 2018, Vol. 38
2018, Vol. 38文章信息
- 陈秋阳, 赵彬洁, 袁洁, 张健, 谭香, 张全发.
- CHEN Qiuyang, ZHAO Binjie, YUAN Jie, ZHANG Jian, TAN Xiang, ZHANG Quanfa.
- 人工遮光和营养添加对河流反硝化活性和反硝化细菌群落结构的影响
- Influences of artificial shading and nutrient addition on denitrification activity and community structure of denitrifying bacteria in a stream
- 生态学报. 2018, 38(15): 5566-5576
- Acta Ecologica Sinica. 2018, 38(15): 5566-5576
- http://dx.doi.org/10.5846/stxb201704050582
-
文章历史
- 收稿日期: 2017-04-05
- 修订日期: 2018-01-30

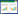




2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
反硝化是去除河流氮污染、调节氮循环的关键过程。通过大气氮沉降、氮肥的使用、粪便的排放和生物固氮等方式输入河流的氮元素, 在最终汇入海洋前, 有将近一半由反硝化的作用去除, 释放到大气中[1]。河流中的氮循环微生物参与了包括固氮、硝化、反硝化、厌氧氨氧化和氨化等生态过程, 各形态的氮作为主要反应物, 河流中的氮素水平直接影响了微生物的群落特征和功能作用, 因此施肥行为将显著改变氮循环微生物的群落结构, 增强其生物功能活性。研究表明, 在河流砂质沉积物内添加氮质营养能促进反硝化进程[2];氮肥的施用促进了反硝化细菌的活性, 有机肥还能提高土壤反硝化细菌多样性[3]。光照的强度对底栖和浮游藻类的生长具有显著的影响[4], 从而影响藻类对河流沉积物中氮磷营养盐和有机碳的吸收和释放[5-7]。比如, 暗光环境会抑制沉积物中异养细菌的生长, 低光照强度能降低异养细菌的碳生物量贡献率[8]。
迄今为止, 反硝化细菌群落对营养响应的生态试验常见于农业土壤和水产养殖沿岸土壤的研究中[9-10], 利用野外原位生态试验探索营养添加和遮光处理对河流沉积物反硝化微生物多样性影响的研究鲜有报道。本实验选择汉江上游金水河流域中的森林溪流, 采集进行原位营养添加和遮光处理后的河流砂质沉积物, 选用nirS功能基因作分子标记, 采用16S rDNA技术研究两种处理对反硝化细菌群落结构和物种多样性的影响, 可以为深入理解人类活动对流域脱氮能力的改变提供依据。
1 实验方法 1.1 研究区域控制实验于2015年10月至2015年12月在陕西省秦岭南坡的佛坪县境内金水河流域进行。金水河是汉江上游区域的一级支流, 发源于佛坪县, 在洋县的黄金峡峡口汇入汉江, 河流长度87 km, 流域面积730 km2, 全流域无工业区分布, 中下游两岸土地被开垦为农田。金水河流域处于北亚热带山地暖温带湿润季风气候区, 多年平均气温11.5—14.5℃, 降水量在900—1244 mm之间。实验样地位于金水河流域上游的佛坪国家自然保护区的核心区域内, 人为干扰小, 森林覆盖率高(>95%), 其中竹林面积占44.7%[11]。金水河的河水水质较好, 呈弱碱性, 离子含量较低, 水化学类型为Ca2+-HCO3-型, 河水中离子主要是受碳酸盐岩和硅酸盐岩风化的控制[12]。
在金水河上游的东河及其支流中选取6条溪流作为实验样地, 分布如图 1, 自上游到下游分别为李家沟、东河、倒流水沟、牌坊沟、瓦房沟和转弯沟。

|
| 图 1 实验样地分布图 Fig. 1 Map of the study area and sampling sites |
控制实验设置在有连续水域的溪流河段上, 水面无树木遮挡。每条溪流选取长20 m、宽8 m的河段, 自上游至下游分成4个区域:对照区(O:Open)、遮光区(S: Shaded)、营养区(ON: Open & Nutrient)和遮光营养区(SN: Shaded & Nutrient)。每个区域尺寸为:长5 m、宽8 m。每个区域之间的间隔为30 m。图 2是对照区和3个处理区的实验布置示意图。对照区(O)河段不进行任何特殊处理, 反映自然状态下的溪流;遮光区(S)在水面以上0.5 m处覆盖长方形遮阳网, 选用遮光率大于95%的黑色遮阳网, 应完全遮盖该区域的水面;在营养区(ON)的沉积物中均匀施加50 kg缓释肥, 缓释肥用小型蛇皮袋分装, 包扎后均匀的投放在河面上;遮光营养区(SN)在沉积物中用相同的方法均匀施加50 kg缓释肥后, 在河面以上50 cm处覆盖同样的黑色遮阳网。由于森林河流在源头处的郁闭度极高, 遮光处理可以模拟河流源头区域通常比较隐蔽光照强度不足的状况;人类沿河耕种施肥和排放生活污水带给河流营养的输入状态可以用营养添加处理来模拟。营养添加处理中使用的缓释肥是美国Scotts公司生产的精准奥绿肥(Osmocote Exact), 有效成分比N:P:K=15:19:12。控制实验的处理时间为60天, 2015年10月布设, 2015年12月进行样品采集。样品采集时, 每一个处理区域选取3个平行河水样品和砂质沉积物样品用于后期实验室内测量反硝化活性和微生物物种多样性。现场测量的指标均由YSI6920水质检测仪完成, 有:河水温度(Temperature)、溶解氧含量(Dissolved Oxygen)、pH和浊度(Turbidity)。总氮(Total Nitrogen;TN)由碳氮仪Multi N/C ®2100测定, 总磷(Total Phosphorus;TP)由钼酸铵分光光度法(GB 11893-89)测得。

