |
| [15] | 李新华, 孙志高, 孙文广, 朱振林. 黄河三角洲潮滩湿地系统二甲基硫排放通量的时空变化. 环境科学学报, 2015, 35(12): 3947–3955. |
| [16] | 曹爱丽. 长江口滨海沉积物中无机硫的形态特征及其环境意义[D]. 上海: 复旦大学, 2010. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y2225512 |
| [17] | 曾从盛, 张林海, 王天鹅, 张文娟, 仝川. 闽江河口湿地植物枯落物立枯和倒伏分解主要元素动态. 生态学报, 2012, 32(20): 6289–6299. |
| [18] | 胡敏杰, 邹芳芳, 任鹏, 杜威宁, 仝川. 河口潮滩湿地CH4、CO2排放通量对氮硫负荷增强的响应. 环境科学学报, 2016, 36(4): 1359–1368. |
| [19] | Sun Z G, Sun W G, Tong C, Zeng C S, Yu X, Mou X J. China's coastal wetlands:Conservation history, implementation efforts, existing issues and strategies for future improvement. Environment International, 2015, 79: 25–41. DOI:10.1016/j.envint.2015.02.017 |
| [20] | 遆超普, 颜晓元. 基于氮排放数据的中国大陆大气氮素湿沉降量估算. 农业环境科学学报, 2010, 29(8): 1606–1611. |
| [21] | 段雷, 郝吉明, 谢绍东, 周中平. 用稳态法确定中国土壤的硫沉降和氮沉降临界负荷. 环境科学, 2002, 23(2): 7–12. |
| [22] | Aber J D, McDowell W, Nadelhoffer K, Magill A, Berntson G, Kamakea M, McNulty S, Currie W, Rustad L, Fernandez I. Nitrogen saturation in temperate forest ecosystems:Hypotheses revisited. BioScience, 1998, 48(11): 921–934. DOI:10.2307/1313296 |
| [23] | Bauer G A, Bazzaz F A, Minocha R, Long S, Magill A, Aber J D, Berntson G M. Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (Pinus resinosa Ait.) stand in the NE United States. Forest Ecology and Management, 2004, 196(1): 173–186. DOI:10.1016/j.foreco.2004.03.032 |
| [24] | 郝庆菊, 王起超, 王其存, 李志博. 三江平原典型湿地及其开垦后土壤中总硫变化的初步研究. 应用生态学报, 2003, 14(12): 2191–2194. DOI:10.3321/j.issn:1001-9332.2003.12.022 |
| [25] | 李新华. 三江平原小叶章湿地系统硫的形态分布和转化过程研究[D]. 长春: 中国科学院东北地理与农业生态研究所, 2007. http://www.irgrid.ac.cn/handle/1471x/504562 |
| [26] | 李新华, 刘景双, 王金达, 孙志高. 陆地生态系统含硫气体释放研究进展. 生态环境学报, 2005, 14(1): 117–120. DOI:10.3969/j.issn.1674-5906.2005.01.025 |
| [27] | Sun Z G, Mou X J, Song H L, Jiang H H. Sulfur biological cycle of the different Suaeda salsa marshes in the intertidal zone of the Yellow River estuary, China. Ecological Engineering, 2013, 53(2): 153–164. |
| [28] | Tretiach M, Baruffo L. Effects of H2S on CO2 gas exchanges and growth rates of the epiphytic lichen Parmelia sulcata Taylor. Symbiosis. Symbiosis, 2001, 31(1): 35–46. |
| [29] | Welle M E W V D, Cuppens M, Lamers L P M, Roelofs J G M. Detoxifying toxicants:Interactions between sulfide and iron toxicity in freshwater wetlands. Environmental Toxicology and Chemistry, 2006, 25(6): 1592–1597. DOI:10.1897/05-283R.1 |
| [30] | Ghani A, Mclaren R G, Swift R S. Sulphur mineralisation and transformations in soils as influenced by additions of carbon, nitrogen and sulphur. Soil Biology and Biochemistry, 1992, 24(4): 331–341. DOI:10.1016/0038-0717(92)90193-2 |
| [31] | Kanda K, Tsuruta H, Minami K. Emission of dimethyl sulfide, carbonyl sulfide, and carbon bisulfide from paddy fields. Soil Science and Plant Nutrition, 1992, 38(4): 709–716. DOI:10.1080/00380768.1992.10416701 |
