 2018, Vol. 38
2018, Vol. 38文章信息
- 吴晓敏, 郝瑞娟, 王丽卿, 潘宏博.
- WU Xiaomin, HAO Ruijuan, WANG Liqing, PAN Hongbo.
- 景观水体周丛生纤毛虫群落结构及其与环境因子的关系
- Community structure of periphytic ciliates in scenic water and its relationship with environmental factors
- 生态学报. 2018, 38(15): 5541-5553
- Acta Ecologica Sinica. 2018, 38(15): 5541-5553
- http://dx.doi.org/10.5846/stxb201709281752
-
文章历史
- 收稿日期: 2017-09-28
- 网络出版日期: 2018-04-19





2. 上海海洋大学农业部鱼类营养与环境生态研究中心, 上海 201306;
3. 上海海洋大学水产种质资源发掘与利用教育部重点实验室, 上海 201306
2. Centre for Research on Environmental Ecology and Fish Nutrient of the Ministry of Agriculture, Shanghai Ocean University, Shanghai 201306, China;
3. Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Shanghai Ocean University, Ministry of Education, Shanghai 201306, China
与高等真核生物相比, 原生动物为单细胞动物, 它分布广泛, 能更直接与它所生存的环境接触, 对环境变化如水体污染的反应更敏感[1]。因此, 在水质监测中, 原生动物可作为水体污染评价理想的环境指示生物[2-6]。
周丛生纤毛虫是水域生态系统的重要组成部分, 其在微食物网和生态系统的物质循环以及能量流动中起着非常重要的作用[7-10]。纤毛虫生长速度快, 可短时间在人造基质(如载玻片)上群集并获得较高的密度和丰度, 成为周丛生群落的主要类群[11]。自Cairns等[12]首次用聚氨酯泡沫(PFU)法采集微型生物群落, 周丛生纤毛虫已被广泛地用于生态调查的水质评估和监测中[11, 13-14]。
目前用于生物监测的周丛生纤毛虫采样法主要是PFU法和载玻片法两种。沈蕴芬等[15-16]对原生动物群落进行详尽的研究, 并对PFU法进行改进和完善, 将其发展成国标, 从而用于测试水体的污染。近十几年来, PFU法广泛应用于淡水河流、湖泊和海洋的水质监测[5, 17-20], 载玻片法多应用于海洋监测[4, 21-22]。由于景观水体面积小、水浅, PFU上的小孔极易被搅起的底质或大颗粒悬浮物堵塞而造成群落结构失真, 无法正确监测水质, 载玻片法则可以避免这个问题。基于载玻片法的生物监测在淡水尤其是景观水体研究极少, 其上生长的纤毛虫等原生动物群落能否较好反映景观水体水质还有待确定。本研究于2015年1—12月在上海市东南角的一处浅型景观水体中利用载玻片法采集周丛生纤毛虫, 并分析其群落结构与环境因子的关系, 以期为周丛生原生动物的生态学研究提供第一手参考资料; 同时, 为评估景观水体的负荷力和能量流动规律之间的关系、建立景观水体中各个相关功能类群的动力学参数以及景观水体的水质监测等方面提供重要的资料。
1 材料与方法 1.1 采样时间和地点2015年1—12月逐月采集上海市东南角一处人工景观水体(地理坐标为30°53′11.01′′N, 121°53′26.85′′E)的周丛生纤毛虫。该水体位于上海南汇东滩湿地, 横穿上海海洋大学校园, 其一端有闸口, 外接芦潮引河直入东海。总水体面积约为0.061 km2, 平均水深约为3.20 m, 海拔约为4 m[23]。
1.2 样品采集与处理本研究利用载玻片(2.6 cm×7.6 cm)作为人工附着基进行景观水体周丛生纤毛虫的采集, 使用前将载玻片浸泡在酒精中24 h后进行燃烧去除其表面脂结构。每个月放置8个载玻片框(112张载玻片)于湖中, 采集的载玻片框(每框内置14片载玻片, 两两背对着放置于框内的一个玻片槽中)悬挂于水深1 m处(载玻片框装置参考Xu等[4]), 待周丛生纤毛虫群落达到成熟(春、夏、秋季5—8 d, 冬季12—14 d)后将载玻片框取出, 并放于原位水中带回实验室。
1.3 种类鉴定与定量研究在体视显微镜下对一个载玻片框中的14张载玻片进行全片观察(65×)以获取种类数; 随机选取2张载玻片, 每张随机选取10个视野进行物种密度的统计[4], 所有的计数均在12 h内完成[4]。种类鉴定主要参考文献[16, 24-29]。
1.4 水质参数的测定酸碱度(pH)、溶解氧(DO)、水温(T)、电导率(Spc)、盐度(Sal)用便携式YSI ProPlus进行现场测定, 透明度(SD)用透明度盘现场测定。用5 L有机玻璃采水器采集混合水样, 取1 L混合水样于聚乙烯瓶中, 置于4℃冷藏箱中, 4 h内完成总氮(TN)、总磷(TP), 叶绿素a(Chl-a), 高锰酸盐指数(CODMn)和亚硝酸盐氮(NO2--N)等指标的测定, 具体测定方法参照《水和废水监测分析方法(第四版)》[30]进行。
1.5 数据统计和分析 1.5.1 优势度的计算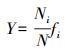
选取Shannon-Wiener多样性指数(H′)[32], Margalef丰富度指数(D)[33]和Pielou均匀度指数(J′)[34]来分析周丛生纤毛虫群落结构的多样性。
Shannon-Wiener多样性指数(H′):
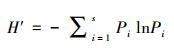
|
Margalef丰富度指数(D):
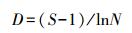
|
Pielou均匀度指数(J′):

|
式中, Pi=Ni/N, Ni为第i物种的个体数, N表示样品中所有种类的总个体数, S为样品中物种的总种类数。
1.5.3 分析软件本研究数据的处理和分析采用SPSS 18.0软件进行Pearson相关性分析; 折线和柱状图均采用OriginPro 2016绘制; 采用Canoco for Windows 4.5软件对周丛生纤毛虫优势种的密度进行ln(X+1)转化, 并将其与环境因子进行冗余分析(Redundancy analysis, RDA); 采用PRIMER 6软件对周丛生纤毛虫密度进行平方根转化、对环境变量进行Log(X+1)转化并进行聚类分析(CLUSTER)以及生物-环境变量的相关性分析(BIOENV)。