|
| 图 2 河流遮盖和营养加富实验布置示意图 Fig. 2 The experiment diagram of coverage and enrichment treatments |
营养添加及河面遮盖2个月后, 取每条溪流四个区域的沉积物。用采样袋采集200 g砂质沉积物, 放入干冰和保温箱中带回实验室, 冻干后用于DNA的提取。用采样袋采集500 g砂质沉积物, 在4℃条件下带回实验室, 测定反硝化活性, 即, 利用乙炔抑制法培养沉积物4 h, 用真空气瓶收集产生的N2O气体, N2O浓度用装有63Ni电子捕获器(Electron Capture Detector)的气相色谱仪测定。每个样品3个重复。评价沉积物的反硝化活性的指标是培养沉积物样品4 h内N2O生成速率, 根据Terry等提供的公式计算水中溶解N2O的浓度[13]。4 h内的平均反硝化速率v为:
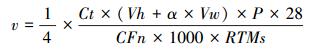
|
单位是:μg N2O-N g-1 h-1, 表示每1 g烘干沉积泥沙在4 h内产生的N2O态N的质量。其中Ct是4 h后顶空气体中N2O的浓度;Vh是顶空气体的体积;Vw是加入的水的体积;α为本森系数;CFn是修正系数, 当只抽取一次气体时, CFn的值为1;Ms为烘干后的沉积物质量;T为培养时的温度;P为压强;R=8.31451 KPa mol-1K-1。
1.4 细菌总DNA提取和PCR扩增选取倒流水沟和东河两条溪流的各区域河流砂质沉积物进行反硝化微生物分析, 分析流程如下:
其中, 使用Omega Soil kit试剂盒从河流沉积物样品中分离出DNA;用NanoDrop分光光度计检验提取的DNA纯度;用Qubit®2.0荧光定量仪测量DNA浓度;利用琼脂糖凝胶电泳检测DNA完整性。
测序区域:反硝化细菌16SrDNA的Cd3aF-R3cd;引物:Cd3aF(5′-GTSAACGTSAAGGARACSGG-3′)和R3cd(5′-GASTTCGGRTGSGTCTTGA-3′)。PCR扩增采用TransStart® Fast pfu DNA Polymerase(20 μl反应体系)。
1.5 序列分析OTU(Operational Taxonomic Unit)及物种群落分析:使用Uparse软件(Uparse v7.0.1001)对所有样品的全部有效标记(Effective Tags)序列进行一致性(Identity)聚类, 形成OTU。在对OTU注释并去除污染OTU后, 使用R软件进行各个分类等级的注释比例和物种相对丰度的统计。选取关注细菌属的OTU使用QIIME软件进行系统进化树构建, 进行系统进化关系展示。最后使用R软件进行纲级别的相对丰度热图绘制, 同时进行样本间和物种间的聚类分析。
Chao指数是常用的通过预测样品中OTU数量, 衡量样品中OTU丰富度的指数, 用来评估沉积物样本的物种多样性, 指数越高, 表明样本的多样性越复杂。覆盖百分率(Coverage)通过下列公式计算:
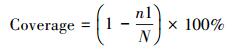
|
式中, n1表示克隆文库中只出现一次的OTU数量, N表示每个克隆文库的克隆总数。覆盖百分率越高, 表明文库表达真实环境中nirS基因群落结构的置信度越高。在文库库容相同的情况下, 覆盖百分率越高, nirS型反硝化细菌的群落多样性就越低。
1.6 统计分析多样性指数(Chao), 使用QIIME软件计算。
沉积物反硝化活性在遮光和营养添加处理下的差异显著性分析, 以及遮光和营养添加处理下沉积物nirS型反硝化细菌物种相对丰度和物种数量的差异显著性分析, 由SPSS 16.0进行方差分析(ANOVA)和LSD多重比较分析方法。
2 结果 2.1 河流水体的基本理化性质控制实验实施前, 6条溪流河水的理化指标背景值如表 1所示。6条溪流的河水均为弱碱性, 溶解氧含量较高, 浊度很低。当四种处理持续2个月时(2015年12月), 营养添加处理前后水体理化性质的改变可对比表 2和表 3。2个月的处理后, 较对照区域(O), 营养添加处理区域(ON)的河水酸碱度显著(P < 0.05)降低, 溶解氧含量显著(P < 0.05)上升。总氮和总磷含量在6条河流中的变化各有不同。较对照区而言, 营养添加仅使倒流水(DLSG)和牌坊沟(PFG)的总氮TN显著(P < 0.05)增加, 对其他河流无显著影响。较对照区而言, 营养添加仅使李家沟(LJG)、倒流水沟(DLSG)、牌坊沟(PFG)和转弯沟(ZWG)的总磷TP显著(P < 0.05)增加, 对其他河流无显著影响。
| 对照区 Open |
溶解氧 Dissolved oxygen/(mg/L) |
pH | 浊度 Turbidity/NTU |
总氮 Total nitrogen/(mg/L) |
总磷 Total phosphorus/(mg/L) |
| 东河DH | 6.46±0.07 | 7.01± < 0.01 | 0.6± < 0.01 | 0.80±0.17 | 0.015± < 0.01 |
| 李家沟LJG | 7.95±0.21 | 7.08±0.11 | 0.1± < 0.01 | 0.79±0.17 | 0.015±0.01 |
| 倒流水DLSG | 9.55±0.10 | 7.42± < 0.01 | 0.2± < 0.01 | 0.81±0.06 | 0.016± < 0.01 |