| [32] | Wu S B, Kuschk P, Wiessner A, Müller J, Saad R A B, Dong R J. Sulphur transformations in constructed wetlands for wastewater treatment:A review. Ecological Engineering, 2013, 52(2): 278–289. |
| [33] | Shao M F, Zhang T, Fang H H P, Li X D. The effect of nitrate concentration on sulfide-driven autotrophic denitrification in marine sediment. Chemosphere, 2011, 83(1): 1–6. DOI:10.1016/j.chemosphere.2011.01.042 |
| [34] | 靳英华, 许嘉巍, 宗盛伟, 王鹏. 氮沉降对长白山苔原植被影响的试验研究. 地理科学, 2014, 34(12): 1526–1532. |
| [35] | Duman F, Cicek M, Sezen G. Seasonal changes of metal accumulation and distribution in Common club rush(Schoenoplectus lacustris) and Common reed(Phragmites australis). Ecotoxicology, 2007, 16(6): 457–463. DOI:10.1007/s10646-007-0150-4 |
| [36] | 李新华, 刘景双, 孙志高, 杨继松. 三江平原小叶章湿地生态系统硫的生物地球化学循环. 生态学报, 2007, 27(6): 2199–2207. DOI:10.3321/j.issn:1000-0933.2007.06.007 |
| [37] | Sunarpi, Anderson J W. Distribution and Redistribution of Sulfur Supplied as[35S]Sulfate to Roots during Vegetative Growth of Soybean. Plant Physiology, 1996, 110(4): 1151–1157. DOI:10.1104/pp.110.4.1151 |
| [38] | Mcgrath S P, Zhao F J, Withers P J A. Development of sulphur deficiency in crops and its treatment//Proceedings of the Fertiliser Society, No. 379. Peterborough, UK: The Fertilisers Society, 1996. http://agris.fao.org/openagris/search.do?recordID=GB9700344 |
| [39] | Rasmussen P E, Kresge P O. Plant response to sulfur in the Western United States. Agronomy. A Series of Monographs - American Society of Agronomy, 1986: 357–374. |
| [40] | 胡星云, 孙志高, 孙文广, 王苗苗, 王伟, 田莉萍. 黄河口新生湿地碱蓬生物量及氮累积与分配对外源氮输入的响应. 生态学报, 2017, 37(1): 226–237. |
| [41] | Sun Z G, Liu J S. Distribution and fate of anthropogenic nitrogen in the Calamagrostis angustifolia wetland ecosystem of Sanjiang Plain, Northeast China. Journal of Integrative Plant Biology, 2008, 50(4): 402–414. DOI:10.1111/jipb.2008.50.issue-4 |
| [42] | Xie Y H, Wen M Z, Yu D, Li Y K. Growth and resource allocation of water hyacinth as affected by gradually increasing nutrient concentrations. Aquatic Botany, 2004, 79(3): 257–266. DOI:10.1016/j.aquabot.2004.04.002 |
| [43] | Haraguchi A, Hasegawa C, Hirayama A, Kojimab H. Decomposition activity of peat soils in geogenous mires in Sasakami, central Japan. European Journal of Soil Biology, 2003, 39(4): 191–196. DOI:10.1016/j.ejsobi.2003.05.001 |
| [44] | Davis S M. Growth, decomposition, and nutrient retention of Cladium jamaicense Crantz and Typha domingensis Pers. in the Florida Everglades. Aquatic Botany, 1991, 40(3): 203–224. DOI:10.1016/0304-3770(91)90059-E |
| [45] | Sun Z G, Mou X J, Lin G H, Wang L L, Song H L, Jiang H H. Effects of sediment burial disturbance on seedling survival and growth of Suaeda salsa in the tidal wetland of the Yellow River estuary. Plant and Soil, 2010, 337(1/2): 457–468. |
| [46] | 孙志高, 牟晓杰, 王玲玲, 孙万龙, 孙文广. 黄河口潮滩盐沼沉积强度对碱蓬残体分解及氮动态的影响. 湿地科学, 2015, 13(2): 135–144. |
| [47] | 曹志洪, 孟赐福, 胡正义. 中国农业与环境中的硫. 北京: 科学出版社, 2011: 1–83. |