2 结果与分析 2.1 水体环境因子的周年变化本研究中, 水体高锰酸盐指数(CODMn)的年均值为(6.82±1.04) mg/L, 变化范围为5.22—8.58 mg/L, 5月最低, 8月最高; 叶绿素a(Chl-a)浓度的年均值为(0.02±0.02) mg/L, 变化范围为0.01—0.07 mg/L, 8月浓度最高。
亚硝酸盐氮(NO2--N)浓度的年均值为(0.07±0.02) mg/L, 变化范围为0.04—0.08 mg/L, 3月含量最低, 5月和7月含量最高; 总氮(TN)的年均值为(1.21±0.37) mg/L, 变化范围为0.77—1.97 mg/L, 7月含量最低, 1月含量最高; 总磷(TP)的年均值为(0.08±0.03) mg/L, 变化范围为0.05—0.15 mg/L, 1月含量最高。
透明度(SD)年均值为(98.50±8.98) cm, 其变化范围为80—110 cm, 1月透明度最低, 最高值出现在8、10和12月; 水温(T)的平均值为(19.14±7.66)℃, 变化范围为7.60—29.60℃, 全年水温整体呈先升高后降低的趋势, 最低水温出现在2月, 最高水温出现在7月; 溶解氧(DO)的年均值为(6.64±3.45) mg/L, 变动范围为2.44—11.01 mg/L, 6月最低, 4月最高; 电导率(Spc)的年均值为(971.09±182.57) μs/cm, 变化范围为750.00—1282.00 μs/cm, 9月最低, 6月最高; 年均盐度(Sal)为(0.56±0.07)‰, 变化范围为0.44—0.64‰, 10月最低, 5月最高; 酸碱度(pH)年平均值为(7.75±0.31), 在7.15—8.35之间变动, 11月最低, 2月最高, 表明本研究水体的水质酸碱度呈中性偏碱性(表 1)。
| 月份 Month |
高锰酸盐指数 Permanganate index/(mg/L) |
叶绿素a Chlorophyll a/(mg/L) |
亚硝酸盐氮 Nitrite nitrogen/(mg/L) |
总氮 Total nitrogen/(mg/L) |
总磷 Total phosphorus/(mg/L) |
透明度 Transparency/cm |
水温 Water temperature/℃ |
溶解氧 Dissolved oxygen/(mg/L) |
电导率 Conductivity/(μs/cm) |
盐度 Salinity/‰ |
pH |
| 1 | 8.00 | 0.03 | 0.06 | 1.97 | 0.15 | 80.00 | 7.90 | 8.54 | 794.00 | 0.59 | 7.48 |
| 2 | 6.81 | 0.02 | 0.06 | 1.51 | 0.05 | 90.00 | 7.60 | 10.41 | 809.00 | 0.61 | 8.35 |
| 3 | 6.79 | 0.02 | 0.04 | 1.11 | 0.12 | 95.00 | 15.10 | 9.67 | 1016.00 | 0.63 | 7.98 |
| 4 | 6.52 | 0.01 | 0.07 | 1.22 | 0.05 | 95.00 | 17.30 | 11.01 | 957.00 | 0.56 | 7.66 |
| 5 | 5.22 | 0.04 | 0.08 | 1.08 | 0.10 | 100.00 | 24.30 | 9.01 | 1275.00 | 0.64 | 7.89 |
| 6 | 6.63 | 0.02 | 0.07 | 1.06 | 0.13 | 100.00 | 24.20 | 2.44 | 1282.00 | 0.63 | 7.44 |
| 7 | 7.34 | 0.02 | 0.08 | 0.77 | 0.10 | 92.00 | 29.60 | 4.11 | 1009.00 | 0.45 | 7.66 |
| 8 | 8.58 | 0.07 | 0.06 | 1.20 | 0.07 | 110.00 | 28.67 | 2.65 | 1150.10 | 0.53 | 7.60 |
| 9 | 7.11 | 0.02 | 0.05 | 1.21 | 0.07 | 100.00 | 25.00 | 2.67 | 750.00 | 0.50 | 7.91 |
| 10 | 7.84 | 0.01 | 0.07 | 0.74 | 0.05 | 110.00 | 21.60 | 4.43 | 834.00 | 0.44 | 7.92 |
| 11 | 5.39 | 0.01 | 0.06 | 0.90 | 0.05 | 100.00 | 17.70 | 4.42 | 927.00 | 0.54 | 7.15 |
| 12 | 5.66 | 0.01 | 0.10 | 1.74 | 0.07 | 110.00 | 10.70 | 10.32 | 850.00 | 0.58 | 7.94 |
| 年均值 Annual average value |
6.82±1.04 | 0.02±0.02 | 0.07±0.02 | 1.21±0.37 | 0.08±0.03 | 98.50±8.98 | 19.14±7.66 | 6.64±3.45 | 971.09±182.57 | 0.56±0.07 | 7.75±0.31 |
周丛生纤毛虫的年平均密度为127.29个/cm2, 年密度变化在24.27—248.57个/cm2之间, 2月密度最低, 密度最高峰出现在5月, 在8月出现次高峰。在全年的研究中, 共鉴定周丛生纤毛虫51种, 隶属于12目。种类数为春季>冬季>夏季>秋季。1月检出的周丛生纤毛虫种类最多, 2月种类数最少, 并且到5月一直处于增加的趋势, 变化范围为10—27种; 6—12月的种类数波动较小, 在11—17种之间变化(图 1)。按照分类阶元划分, 缘毛目纤毛虫所占比例最高, 为35%;其次为侧口目纤毛虫, 所占比例为12%, 裸口目、管口目、毛口目和膜口目纤毛虫所占比例最低, 均为2%(附表); 在全年的各个月份, 缘毛目纤毛虫的种类数所占比例均最高(图 2)。

|
| 图 1 周丛生纤毛虫的种类数及密度的周年变化 Fig. 1 Annual variations of species number and abundance of periphytic ciliates |
| 目 Orders |
种类 Species |
1月 Jan. |
2月 Feb. |
3月 Mar. |
4月 Apr. |
5月 May |
6月 Jun. |
7月 Jul. |
8月 Aug. |
9月 Sep. |
10月 Oct. |
11月 Nov. |
12月 Dec. |