| 牌坊沟PFG | 8.40±0.02 | 7.13±0.01 | 0.2± < 0.01 | 0.68±0.04 | 0.015±0.01 |
| 瓦房沟WFG | 9.98±0.08 | 7.23± < 0.01 | 0.2± < 0.01 | 1.54±0.11 | 0.012± < 0.01 |
| 转弯沟ZWG | 8.65±0.16 | 7.56± < 0.01 | 0.3± < 0.01 | 0.83±0.05 | 0.010± < 0.01 |
| 第一列缩写为对应的拼音首字母缩写, 表中数值为:平均值±标准偏差, 重复次n=3;NTU为Nephelometric Turbidity Unit, 是散射比浊测定法测定浊度的单位 | |||||
| 对照区 Open |
溶解氧 Dissolved oxygen/(mg/L) |
pH | 浊度 Turbidity/NTU |
总氮 Total Nitrogen/(mg/L) |
总磷 Total Phosphorus/(mg/L) |
| 东河DH | 9.33±0.04a | 7.71± < 0.01a | 0.6± < 0.01a | 0.96± < 0.01a | 0.031±0.01a |
| 李家沟LJG | 7.13±1.16a | 8.22±0.37a | 0.7±0.07a | 0.79±0.21a | 0.027±0.01a |
| 倒流水沟DLSG | 9.02±0.03a | 7.71±0.01a | 0.5±0.07a | 1.06±0.08a | 0.03± < 0.01a |
| 牌坊沟PFG | 8.31±0.05a | 8.11±0.01a | 1.2± < 0.01a | 0.74±0.08a | 0.027±0.01a |
| 瓦房沟WFG | 8.37±0.03a | 7.31± < 0.01a | 0.5± < 0.01a | 1.47±0.12a | 0.013± < 0.01a |
| 转弯沟ZWG | 13.59±0.22a | 7.71±0.04a | 0.5±0.19a | 0.72±0.04a | 0.021± < 0.01a |
| 表中数值为:平均值±标准偏差(Mean±SD), 重复次n=3;小写字母a和b表示在P<0.05水平上的差异;NTU为Nephelometric Turbidity Unit, 是散射比浊测定法测定浊度的单位 | |||||
| 营养区 Open & Nutrient |
溶解氧 Dissolved oxygen/(mg/L) |
pH | 浊度 Turbidity/NTU |
总氮 Total Nitrogen/(mg/L) |
总磷 Total Phosphorus/(mg/L) |
| 东河DH | 9.81±0.02 b | 7.40± 0.01 b | 0.4± < 0.01 b | 0.93±0.05 a | 0.026± < 0.01 a |
| 李家沟LJG | 9.08±0.02 b | 7.13±0.01 b | 0.3±0.07 b | 0.64±0.01 a | 0.046± < 0.01 b |
| 倒流水沟DLSG | 10.64±0.01 b | 7.32±0.01 b | 0.9± < 0.01 b | 1.61±0.05 b | 0.093±0.01 b |
| 牌坊沟PFG | 9.35±0.02 b | 7.5± < 0.01 b | 0.6± < 0.01 b | 0.88±0.02 b | 0.049± < 0.01 b |
| 瓦房沟WFG | 9.80± 0.06 b | 7.27± 0.01 b | 0.6± < 0.01 a | 1.34±0.01 a | 0.012± < 0.01 a |
| 转弯沟ZWG | 14.42±0.08 b | 7.53±0.02 b | 0.2± < 0.01 b | 0.79±0.05 a | 0.053± < 0.01 b |
| 表中数值为:平均值±标准偏差(Mean±SD), 重复次数n=3;小写字母a和b表示在P<0.05水平上的差异;NTU为Nephelometric Turbidity Unit, 是散射比浊测定法测定浊度的单位 | |||||
对4种处理下6条河流的沉积物4 h平均反硝化速率做单因素方差分析和LSD法多重比较分析, 结果表明, 遮光处理S与对照O、营养添加ON和遮光且营养添加SN这3种处理之间的差异性显著(P<0.05), O、ON和SN 3种处理之间彼此没有显著差异(P>0.05)。如图 3所示是在6条河流中4种处理下的河流沉积物反硝化活性。相比于对照区(O), 河面遮光处理(S)显著(P<0.05)降低了沉积物的反硝化活性, 营养添加处理(ON)和营养添加且河面遮盖处理(SN)的沉积物反硝化活性略微增加, 但与对照区(O)没有显著的差异(P>0.05)。均进行了遮光处理的S和SN两组之间比较, 遮光且营养添加处理(SN)显著(P<0.05)提高了沉积物的反硝化活性。

|
| 图 3 不同处理下的河流沉积物培养4 h内的反硝化速率 Fig. 3 Denitrification rate of 4-hour-inhibited sediments in different segments 纵坐标反硝化速率的单位ng N2O-N g-1 h-1, 表示每1 g烘干沉积泥沙在4 h内产生的N2O态N的质量;O:对照区(Open), S:遮光区(Shading), ON:营养区(Open & Nutrient), SN:遮光营养区(Shading & Nutrient);a和b表示小写字母表示在P < 0.05水平上的差异 |
