 2018, Vol. 38
2018, Vol. 38文章信息
- 杨振奇, 秦富仓, 张晓娜, 李晓琴, 牛晓乐, 刘力川.
- YANG Zhenqi, QIN Fucang, ZHANG Xiaona, LI Xiaoqin, NIU Xiaole, LIU Lichuan.
- 砒砂岩区不同立地类型人工沙棘林下草本物种多样性环境解释
- Environmental interpretation of herb species diversity under different site types of Hippophae rhamnoides forest in feldspathic sandstone region
- 生态学报. 2018, 38(14): 5132-5140
- Acta Ecologica Sinica. 2018, 38(14): 5132-5140
- http://dx.doi.org/10.5846/stxb201707231320
-
文章历史
- 收稿日期: 2017-07-23
- 网络出版日期: 2018-04-04






物种多样性是指生态系统中存在着不同种类的生物, 即物种水平上的生物多样性[1]。物种多样性是生物在特定的时空范围内进化的重要产物, 表现了生物对环境的适应性, 强调了物种的变异性。物种多样性随生物栖息环境的梯度变化而表现出一定差异规律, 运用单一的描述统计方法和简单的线性数学方法在探讨植被物种随环境梯度变化上还存在着较大的局限性[2], 近年来应用对应分析、去趋势对应分析以及典范对应分析等方法来定量分析植被与环境的关系已逐步成为植被生态学的主要研究手段[3], 探明生境与植物种多样性的关系, 并籍此提高人工植被生态系统自我修复能力和抵御外界干扰能力是现阶段我国人工植被建设工作的重点内容之一。林下草本层是人工林生态系统的基础功能层[4], 李民义等[5]研究显示, 人工林下草本层物种多样性与人工林森林生态水文功能存在密切联系。陈彩虹等[6]研究表明, 林下草本层物种多样性对于森林土壤理化性质有明显改良作用。李国雷等[7]则从人工林生长发育规律角度阐述了人工林下草本物种多样性的价值, 林下草本层物种多样性常作为衡量人工林生态功能恢复的关键指标[8]。
砒砂岩区分布在黄河上中游段晋陕蒙交界三角地带, 砒砂岩成岩程度低, 颗粒间胶结程度差, 在水力、风力交互作用下极易发生风化剥蚀, 使得这一地区成为黄土高原区侵蚀最剧烈的区域。该区自20世纪80年代末大面积营造人工林, 对当地植被恢复、生态环境调控, 地方经济发展都起到了一定程度促进作用。沙棘作为治理砒砂岩区水土流失的关键树种, 其减水减沙、固持水土方面能力一直是专家学者研究的热点, 但是从生物多样性角度入手, 分析砒砂岩区环境对沙棘林下草本植被影响的理论和成果尚属空白。研究该区沙棘人工林下草本群落物种多样性及其与环境的关系, 是深入研究砒砂岩区人工林生态系统功能变化规律并科学解读砒砂岩区植被恢复机理的有效途径, 对于促使植被恢复过程向理想方向发展具有重要参考价值。本文以砒砂岩区不同立地类型人工沙棘林下草本植物为研究对象, 通过方差分析、典范对应分析等方法, 探讨其生物多样性指数与环境的关系, 定量的揭示砒砂岩区立地条件对沙棘林下草本物种多样性的影响, 以期为砒砂岩区植被建设从生态功能恢复角度上选择适宜立地条件提供理论依据。
1 材料与方法 1.1 研究区概况研究区鲍家沟流域位于鄂尔多斯市准格尔旗西部暖水乡内, 地理坐标为110°31′—110°35′E, 39°46′—39°48′N, 海拔在1110—1300 m之间;流域地形北高南低;属温带大陆性气候, 冬季漫长干燥, 夏季短暂温热, 年均气温7.2 ℃, 年降雨量400 mm;土壤类型以黄绵土、风沙土为主。主要乔木有:青杨(Populus cathayana)、旱柳(Salix matsudana)、油松(Pinus tabuliformis)、侧柏(Platycladus orientalis)等;灌木有柠条(Caragana korshinskii)、沙棘(Hippophae rhamnoides)、山杏(Prunus armeniaca)等;草本植物主要有羊草(Leymus chinensis)、猪毛菜(Salsola nitraria)、阿尔泰狗娃花(Heteropappus altaicus)等。