| 缘毛目 | 沟钟虫Vorticella convallaria | + | + | + | ++ | ++++ | + | + | + | +++ | +++ | +++ | |
| Peritrichida | 钟形钟虫Vorticella campanula | ++ | + | + | ++ | ++ | ++ | +++ | ++ | +++ | ++ | ++ | |
| Vorticella influsinum | ++ | + | ++ | + | + | + | + | + | + | + | |||
| 伪钟虫sp. Pseudovorticella sp. | + | ||||||||||||
| 钟虫sp. 1 Vorticella sp. 1 | + | + | |||||||||||
| 钟虫sp. 2 Vorticella sp. 2 | + | + | +++ | ++ | +++ | ++ | |||||||
| 钟虫sp. 3 Vorticella sp. 3 | + | + | |||||||||||
| 伞状聚钟虫Campanella umbellaria | + | + | + | + | + | + | + | + | |||||
| 聚缩虫sp. 1 Zoothamnium sp. 1 | + | ++ | + | + | + | ++ | + | + | ++ | + | + | ||
| 螅状独缩虫Carchesium polypinum | + | ++ | ++ | ++ | + | + | + | + | |||||
| 聚缩虫sp. 2 Zoothamnium sp. 2 | + | ||||||||||||
| 累枝虫sp. 1 Epistylis sp. 1 | + | ||||||||||||
| 累枝虫sp. 2 Epistylis sp. 2 | ++ | + | + | ++ | + | + | |||||||
| 亨氏累枝虫Epistylis hentscheli | + | + | ++ | + | ++ | +++ | ++ | ++ | +++ | ++ | + | + | |
| 累枝虫sp. 3 Epistylis sp. 3 | + | ||||||||||||
| 累枝虫sp. 4 Epistylis sp. 4 | + | + | |||||||||||
| 果盖虫Orbopcrcularia berberina | + | + | |||||||||||
| 躺平鞘居虫Platycola decumbens | + | ||||||||||||
| 寡毛目 | 弹跳虫sp. Collembola sp. | + | + | ||||||||||
| Oligotrichida | 大弹跳虫Halteria grandinella | + | |||||||||||
| 急游虫sp. Stombidium sp. | + | ||||||||||||
| 楯纤虫Aspidisca | + | ||||||||||||
| 吸管目 | 四分锤吸管虫Tokophrya quadripartita | + | + | + | |||||||||
| Suctorida | 华丽十字吸管虫Staurophrya elegans | + | + | + | + | ||||||||
| 长柄球吸管虫Metacineta macrocaulis | + | + | + | ||||||||||
| 累枝毛吸管虫Trichophrya epistylidis | + | ||||||||||||
| Denarosoma radians | + | + | |||||||||||
| 异毛目 | 带核喇叭虫Stentor roeseli | + | + | + | + | + | + | + | |||||
| Heterotrichida | 多态喇叭虫Stentor polymorphrus | + | + | ||||||||||
| 丁丁目 | 淡水筒壳虫Tintinnidium fluviatile | + | + | + | + | ||||||||
| Tintinnida | 恩茨筒壳虫Tintinnidium entzii | + | + | + | |||||||||
| 小筒壳虫Tintinnidium pusillum | + | ||||||||||||
| 樽形拟铃虫Tintinnopsis potiformis | + | ||||||||||||
| 裸口目 Protostomatida |
毛板壳虫Coleps hirtus | + | |||||||||||
| 刺钩目 | 双环栉毛虫Didinium nasufum | + | |||||||||||
| Haptorida | Paradifetus elephantimus | + | + | + | |||||||||
| 卵圆口虫Trachelius ovum | + | + | + | + | + | + | + | ||||||
| 天鹅长吻虫Lacrymaria olor | + | ||||||||||||
| 管口目 Cyrtophorida |
钩刺斜管虫Chilodonella uncineta | + | + | + | + | + | + | + | + | + | |||
| 毛口目 Trichostomatida |
斜口虫sp. Plagiopyla sp. | + | |||||||||||
| 侧口目 | 裂口虫sp. 1 Amphileptus sp. 1 | + | |||||||||||
| Pleurostomatida | 裂口虫sp. 2 Amphileptus sp. 2 | + | + | ||||||||||
| 拟裂口虫sp. Amphileptiscus sp. | + | ||||||||||||
| 裂口虫sp. 3 Amphileptus sp. 3 | + | + | + | ||||||||||
| Amphileptus carchesii | + | + | |||||||||||
| 天鹅漫游虫Litonotus cygnus | + | + | + | + | + | + | |||||||
| 膜口目 Hymenostomatida |
前口虫sp. Frontonia sp. | + | + | ||||||||||
| 腹毛目 | 贻贝棘尾虫Stylonychia mytilus | + | + | + | |||||||||
| Hypotrichida | 尾柱虫sp. Urostyla sp. | + | + | ||||||||||
| 尖毛虫sp. Oxytricha sp. | + | + | + | + | + | + | + | + | + | ||||