见表 4, 6条河流中, 经过S、ON和SN 3种处理后, 倒流水沟的沉积物反硝化活性变化幅度最大, 4 h内平均反硝化速率在河面遮盖区域达到最小值15.98 ng N2O-N g-1 h-1, 在营养添加区域达到最大值114.90 ng N2O-N g-1 h-1。牌坊沟的沉积物反硝化活性变化幅度最小, 4 h内平均反硝化速率在河面遮盖区域达到最小值63.15 ng N2O-N g-1 h-1, 在营养添加区域达到最大值77.70 ng N2O-N g-1 h-1。
| 处理 Treatments |
李家沟LJG | 倒流水沟DLSG | 牌坊沟PFG | 东河DH | 瓦房沟WFG | 转弯沟ZWG |
| O | 70.19±18.75 | 90.51±32.60 | 71.27±23.09 | 75.67±14.16 | 73.60±9.77 | 87.25±28.20 |
| S | 24.98±8.18 | 15.98±2.360 | 63.15±18.92 | 29.02±3.22 | 41.14±12.64 | 18.18±10.33 |
| ON | 72.76±30.58 | 114.90±30.61 | 77.70±25.81 | 98.84±18.77 | 85.60±20.01 | 89.91±12.19 |
| SN | 60.80±10.79 | 86.09±15.86 | 75.49±25.52 | 87.17±30.28 | 70.57±29.52 | 71.83±20.74 |
| O:对照区, Open; S:遮光区, Shading; ON:营养区, Open & Nutrient; SN:遮光营养区, Shading & Nutrient; 表中数值为:平均值±标准偏差(Mean±SD), 重复次数n= 3 | ||||||
反硝化细菌的序列分析是基于东河(DH)和倒流水沟(DL)中四种处理区域的沉积物样品进行的。基于样本内的OTU的物种系统进化关系及相对丰度信息, 在属水平上, 选择每个分类中所有样品相对丰度最大的前10个分类, 分别是Alpha变形菌纲的趋磁螺菌属(Magnetospirillum), Beta变形菌纲的未分类Beta变形菌(Candidatus_ Accumulibacter_phosphatis)、纤毛菌属(Leptothrix、Rubrivivax)、固氮螺菌属(Azospira)、脱氯单胞杆菌属(Dechloromonas)和冻胶菌属(Zoogloea), Gamma变形菌纲的假单胞菌属(Pseudomonas)和黄杆菌属(Rhodanobacter), 除此之外的反硝化细菌序列均属于非变形菌门的环境样品(environmental_samples_norank)。在可识别的反硝化细菌中, 脱氯单胞杆菌属具有最高的相对丰度, 占所有测序nirS型反硝化细菌的15.476%, 远高于其他属, 在控制实验四种处理区域内的沉积物中均为优势属(图 4)。

|
| 图 4 4种处理区域内沉积物中nirS型反硝化细菌在各分类学分支的分布 Fig. 4 Taxonomic distribution of denitrifying bacteria by encoding nirS gene in sediments from 4 segments 扇形的面积表示不同处理区域内的样品在该细菌分类上相对丰度的比例大小;分类名下方的数字表示4种处理区域种的所有沉积物样品在该分类上的平均相对丰度百分率 |
对东河与倒流水沟采用营养添加、河面遮盖、营养添加且河面遮盖这3种处理后, 河流沉积物反硝化微生物的群落产生了较为明显的变化。相比于对照区的样点, 3种处理点的细菌在沉积物中所有nirS型细菌的总相对丰度均有变化, 在河面遮盖点的降幅约为14.29%±1.21%, 在营养添加点的降幅约为9.22%±0.83%, 在营养添加且河面遮盖的点降幅为10.66%±0.98%。此外, 细菌的物种组成和各物种丰度也发生了相应的改变。在东河及其倒流水沟的沉积物中, 优势属均为脱氯单胞杆菌属, 在各支流沉积物反硝化细菌中的相对丰度在19.46%—74.99%之间。3种处理方式均降低了脱氯单胞杆菌在沉积物中nirS型反硝化细菌里的相对丰度。将3种处理点和控制点对比, 河面遮盖的处理非常显著(P<0.001)增加了假单胞菌属的物种数量和相对丰度, 显著(P<0.05)降低了固氮螺菌属和脱氯单胞杆菌属的物种数量和物种丰度;营养添加的处理非常显著(P<0.001)增加了假单胞菌的物种丰度, 显著(P<0.05)降低了脱氯单胞杆菌的物种丰度。
根据所有样品在纲水平的物种注释及丰度信息绘制热图, 对样品进行聚类分析。图 5中横向为样品信息, 纵向为物种注释信息, 图中左侧的聚类树为物种聚类树, 上方的聚类树为样品聚类树, 中间热图对应的值为每一行物种相对丰度经过标准化处理后得到的标准化值(http://en.wikipedia.org/wiki/Standard_score), 即, 一个样品在该分类上的相对丰度和所有样品在该分类的平均相对丰度的差除以所有样品在该分类上的标准差所得到的值。黄色柱形高度代表标准化值相对大小。在此丰度聚类分析中, 东河的控制区域(DH.O)和营养添加区域(DH.ON)沉积物反硝化细菌聚为一类, 河面遮盖区(DH.S)与营养添加且河面遮盖区域(DH.SN)分属两类;倒流水沟的控制区域(DL.O)和营养添加区域(DL.ON)沉积物反硝化细菌聚为一类, 河面遮盖区(DL.S)与营养添加且河面遮盖区域(DL.SN)聚为一类。