1.2 研究方法 1.2.1 样地设置与采样于2016年7月在研究区内按照不同立地类型和不同配置模式进行人工沙棘林(林龄相同)的样地布设, 其中峁顶和沟底各1个, 阴坡和阳坡各2个(均为坡中), 对照样地天然草地1个, 共设置7个大样地(沙棘纯林4个和油松+沙棘混交林2个), 样地分布见图 1, 样地基本情况见表 1。详细记录大样地的高程、海拔、坡度、坡向。每个大样地内布设3个15 m×15 m灌木样地, 对样地内林木进行每木检尺, 并在每个灌木样地四角及中心各布设1个1 m×1 m草样方, 共计105个草样方, 测量草样方内植物种类、数量、高度、盖度等指标。清除表层植被和枯落物后开挖土壤剖面, 记录土层厚度、土壤质地等指标, 用铝盒和环刀等工具分别按0—20, 20—40, 40—60 cm采集土样, 进行风干处理备用。

|
| 图 1 研究区样地分布 Fig. 1 Distribution of sample plot |
| 样地编号 Sample number |
立地类型 Site types |
混交结构 Mixed structure |
海拔 Elevation/m |
坡度 Slope/° |
坡向 Slope aspect |
| 1 | 峁顶 | 灌木纯林 | 1260 | 5 | — |
| 2 | 阴坡 | 灌木纯林 | 1228 | 20 | EN |
| 3 | 阴坡 | 乔灌混交林 | 1221 | 18 | EN |
| 4 | 阳坡 | 灌木纯林 | 1202 | 21 | ES |
| 5 | 阳坡 | 乔灌混交林 | 1207 | 27 | ES |
| 6 | 沟底 | 灌木纯林 | 1155 | 2 | — |
| 7 | 沟底 | 天然草地 | 1155 | 2 | — |
重要值=(相对密度+相对盖度+相对频度)/3
植被群落多样性指数采用Shannon-Wiener多样性指数、Simpson优势度指数、Pielou均匀度指数进行分析, 其公式如下:
(1) Shannon-Wiener多样性指数:H=∑PilnPi
(2) Simpson优势度指数:SP=1-∑Pi2
(3) Pielou均匀度指数:Jsw=H/lnS
(4) Patrick指数:S
式中: Pi=Ni/N, Ni为样方中第i种植物的重要值;N为样方植物重要值总和, S为物种数。
土壤含水量采用烘干法, 土壤容重采用环刀法, 土壤pH采用pH计法, 土壤有机质采用重铬酸钾外加热法。
生物因子中是否混交设置为二进制形式, 0为非混交结构, 1为混交结构。
地形因子中将坡向方位角转换为0—1之间的TRASP坡向指数, 公式如下[9]:
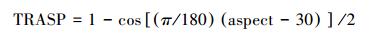
|
运用Canoco 5.0软件对10个环境和生物因子与105个样地草本植物重要值先后采取DCA、CCA、PCCA排序的方法, 进行物种与环境梯度关系的分析, 单因素方差分析和LSD多重比较运用SPSS完成, 数据可视化在Orgin 9.0下完成。
2 结果分析 2.1 不同立地类型人工沙棘林下植物种类组成本次研究调查7种立地类型人工沙棘林下草本层, 共发现草本植物种51种, 隶属于12科36属, 其中禾本科和菊科均含15种以上物种, 占研究区所调查植物种数59%以上。表 2列出了人工沙棘林下草本层植物种重要值排名前3的主要植物种, 可以看出研究区人工沙棘林下草本生活型主要以半灌木和多年生草本为主, 植物水分生态类型以旱生和中旱生为主, 半灌木优势种主要以茭蒿(Artemisia leucophylla)、百里香(Thymus serpyllum var. mongolicus)、铁杆蒿(Artemisia sacrorum)为代表, 多年生草本优势种以本氏针茅(Stipa bungeana)、假苇拂子茅(Calamagrostis pseudaphragmites)为主, 其他伴生种主要有白草(Pennisetum centrasiaticum)、赖草(Leymus secalinus)、阿尔泰狗娃花(Heteropappus altaicus)、硬质早熟禾(Poa sphondylodes)等。
| 样地 Sample |
植物种 Specices |
重要值 Important value |
植物种 Specices |
重要值 Important value |
植物种 Specices |
重要值 Important value |
| 1 | 百里香Thymus serpyllum var. mongolicus | 27.17 | 本氏针茅Stipa bungeana | 13.31 | 达乌里胡枝子 Lespedeza davurica |
11.21 |
| 2 | 铁杆蒿Artemisia sacrorum | 17.67 | 赖草Leymus secalinus | 15.44 | 白草 Pennisetum centrasiaticum |