| 伍氏游仆虫Euplotes woodruffi | + | + | |||||||||||
| 周丛生纤毛虫的平均密度用+, ++, +++, ++++表示(+ = 0.02—9.99个/cm2; ++ = 10.00—50.00个/cm2; +++ = 50.01—100.00个/cm2; ++++ = 100.01—160.00个/cm2) | |||||||||||||

|
| 图 2 周丛生纤毛虫各目的种类数所占比例的周年变化 Fig. 2 Annual variations of percentage of species number on different orders of periphytic ciliates |
沟钟虫(Vorticella convallaria), 钟形钟虫(Vorticella campanula), Vorticella influsinum, 钟虫sp. 2(Vorticella sp. 2), 聚缩虫sp. 1(Zoothamnium sp. 1), 螅状独缩虫(Carchesium polypinum), 亨氏累枝虫(Epistylis hentscheli)为优势种(表 2)。沟钟虫密度在春季特别是5月最高, 其次是在10—12月密度较高; 钟形钟虫密度除在2月和6月以外的其他月份均较高; Vorticella influsinum的密度在1月和3月出现两次峰值; 钟虫sp. 2在6—8月密度较高, 其他月份密度均较低; 聚缩虫sp. 1在整个年度出现3次峰值, 分别为3月, 7月和10月, 其他月份密度都较低; 螅状独缩虫在12—4月的密度均较高, 其他月份密度较低; 亨氏累枝虫密度在整个年度出现两次峰值, 分别为6月与9月, 整体为5—9月密度最高, 其他月份密度较低(图 3)。
| 优势种Dominant species | 优势度(Y)Dominance |
| 沟钟虫Vorticella convallaria | 0.21 |
| 钟形钟虫Vorticella campanula | 0.20 |
| Vorticella influsinum | 0.03 |
| 钟虫sp. 2 Vorticella sp. 2 | 0.06 |
| 聚缩虫sp. 1 Zoothamnium sp. 1 | 0.07 |
| 螅状独缩虫Carchesium polypinum | 0.03 |
| 亨氏累枝虫Epistylis hentscheli | 0.19 |

|
| 图 3 周丛生纤毛虫优势种密度的周年变化 Fig. 3 Annual variations of abundance on dominant periphytic ciliates |
周丛生纤毛虫H′值从1—12月呈缓慢下降的趋势, 变幅为1.05—2.10, 年均值为1.53;D值变幅为1.91—6.33, 年均值为3.16, 变化情况与H′接近, 且在8月最低, 1月最高; J′值变幅为0.41—0.72, 各月变化不大, 年均值为0.56(图 4)。

|
| 图 4 周丛生纤毛虫多样性指数的周年变化 Fig. 4 Annual variations of diversity index of periphytic ciliates |
在相似系数为50%时, 景观水体周丛生纤毛虫可分为4个类群(ANOSIM, P < 0.001):第一类群(Ⅰ)为在2月聚集的周丛生纤毛虫群落; 第二类群(Ⅱ)为在6月聚集的周丛生纤毛虫群落; 第三类群(Ⅲ)为在1月和3月聚集的周丛生纤毛虫群落; 将其余月份聚集的周丛生纤毛虫群落归为第四类群(Ⅳ)。在相似系数为65%时, 可将第四类群的周丛生纤毛虫分为四类:第一类(Ⅳa)为在7, 8和9月聚集的周丛生纤毛虫群落; 第二类(Ⅳb)为在5月聚集的周丛生纤毛虫群落; 第三类(Ⅳc)为在4月聚集的周丛生纤毛虫群落; 第四类(Ⅳd)为在10, 11月和12月聚集的周丛生纤毛虫群落(图 5)。

|
| 图 5 周丛生纤毛虫群落的聚类分析树状图 Fig. 5 Clustering dendrogram of the periphytic ciliates communities |
对周丛生纤毛虫群落结构参数与水温(T)、溶解氧(DO)以及电导率(Spc)等11个水质参数进行Pearson相关性分析, 周丛生纤毛虫的月平均密度与水体温度呈极显著正相关(P < 0.01), 与水体电导率呈显著正相关(P < 0.05);丰富度指数与水体透明度呈极显著负相关(P < 0.01)(表 3)。对周丛生纤毛虫各个种的密度与环境因子进行BIOENV分析, 在生物—环境相关性排序前10的环境变量组合中, 总磷(TP)浓度和水温(T)是影响周丛生纤毛虫群落的主要环境因子, 其中, 周丛生纤毛虫的群落结构与TP、T、DO、pH(ρ值:0.625, P:0.001), TP、SD、T、DO、pH(ρ值:0.619, P:0.001)这两个环境因子组合的相关性最高(表 4)。
| 群落结构参数 Community structural parameters |
高锰酸盐指数 Permanganate index/(mg/L) |
亚硝酸盐氮 Nitrite nitrogen/(mg/L) |
总氮 Total nitrogen/(mg/L) |
总磷 Total phosphorus/(mg/L) |
叶绿素a Chlorophyll a/(mg/L) |
透明度 Transparency/cm |
水温 Water temperature/℃ |
溶解氧 Dissolved oxygen/(mg/L) |
电导率 Conductivity/(μs/cm) |
盐度 Salinity |
pH |
| 种类数 Species number |
0.13 | -0.13 | 0.27 | 0.49 | -0.02 | -0.53 | -0.20 | 0.37 | -0.07 | 0.15×10-3 | -0.09 |
| 密度 Abundance |
-0.07 | 0.18 | -0.41 | 0.15 | 0.49 | 0.56 | 0.73** | -0.4 | 0.67* | 0.01×10-3 | -0.04 |
| 多样性指数 Shannon-Wiener index |
0.49 | -0.41 | 0.27 | 0.42 | 0.08 | -0.55 | -0.20 | 0.29 | -0.34 | -0.11×10-3 | 0.11 |
| 丰富度指数 Margalef index |
0.19 | -0.19 | 0.45 | 0.45 | -0.11 | -0.72** | -0.45 | 0.48 | -0.27 | 0.19×10-3 | -0.05 |
| 均匀度指数 Pielou index |