|
| 图 5 东河与倒流水沟各处理区沉积物nirS反硝化细菌在纲级别的丰度聚类 Fig. 5 Class-level clustering of denitrifying bacteria by encoding nirS gene in sediments from all segments in DH and DL streams DL.O:倒流水沟对照区, Dao Liushui Open;DL.S:倒流水沟遮光区, Dao Liushui Shaded;DL.ON:倒流水沟营养区, Dao Liushui Open and Nutrient;DL.SN:倒流水沟遮光营养区, Dao Liushui Shaded and Nutrient; DH.O:东河对照区, Dong He Open;DH.S:东河遮光区, Dong He Shaded;DH.ON:东河营养区, Dong He Open and Nutrient;DH.SN:东河遮光营养区, Dong He Shaded and Nutrient |
表 5第二列为反硝化细菌群落OTU种类划分, 可以看出倒流水沟(DL)各处理状态下沉积物的划分的OTU种类均低于同处理状态下的东河(DH)各区域。本次实验样品文库覆盖率均达到78%以上, 说明能够较为真实的反映出测序结果的真实情况。在倒流水沟的四个处理中, 控制区域的沉积物反硝化细菌的Chao指数最大, 即物种多样性最大, 营养添加区域Chao指数最小, 即物种多样性最小;在东河的4个处理中, 控制区域的沉积物反硝化细菌物种多样性最大, 营养添加区域物种多样性最小。
| 处理 Treatment |
OTU种类 OTUs |
Chao指数 Chao Index |
文库覆盖度 Coverage/% |
| DL.O | 589 | 713.56 | 79.38 |
| DL.S | 649 | 680.54 | 85.39 |
| DL.ON | 505 | 524.5 | 88.28 |
| DL.SN | 594 | 632.01 | 78.05 |
| DH.O | 934 | 1107.31 | 88.27 |
| DH.S | 814 | 936.31 | 93.46 |
| DH.ON | 655 | 728.23 | 87.80 |
| DH.SN | 845 | 920.87 | 90.96 |
| DL.O:倒流水沟对照区, Dao Liushui Open;DL.S:倒流水沟遮光区, Dao Liushui Shaded;DL.ON:倒流水沟营养区, Dao Liushui Open and Nutrient;DL.SN:倒流水沟遮光营养区, Dao Liushui Shaded and Nutrient; DH.O:东河对照区, Dong He Open;DH.S:东河遮光区, Dong He Shaded;DH.ON:东河营养区, Dong He Open and Nutrient;DH.SN:东河遮光营养区, Dong He Shaded and Nutrient | |||
反硝化还原酶编码基因作为反硝化微生物的功能基因, 常用来筛选和检测环境中反硝化微生物。反硝化微生物从有机化合物中获得碳源和能量, 并将其作为电子供体, 将电子传递给氮氧化物, 完全反应过程为:NO3-→NO2-→NO→N2O→N2。其中, 亚硝酸盐还原是指在亚硝酸盐还原酶(Nitrite reductase, Nir)的作用下亚硝酸盐被还原成气态中间产物NO, 该过程被认为是反硝化作用中重要的限速步骤, 因此两种形态的亚硝酸盐还原酶对应的编码基因(nirS、nirK)被作为功能基因最早地出现在有关反硝化细菌的分子生态学研究中[14]。大多数反硝化作用的研究都集中在反硝化细菌上, 反硝化细菌被认为是自然反硝化过程的主要承担者[15], 然而真菌、放线菌、酵母菌和深海孔虫的反硝化作用也逐渐被发现[16]。目前已经发现的反硝化细菌约有50个属, 主要集中在变形菌纲(Proteobacteria)和拟杆菌纲(Bacteroidetes)。但实验室培养和鉴定出的反硝化细菌种类有限, 因此数据库中能用于鉴定反硝化作用的功能基因序列较少, 即使是被广泛研究的nirS和nirK基因, 扩增人工设计的引物之后也只能识别出变形菌门的部分属, 大多数环境样品研究中产生的序列片段均来自于未知物种[17]。
森林溪流沉积物中可检测出的10个主要nirS型反硝化细菌属中, 好氧反硝化细菌的相对丰度对水体溶解氧变化的响应十分显著。假单胞菌属和冻胶菌属是好氧反硝化细菌, 其反硝化能力受环境中溶解氧的限制较少。在人工培养环境下, 一种假单胞菌属的菌株(Stutzeri SU2)即使在浓度达到92%的O2氛围中, 也能在92 h的培养后, 实现99.24%的硝态氮去除率[18]。在本次控制实验的四种处理区域中, 营养区缓释肥的加入导致水体溶解氧含量的上升, 同时营养区的假单胞菌属和冻胶菌属的相对丰度均显著的高于对照区;遮光处理抑制了水体中光合作用的进行, 使得遮盖区溶解氧含量较对照区降低, 同时假单胞菌属和冻胶菌属的相对丰度在遮光区也显著低于对照区。因此, 在实验区溪流水体中较高的溶解氧含量促进了沉积物中好氧反硝化细菌的广泛分布。此外, 兼性异养反硝化细菌也会受到溪流水体中溶解氧升高的影响。Beta变形菌纲的红长命菌属是兼性异养细菌, 它在实验区溪流沉积物样品中的相对丰度仅次于脱氯单胞杆菌属。红长命菌可以利用无机氮氧化物进行光合作用, 但它不可直接利用硝酸盐, 只能将亚硝酸盐还原成N2, 因此红长命菌只能在厌氧和低氧条件下进行反硝化作用[19]。在营养添加和遮光这两种单独处理下的沉积物中, 红长命菌属的相对丰度均显著高于对照区, 但遮光且营养区的红长命菌属的相对丰度与对照区比不具备显著性, 说明遮光和营养添加增加红长命菌属的相对丰度的效应不会累加。