0.52 | -0.38 | 0.21 | 0.18 | 0.16 | -0.34 | -0.14 | 0.15 | -0.38 | -0.22×10-3 | 0.23 |
| *在0.05水平(双侧)上显著相关; **在0.01水平(双侧)上显著相关 | |||||||||||
| 排序 Rank |
环境变量 Environment variables |
ρ | 显著性水平 P |
| 1 | TP, T, DO, pH | 0.625 | 0.001 |
| 2 | TP, SD, T, DO, pH | 0.619 | 0.001 |
| 3 | TP, T | 0.613 | 0.002 |
| 4 | TP, T, DO | 0.603 | 0.001 |
| 5 | TP, SD, T, DO | 0.594 | 0.001 |
| 6 | TP, T, pH | 0.587 | 0.005 |
| 7 | TP, T, DO, Spc, pH | 0.586 | 0.001 |
| 8 | TP, SD, T, pH | 0.549 | 0.009 |
| 9 | TN, TP, T, DO, pH | 0.543 | 0.002 |
| 10 | TP, T, DO, Sal, pH | 0.537 | 0.001 |
| TP:总磷, Total phosphorus; T:水温, Water temperature; DO:溶解氧, Dissolved oxygen; pH;SD:透明度, Transparency; Spc:电导率, Conductivity; TN:总氮, Total nitrogen; Sal:盐度, Salinity | |||
Pearson相关性分析结果表明, 缘毛目纤毛虫密度与透明度以及电导率呈显著正相关(P < 0.05), 与水温呈极显著正相关(P < 0.01);丁丁目、毛口目和侧口目纤毛虫密度与透明度呈显著负相关(P < 0.05), 其中丁丁目纤毛虫密度与水温呈极显著负相关(P < 0.01)、与总氮浓度呈极显著正相关(P < 0.01), 毛口目纤毛虫密度与总氮和总磷浓度均呈显著正相关(P < 0.05)(表 5)。
| 目 Orders |
高锰酸盐指数 Permanganate index/(mg/L) |
亚硝酸盐氮 Nitrite nitrogen/(mg/L) |
总氮 Total nitrogen/(mg/L) |
总磷 Total phosphorus/(mg/L) |
叶绿素a Chlorophyll a/(mg/L) |
透明度 Transparency/cm |
水温 Water temperature/℃ |
溶解氧 Dissolved oxygen/(mg/L) |
电导率 Conductivity/(μs/cm) |
盐度 Salinity/‰ |
pH |
| 缘毛目Peritrichida | -0.08 | 0.20 | -0.41 | 0.12 | 0.49 | 0.59* | 0.72** | -0.41 | 0.66* | 0.01 | -0.05 |
| 寡毛目Oligotrichida | 0.15 | -0.30 | 0.10 | 0.48 | -0.25 | -0.34 | -0.23 | 0.22 | 0.16 | 0.36 | -0.20 |
| 吸管目Suctorida | 0.02 | -0.54 | -0.08 | 0.13 | -0.15 | -0.16 | -0.30 | 0.43 | -0.04 | 0.31 | 0.44 |
| 异毛目Heterotrichida | 0.12 | 0.27 | -0.46 | 0.25 | -0.01 | -0.17 | 0.55 | -0.42 | 0.26 | -0.40 | -0.20 |
| 丁丁目Tintinnida | 0.25 | -0.11 | 0.81** | 0.41 | -0.04 | -0.67* | -0.79** | 0.47 | -0.47 | 0.34 | 0.18 |
| 裸口目Protostomatida | -0.09 | 0.04 | 0.01 | -0.29 | -0.27 | -0.12 | -0.08 | 0.40 | -0.02 | 0.01 | -0.09 |
| 刺钩目Haptorida | 0.22 | -0.02 | -0.18 | 0.14 | -0.03 | -0.41 | 0.37 | -0.30 | -0.25 | -0.57 | -0.18 |
| 管口目Cyrtophorida | -0.04 | -0.18 | -0.15 | 0.15 | -0.21 | 0.01 | -0.04 | 0.20 | -0.12 | -0.09 | 0.16 |
| 毛口目Trichostomatida | 0.36 | -0.12 | 0.65* | 0.59* | 0.08 | -0.65* | -0.46 | 0.17 | -0.31 | 0.15 | -0.27 |
| 侧口目Pleurostomatida | 0.25 | -0.19 | 0.53 | 0.32 | -0.15 | -0.61* | -0.43 | 0.40 | -0.33 | 0.12 | -0.20 |
| 膜口目Hymenostomatida | -0.09 | 0.01 | 0.13 | -0.41 | -0.29 | -0.25 | -0.29 | 0.53 | -0.15 | 0.12 | 0.20 |
| 腹毛目Hypotrichida | 0.04 | 0.01 | -0.34 | 0.12 | -0.06 | -0.08 | 0.32 | -0.03 | 0.03 | -0.35 | -0.04 |
| *在0.05水平(双侧)上显著相关; **在0.01水平(双侧)上显著相关 | |||||||||||
对不同优势种的周从生纤毛虫密度与环境因子进行冗余分析(RDA)。前两个排序轴的特征值分别为0.42和0.29, 第一排序轴解释了样本中42%的变异, 第二排序轴解释了样本29%的变异, 两者合并解释了样本71.10%的总变异(表 6)。T、SD、DO、TN、TP、CODMn和Sal对周丛生纤毛虫各个优势种的影响较大(图 6)。其中, 钟形钟虫、钟虫sp. 2、聚缩虫sp. 1、亨氏累枝虫与SD和T呈正相关, 与DO、TN和Sal呈显著负相关; 沟钟虫与SD呈显著正相关, 与TN、TP、CODMn和Sal呈显著负相关; 螅状独缩虫与T、SD呈显著负相关, 与DO、TN和Sal呈显著正相关。
| 冗余分析的统计计量 Statistical measurements of redundancy analysis |
第一排序轴 The first axis |