反硝化细菌的除磷效应会影响反硝化细菌对水体营养荷载的响应方式。好氧反硝化细菌在进行反硝化时, 一般还具有除磷的作用。实验区沉积物内检测出的10种主要nirS型反硝化细菌广泛具有除磷效应。除好氧细菌(假单胞菌属和冻胶菌属)以外, Beta变形菌纲红环菌目(Rhodocyclales)中的脱氯单胞杆菌属和固氮螺菌属也具有除磷的作用, 是工业除磷活性污泥中常用的兼性反硝化除磷菌 [20]。红环菌目的各类细菌普遍具有除磷作用, 是水污染处理中脱磷除氮系统的主要菌种之一, 且对无机磷营养的添加有正向响应, 研究结果表示磷矿石的添加会促进红环菌目细菌优势菌群的数量, 从而提高水体硝酸盐的去除效果[21]。然而一些研究也表明, 活性污泥系统中的好氧反硝化菌对磷营养盐增加的响应不明显, 而污泥中的溶解氧含量、碳源含量和碳源种类均显著影响除磷效果, 但对反硝化细菌除磷作用的影响机制较为复杂, 不同种类的细菌在不同溶氧环境下对各类有机碳源会选择性吸收[22-24]。脱氯单胞杆菌属在实验区沉积物中是相对丰度最高的是优势菌属。在四种处理区域内, 无机营养盐的添加处理对脱氯单胞杆菌的丰度无显著影响, 遮光处理却显著降低了脱氯单胞杆菌的丰度。遮光后水体溶解氧减少, 藻类和光合细菌的化能合成受阻, 水体有机碳含量减少, 这验证了溶解氧和有机碳的变化会对除磷菌的反硝化作用有影响。
在不同环境下反硝化细菌群落结构的差异很大, 因此在不同自然环境下, 营养添加处理会对反硝化微生物群落和物种多样性产生不同的影响。例如在水稻种植土壤中, nirS型反硝化细菌对仅仅施加无机营养的反应不显著, 而有机营养和无机营养同时施加比仅仅添加有机营养更能够显著提高反硝化细菌的种群丰度[25-27]。在控制实验中施加NPK为主成分的缓释肥, 极显著地降低了沉积物中优势菌属(脱氯单胞杆菌属)的相对丰度, 增加了假单胞菌属的相对丰度, 且促进了沉积物反硝化活性。
营养添加和遮光处理对河流沉积物反硝化细菌的影响有助于理解不同土地利用类型的河岸带反硝化作用的差异。研究发现, 农田和草地河岸带土壤中的反硝化酶活性低于林地河岸带, 主要是因为农田和草地河岸带上的人类活动更频繁, 促进了土壤中的全氮和有机碳的损失[28]。虽然河口湿地潮滩有机碳源充足, 而有植被覆盖的潮滩土壤反硝化活性大于无植被覆盖区域[29]。由此可见, 河岸带覆盖植被的差异对沉积物反硝化过程影响的途径之一是改变环境中的可利用的碳源和氮源。
控制实验中光照条件的变化也会影响水体沉积物中反硝化细菌可利用的氮源和碳源。光能自养生物膜是指在陆地和水生环境表面形成的由光能驱动的微生物群落, 其中, 富氧光养微生物(如底栖硅藻、蓝藻和底栖绿藻)进行光合作用产生能量, 为生物膜上的异养微生物提供有机碳和氧[30]。生物膜上好氧的硝化细菌利用产生的有机碳和氧将水体里的氨态氮转化为硝酸盐, 为反硝化细菌提供了可用无机氮。当光照不充足时(遮光处理或者夜间), 生物膜内缺氧, 厌氧或兼性厌氧的反硝化细菌利用生物膜内产生的有机碳和硝酸盐, 进行反硝化作用[31]。当光照强度升高, 光能自养生物膜上合成的有机碳很快被同系统的异养反硝化细菌利用, 反硝化效率增高[32]。遮盖处理极大降低光照强度, 光合作用的缺乏导致生物膜内层出现相对缺氧的状态, 好氧菌的反硝化作用被抑制, 仅厌氧菌或兼性菌具有反硝化能力。若长期缺乏光照, 初级生产者藻类的光合作用降低, 从而合成有机碳的减少, 异养菌和兼性菌的反硝化能力也会随之下降。因此, 在控制实验中, 2个月的遮光处理导致水体中藻类和光合细菌化能合成作用能力下降, 溶解氧的缺乏抑制了好氧反硝化细菌的活性, 有机碳的缺乏抑制了异养和兼性厌氧反硝化细菌的生长, 造成了遮光处理下沉积物反硝化活性的显著降低。而在营养添加且遮光(SN)区, 添加的营养盐缓解了河水中长期缺乏营养的状况, 因而沉积物的反硝化能力恢复, 与控制区无显著差异。
4 结论(1) 营养添加和河面遮盖对选取的6条实验河流中沉积物的反硝化活性产生了相似的影响:营养添加处理会增加沉积物的反硝化活性, 遮盖处理会降低沉积物的反硝化活性。
(2) 在东河与倒流水沟各处理区域的沉积物样品中, 优势属均为脱氯单胞杆菌属。河面遮盖和营养添加这两种处理方式均降低了脱氯单胞杆菌属的相对丰度。河面遮盖的处理显著提高了假单胞菌属的物种数量和相对丰度, 固氮螺菌属和脱氯单胞菌的物种数量和物种丰度显著减少;营养添加的处理后, 假单胞菌属的相对丰度与物种数量显著增加, 脱氯单胞杆菌的相对丰度显著减少。
(3) 在东河与倒流水沟各处理区域中, NPK营养的添加和河面遮盖均降低了nirS型反硝化细菌的物种多样性, 营养添加处理后的河流沉积物中nirS型反硝化细菌的物种多样性是各处理区域中最低的。
| [1] | Galloway J N, Aber J D, Erisman J W, Seitzinger S P, Howarth R W, Cowling E B, Cosby B J. The nitrogen cascade. BioScience, 2003, 53(4): 341–356. DOI:10.1641/0006-3568(2003)053[0341:TNC]2.0.CO;2 |
| [2] | Magalhães C M, Joye S B, Moreira R M, Wiebe W J, Bordalo A A. Effect of salinity and inorganic nitrogen concentrations on nitrification and denitrification rates in intertidal sediments and rocky biofilms of the Douro River estuary, Portugal. Water Research, 2005, 39(9): 1783–1794. DOI:10.1016/j.watres.2005.03.008 |
| [3] | 陈哲, 袁红朝, 吴金水, 魏文学. 长期施肥制度对稻田土壤反硝化细菌群落活性和结构的影响. 生态学报, 2009, 29(11): 5923–5929. DOI:10.3321/j.issn:1000-0933.2009.11.021 |
| [4] | 庄树宏, Sven H. 光强和光质对底栖藻类群落影响Ⅱ.群落和种群的动态和适应模式. 生态学报, 2001, 21(12): 2057–2066. DOI:10.3321/j.issn:1000-0933.2001.12.014 |
| [5] | 姚扬, 金相灿, 姜霞, 李丽和. 光照对湖泊沉积物磷释放及磷形态变化的影响研究. 环境科学研究, 2004, 17(S1): 30–33. |
| [6] | 王琦. 浅水湖泊沉积物磷释放的生物学机制研究[D]. 杨凌: 西北农林科技大学, 2006. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y977673 |
| [7] | 操家顺, 单淇. 光照强度对光能自养生物膜去除低污染水体中硝态氮的影响研究. 环境污染与防治, 2013, 35(3): 15–19. |
| [8] | 余立华. 秋季长江口不同辐照和氮、磷浓度水平下浮游植物营养盐吸收动力学及生长变化研究[D]. 上海: 华东师范大学, 2006. |
| [9] | Wakelin S A, Nelson P N, Armour J D, Rasiah V, Colloff M J. Bacterial community structure and denitrifier (nir-gene) abundance in soil water and groundwater beneath agricultural land in tropical North Queensland, Australia. Soil Research, 2011, 49(1): 65–76. DOI:10.1071/SR10055 |
| [10] | Krishnani K K. Detection and diversity of nitrifying and denitrifying functional genes in coastal aquaculture. Aquaculture, 2010, 302(1/2): 57–70. |
| [11] | 王婧, 袁洁, 谭香, 李思悦, 张全发. 汉江上游金水河悬浮物及水体碳氮稳定同位素组成特征. 生态学报, 2015, 35(22): 7338–7346. |
| [12] | 蒋保刚. 秦岭南坡山区小流域水化学及氢氧同位素研究——以金水河流域为例[D]. 保定: 河北大学, 2013. |
| [13] | Terry R E, Tate Ⅲ R L, Duxbury J M. The effect of flooding on nitrous oxide emissions from an organic soil. Soil Science, 1981, 132(3): 228–232. DOI:10.1097/00010694-198109000-00006 |
| [14] | Braker G, Zhou J Z, Wu L Y, Devol A H, Tiedje J M. Nitrite reductase genes (nirK and nirS) as functional markers to investigate diversity of denitrifying bacteria in pacific northwest marine sediment communities. Applied and Environmental Microbiology, 2000, 66(5): 2096–2104. DOI:10.1128/AEM.66.5.2096-2104.2000 |
| [15] | Lu H J, Chandran K, Stensel D. Microbial ecology of denitrification in biological wastewater treatment. Water Research, 2014, 64: 237–254. DOI:10.1016/j.watres.2014.06.042 |
| [16] | Risgaard-Petersen N, Langezaal A M, Ingvardsen S, Schmid M C, Jetten M S M, den Camp H J M O, Derksen J W M, Piña-Ochoa E, Eriksson S P, Nielsen L P, Revsbech N P, Cedhagen T, van der Zwaan G J. Evidence for complete denitrification in a benthic foraminifer. Nature, 2006, 443(7107): 93–96. DOI:10.1038/nature05070 |