第二排序轴 The second axis |
第三排序轴 The third axis |
第四排序轴 The fourth axis |
| 特征值Eigenvalues | 0.42 | 0.29 | 0.11 | 0.07 |
| 样本-环境关联度 Species-environment correlations |
1.00 | 1.00 | 1.00 | 1.00 |
| 物种累积关联度百分数 Cumulative percentage variance of species data |
42.40 | 71.10 | 82.20 | 89.20 |
| 物种-环境关系累计百分比 Cumulative percentage variance of species-environment relation |
42.40 | 71.10 | 82.20 | 89.20 |

|
| 图 6 周丛生纤毛虫优势种与环境因子的冗余分析 Fig. 6 Redundancy analysis based on relationships between dominant periphytic ciliates and environmental factors V. con:沟钟虫, Vorticella convallaria; V. cam:钟形钟虫, Vorticella campanula; V. inf:Vorticella influsinum; V. sp. 2:钟虫sp. 2, Vorticella sp. 2;Z. sp. 1:聚缩虫sp. 1, Zoothamnium sp. 1;C. pol:螅状独缩虫, Carchesium polypinum; E. hen:亨氏累枝虫, Epistylis hentscheli |
周丛生纤毛虫的群落结构在沿海水域、海湾及湖泊水体中均具有季节性差异, 其应对水体环境变化也呈现显著的季节性变动[11, 35]。Gong等[13]在胶州湾的周年研究中共检测出周丛生纤毛虫37种(30属)。Xu等[21]在韩国沿海水体中检测出周丛生纤毛虫29种。本研究周丛生纤毛虫(51种)的物种多样性显著高于上述近海岸的开放水体, 这可能是与近海岸的开放水体相比, 景观水体中的周丛生纤毛虫种类不易受潮汐扰动有关。而本研究所检测出的周丛生纤毛虫物种数显著低于Agamaliev[36]在里海水中检测出的周丛生纤毛虫物种数(130种), 这可能与本研究的采样范围较小导致物种多样性较低有关。此外, 本研究结果表明, 该景观水体中的周丛生纤毛虫的群落结构存在明显的季节性差异, 其中, 冬季1月检测的周丛生纤毛虫种类最多, 共27种; 冬季2月种类数最少, 共10种; 周丛生纤毛虫以缘毛目纤毛虫为优势类群, 其密度与种类数在全年的各个月份均占绝对优势, 且全年的优势种均隶属于缘毛目, 这可能与以载玻片作为人工基质、水体的水质状态以及食物较适合其生长有关。
3.2 周丛生纤毛虫群落结构与环境因子的关系水体环境因子(如溶解氧、水温、硝酸盐氮、亚硝酸盐氮、可溶性磷酸盐、氨氮、盐度等)的变动是影响周丛生纤毛虫群落结构分布的重要因素。已有研究表明, 海水养殖水体中周丛生纤毛虫的丰度和物种数量与可溶性磷酸盐具有极显著正相关关系[37], 周丛生纤毛虫的密度与水体硝酸盐(NO3-N)和总磷(TP)的浓度呈正相关[38]。本研究的BIOENV分析也发现类似的结果, 总磷(TP)浓度和水温(T)是影响周丛生纤毛虫密度的主要环境因子(表 4)。
缘毛目纤毛虫的密度与温度和透明度呈极显著或显著正相关关系(表 5)。可能因为采样水温为7.60—29.60℃, 原生动物的种群增长率在该温度范围内随温度的升高而升高[39]; 同时它们为滤食性种类, 主要以微微型浮游生物为食[40], 通过其捕食能够提高水体透明度。本研究中, 漫游类的侧口目纤毛虫密度与水体透明度呈显著负相关, 这表明采集的侧口目纤毛虫更适宜于生存在透明度较低的水体中。虽然毛口目纤毛虫也与环境因子显著相关(与总氮浓度及总磷浓度呈显著正相关, 与透明度呈显著负相关), 但因仅发现一种Plagiopyla sp.且只在1月检出, 其相关性分析结果说服力较差。
不同种类最适温度的范围各异, 如一些丁丁目的周丛生纤毛虫主要喜低温[41]。本研究中, 丁丁目周丛生纤毛虫密度与水温呈极显著负相关(P < 0.01), 丁丁目的周丛生纤毛虫的主要种类如淡水筒壳虫(Tintinnidium fluviatile)和恩茨筒壳虫(Tintinnidium entzii)主要出现在温度较低的冬季, 可能是由于这类周丛生纤毛虫喜低温, 同时一些固着类的吸管目、缘毛目周丛生纤毛虫繁殖速度变缓, 使得载玻片上的物种间的空间竞争速度减慢, 丁丁目纤毛虫为代表的浮游类纤毛虫竞争压力变小, 密度增大, 从而也为该类物种在载玻片上提供了一个生存所需的空间。
3.3 优势种对水质的指示作用在沿海水域、淡水及半咸水水体中, 周丛生纤毛虫优势种多为缘毛目纤毛虫, 其对不同类型的水体均具有一定的指示作用。如在青岛沿海的优势种聚缩虫未定种(Zoothamnium sp.)与pH、溶解氧以及营养盐显著相关[35]; 武汉东湖的优势种钟形钟虫与水温具有相关性; 而青岛胶州湾的优势种中国伪钟虫(Pseudovorticella sinensis)与较低的溶解氧(3.80 mg/L)有关[13]。本研究的RDA分析表明, 优势种的密度与水体的环境因子具有较大的相关性(图 6)。其中, 钟形钟虫、钟虫sp. 2、聚缩虫sp. 1、亨氏累枝虫、沟钟虫和螅状独缩虫这6个种的存在以及是否成为优势类群均可以考虑用作评定水体的质量状态, 并且可以作为水体富营养化以及有机污染的潜在指示生物。但这一结论仍需进一步的实验进行验证。
综上所述, 本研究发现载玻片法采集的周丛生纤毛虫原生动物同样也能反映景观水体水质。由于景观水体有面积小和水浅的特点, PFU上的小孔极易被搅起的底质或大颗粒悬浮物堵塞而造成群落结构失真无法正确监测水质, 载玻片法可作为PFU法的一个良好补充或替代。
致谢: 感谢上海海洋大学许朋和邢宏伟同学在样品采集和理化因子测定方面提供的帮助。| [1] | Payne R J. Seven reasons why protists make useful bioindicators. Acta Protozoologica, 2013, 52(3): 105–113. |
| [2] | Cairns J Jr, Lanza G R, Parker B C. Pollution related structural and functional changes in aquatic communities with emphasis on freshwater algae and protozoa. Proceedings of the Academy of Natural Sciences of Philadelphia, 1972, 124: 79–127. |