| [17] | 杨柳燕, 王楚楚, 孙旭, 郭丽芸, 肖琳, 宋晓骏. 淡水湖泊微生物硝化反硝化过程与影响因素研究. 水资源保护, 2016, 32(1): 12–22, 50-50. |
| [18] | Su J J, Liu B Y, Liu C Y. Comparison of aerobic denitrification under high oxygen atmosphere by Thiosphaera pantotropha ATCC 35512 and Pseudomonas stutzeri SU2 newly isolated from the activated sludge of a piggery wastewater treatment system. Journal of Applied Microbiology, 2001, 90(3): 457–462. DOI:10.1046/j.1365-2672.2001.01265.x |
| [19] | Nagashima S, Kamimura A, Shimizu T, Nakamura-Isaki S, Aono E, Sakamoto K, Ichikawa N, Nakazawa H, Sekine M, Yamazaki S, Fujita N, Shimada K, Hanada S, Nagashima K V P. Complete genome sequence of phototrophic betaproteobacterium Rubrivivax gelatinosus IL144. Journal of Bacteriology, 2012, 194(13): 3541–3542. DOI:10.1128/JB.00511-12 |
| [20] | 吕小梅. 反硝化除磷菌群结构与工艺调控策略[D]. 哈尔滨: 哈尔滨工业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10213-1014081360.htm |
| [21] | 张建美, 郝春博, 冯传平, 李思远, 张丽娜, 郝会玲, 朱江. 磷对地下水反硝化系统中细菌菌群结构的影响. 地学前缘, 2014, 21(4): 191–198. |
| [22] | 李相昆, 张杰, 黄荣新, 马力, 鲍林林, 姜安玺. 反硝化聚磷菌的脱氮除磷特性研究. 中国给水排水, 2006, 22(3): 35–39. |
| [23] | 于大禹, 郭威, 张金榜, 张贺, 杨鹏. 好氧反硝化菌的筛选及其脱氮除磷性质的研究. 微生物学通报, 2009, 36(4): 598–603. |
| [24] | 姜丽丽. 生物膜污水处理系统脱氮除磷性能及微生物群落结构研究[D]. 邯郸: 河北工程大学, 2012. http://doi.wanfangdata.com.cn/10.7666/d.y2132741 |
| [25] | Yin C, Fan F L, Song A L, Li Z J, Yu W T, Liang Y C. Different denitrification potential of aquic brown soil in Northeast China under inorganic and organic fertilization accompanied by distinct changes of nirS-and nirK-denitrifying bacterial community. European Journal of Soil Biology, 2014, 65: 47–56. DOI:10.1016/j.ejsobi.2014.09.003 |
| [26] | 罗希茜, 陈哲, 胡荣桂, 吴敏娜, 秦红灵, 魏文学. 长期施用氮肥对水稻土亚硝酸还原酶基因多样性的影响. 环境科学, 2010, 31(2): 423–430. |
| [27] | 尹昌, 范分良, 李兆君, 宋阿琳, 朱平, 彭畅, 梁永超. 长期施用有机和无机肥对黑土nirS型反硝化菌种群结构和丰度的影响. 环境科学, 2012, 33(11): 3967–3975. |
| [28] | 陈刚亮, 李建华, 杨长明. 崇明岛不同土地利用类型河岸带土壤反硝化酶活性特征. 应用生态学报, 2013, 24(10): 2926–2932. |
| [29] | 游丽丽. 河口潮滩湿地植被对沉积物反硝化过程影响初探[D]. 上海: 华东师范大学, 2014. |
| [30] | Roeselers G, M C M Van Loosdrecht, G Muyzer. Phototrophic biofilms and their potential applications. Journal of applied phycology, 2008, 20(3): 227–235. DOI:10.1007/s10811-007-9223-2 |
| [31] | 操家顺, 白王军, 王超, 王海生, 张永兴. 光能自养生物膜形成机制及脱氮影响因素的研究进展. 环境污染与防治, 2011, 33(1): 78–82. |
| [32] | 邓旭, 魏斌, 胡章立. 利用固定化藻菌耦合系统同步去除污水中的COD和氮磷. 环境科学, 2011, 32(8): 2311–2316. |