| [3] | Gomes E A T, Godinho M J L. Structure of the protozooplankton community in a tropical shallow and eutrophic lake in Brazil. Acta Oecologica, 2003, 24(S1): S153–S161. |
| [4] | Xu H L, Min G S, Choi J K, Jung J H, Park M H. An approach to analyses of periphytic ciliate colonization for monitoring water quality using a modified artificial substrate in Korean coastal waters. Marine Pollution Bulletin, 2009, 58(9): 1278–1285. DOI:10.1016/j.marpolbul.2009.05.003 |
| [5] | 许木启, 曹宏. PFU原生动物群落生物监测的生态学原理与应用. 生态学报, 2004, 24(7): 1540–1547. |
| [6] | 彭晓辉. 杭州西湖水域原生动物物种多样性及缘毛目一新属/种的形态学研究[D]. 杭州: 杭州师范大学, 2007. |
| [7] | Azovsky A I. Colonization of sand "islands" by psammophilous ciliates:the effect of microhabitat size and stage of succession. Oikos, 1988, 51(1): 48–56. DOI:10.2307/3565806 |
| [8] | Corliss J O. Biodiversity and biocomplexity of the protists and an overview of their significant roles in maintenance of our biosphere. Acta Protozoologica, 2002, 41(3): 199–219. |
| [9] | Norf H, Arndt H, Weitere M. Effects of resource supplements on mature ciliate biofilms:an empirical test using a new type of flow cell. Biofouling, 2009, 25(8): 769–778. DOI:10.1080/08927010903174581 |
| [10] | Xu H L, Zhang W, Jiang Y, Yang E J. Use of biofilm-dwelling ciliate communities to determine environmental quality status of coastal waters. Science of the Total Environment, 2014, 470-471: 511–518. DOI:10.1016/j.scitotenv.2013.10.025 |
| [11] | Coppellotti O, Matarazzo P. Ciliate colonization of artificial substrates in the Lagoon of Venice. Journal of the Marine Biological Association of the United Kingdom, 2000, 80(3): 419–427. DOI:10.1017/S0025315400002113 |
| [12] | Cairns J Jr, Dahlberg M L, Dickson K L, Smith N, Waller W T. The relationship of fresh-water protozoan communities to the MacArthur-Wilson equilibrium model. The American Naturalist, 1969, 103(933): 439–454. DOI:10.1086/282613 |
| [13] | Gong J, Song W B, Warren A. Periphytic ciliate colonization:annual cycle and responses to environmental conditions. Aquatic Microbial Ecology, 2005, 39(2): 159–170. |
| [14] | Xu Y P, Zhang W, Liu Y Y, Xu H L. Identification of potential surrogates to determine functional parameters of periphytic ciliate colonization for bioassessment in coastal waters. Ecological Indicators, 2014, 46: 438–446. DOI:10.1016/j.ecolind.2014.07.017 |
| [15] | 沈韫芬, 龚循矩, 顾曼如. 用PFU原生动物群落进行生物监测的研究. 水生生物学报, 1985, 9(4): 299–308. |
| [16] | 沈韫芬, 章宗涉, 龚循矩, 顾曼如, 施之新, 魏印心. 微型生物监测新技术. 北京: 中国建筑工业出版社, 1990. |
| [17] | 许木启, 曹宏, 王玉龙. 原生动物群落多样性变化与汉沽稳定塘水质净化效能相互关系的研究. 生态学报, 2000, 20(2): 283–287. |
| [18] | 许木启, 朱江, 曹宏. 白洋淀原生动物群落多样性变化与水质关系研究. 生态学报, 2001, 21(7): 1114–1120. |
| [19] | 杨金鹏, 姜勇, 胡晓钟. 青岛沿岸水体原生生物群落与水质状况的关系. 生态学报, 2012, 32(6): 1703–1712. |
| [20] | Xu K D, Choi J K, Yang E J, Lee K C, Lei Y L. Biomonitoring of coastal pollution status using protozoan communities with a modified PFU method. Marine Pollution Bulletin, 2002, 44(9): 877–886. DOI:10.1016/S0025-326X(02)00090-5 |
| [21] | Xu H L, Zhang W, Jiang Y, Min G S, Choi J K. An approach to identifying potential surrogates of periphytic ciliate communities for monitoring water quality of coastal waters. Ecological Indicators, 2011, 11(5): 1228–1234. DOI:10.1016/j.ecolind.2010.12.025 |
| [22] | Xu H L, Zhang W, Jiang Y, Zhu M Z, Al-Rasheid K A S. Influence of sampling sufficiency on biodiversity analysis of microperiphyton communities for marine bioassessment. Environmental Science and Pollution Research, 2012, 19(2): 540–549. DOI:10.1007/s11356-011-0578-1 |
| [23] | 王丽卿, 郝瑞娟, 李雪琪, 潘宏博. 六种淡水纤毛虫形态学研究. 水生生物学报, 2016, 40(2): 343–349. |
| [24] | Wilbert N. Eine verbesserte Technik der Protargolimprägnation für Ciliaten. Mikrokosmos, 1975, 64: 171–179. |
| [25] | Foissner W, Berger H, Kohmann F. Taxonomische und ökologische revision der ciliaten des saprobiensystems, Band Ⅱ:Peritrichia, Heterotrichida, Odontostomatida. Müchen:Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft, 1992. |
| [26] | Foissner W, Berger H, Blatterer H, Kohmann F. Taxonomische und ökologische Revision der Ciliaten des saprobiensystems, Band Ⅳ:Gymnostomatida, Loxodes, Suctoria. Müchen:Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft, 1995. |
| [27] | Foissner W, Berger H, Schaumburg J. Identification and ecology of limnetic plankton ciliates. München:Informationsberichte des Bayerischen Landesamtes für Wasserwirtschaft, 1999. |
| [28] | 吴利, 冯伟松, 陈相瑞, 沈韫芬. 五种淡水吸管虫原生动物中国新纪录种的描述. 动物分类学报, 2006, 31(2): 311–316. |
| [29] | 张武昌, 丰美萍, 于莹, 张翠霞, 肖天. 砂壳纤毛虫图谱. 北京: 科学出版社, 2012. |
| [30] | 国家环境保护总局. 水和废水监测分析方法.第四版. 北京: 中国环境科学出版社, 2002. |
| [31] | 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13–15, 19. |
| [32] | Wilhm J L. Use of biomass units in Shannon's Formula. Ecology, 1968, 49(1): 153–156. DOI:10.2307/1933573 |
| [33] | Margalef R. Information theory in ecology. International Journal of General Systems, 1958, 3: 36–71. |
| [34] | Pielou E C. Ecological Diversity. New York: John Wiley and Sons, 1975: 1–165. |
| [35] | 张伟. 青岛沿海周丛生纤毛虫群落时空特征及其与环境关系的研究[D]. 青岛: 中国海洋大学, 2015. |
| [36] | Agamaliev F G. Ciliates of the solid surface overgrowth of the Caspian Sea. Acta Protozool, 1974, 13: 53–83. |
| [37] | 李继秋, 林晓凤, 许恒龙, 宋微波. 海水养殖水体周丛纤毛虫的群落结构及其对环境因子的响应. 水生生物学报, 2008, 32(4): 493–500. |
| [38] | Jiang J G, Wu S G, Shen Y F. Effects of seasonal succession and water pollution on the protozoan community structure in an eutrophic lake. Chemosphere, 2007, 66(3): 523–532. DOI:10.1016/j.chemosphere.2006.05.042 |
| [39] | 沈韫芬. 原生动物学. 北京: 科学出版社, 1999. |
| [40] | Šimek K, Bobková J, Macek M, Nedoma J, Psenner R. Ciliategrazing on picoplankton in a eutrophic reservoir during the summer phytoplankton maximum:a study at the species and community level. Limnology and Oceanography, 1995, 40(6): 1077–1090. |
| [41] | 郑小燕, 王丽卿. 淀山湖原生动物种类组成及数量变动. 生态学杂志, 2009, 28(1): 51–57. |