11.98 |
| 3 | 茭蒿Artemisia leucophylla | 15.41 | 本氏针茅Stipa bungeana | 12.49 | 阿尔泰狗娃花 Heteropappus altaicus |
12.24 |
| 4 | 冷蒿Artemisia frigida | 20.16 | 糙隐子草 Cleistogenes squarrosa |
11.71 | 达乌里胡枝子 Lespedeza davurica |
11.16 |
| 5 | 本氏针茅Stipa bungeana | 25.31 | 达乌里胡枝子 Lespedeza davurica |
17.44 | 白草 Pennisetum centrasiaticum |
15.21 |
| 6 | 茭蒿Artemisia leucophylla | 21.42 | 铁杆蒿Artemisia sacrorum | 12.34 | 硬质早熟禾 Poa sphondylodes |
10.67 |
| 7 | 假苇拂子茅 Calamagrostis pseudaphragmites |
21.14 | 艾蒿Artemisia argyi | 17.74 | 硬质早熟禾 Poa sphondylodes |
13.17 |
Patrick指数能真实且客观的反映生境中物种数目的多寡, 由图 2可以看出, 不同立地类型沙棘林下草本Patrick指数存在较大差异, 其中阴坡混交林下Patrick指数最多为23, 阴坡纯林次之为20, 峁顶纯林最少为10, 小于对照天然草地(14)。阴坡纯林与阳坡纯林Patrick指数差异显著, 沟底纯林与峁顶纯林Patrick指数差异显著, 沟底纯林与阴坡纯林Patrick指数差异不显著, 表明阴坡和沟底的立地条件能满足多数植物生长需求, 其物种相较阳坡和峁顶生境更丰富;阴坡混交林与阴坡纯林物种数差异显著, 而阳坡混交林数与阳坡纯林物种没有显著差异, 表明在水热条件适宜植物生长的情况下, 采取混交的造林方式能提高沙棘林下物种丰富度。Pielou指数反映了生境中物种个体分配的均匀程度, 从图 2可以看出, 阳坡混交林Pielou指数最高为0.89, 沟底纯林Pielou指数最低为0.76。阴坡纯林与阳坡纯林、沟底纯林与峁顶纯林、阴坡混交林与阳坡混交林Pielou指数差异均不显著, 而坡面与沟底和峁顶差异显著。这反映出, 各立地类型虽然物种丰富度有一定差异, 但坡面物种分配的均匀性和群落结构的稳定性要高于其他立地类型。图 2的Shannon指数反映了个体出现的不确定程度, 不同立地条件下沙棘林下Shannon指数大小依次为阴坡纯林(2.60)=阴坡混交林(2.60)>阳坡混交林(2.58)>阳坡纯林(2.49)>天然草地(2.32)>沟底纯林(2.24)>峁顶纯林(2.06), 坡面与其他立地类型Shannon指数差异显著, 说明坡面相较其他立地条件物种出现的不确定程度更高, 同时在物种分配均匀性接近的情况下, 沟底物种出现的不确定性明显高于峁顶。图 2的Simpson指数反映了生境物种多样性的综合优势度, 不同立地类型下沙棘林下物种多样性的综合优势度的大小依次为阴坡混交林>阴坡纯林>阳坡混交林>沟底纯林>阳坡混交林>峁顶纯林>天然草地。

|
| 图 2 不同立地类型人工沙棘林下草本植物种多样性 Fig. 2 Herb species diversity under different types of Hippophae rhamnoides forest 图中相同字母代表差异显著(a=0.05水平下);样地1为峁顶纯林, 样地2为阴坡纯林, 样地3为阴坡混交林, 样地4为阳坡纯林, 样地5为阳坡混交林, 样地6为沟底纯林, 样地7为天然草地 |
从表 3中可以看出, 各物种多样性指数与环境因子表现出不同程度相关性, Patrick指数与郁闭度显著正相关, Pielou指数与坡度和土层厚度显著正相关, Shannon指数与坡向显著正相关, Simpson指数与郁闭度、含水量均显著正相关。这表明沙棘林郁闭度对林下草本物种丰富度影响较大, 坡度和土层厚度主要与物种分配的均匀性有关, 坡向影响着物种出现的不确定性, 土壤含水量和沙棘林郁闭度对物种多样性的综合优势度有较大影响。
| 指数 Index |
坡向 Slope aspect |
海拔 Elevation/m |
混交结构 Mixed structure |
郁闭度 Canopy density/% |
土壤容重 Soil bulk density |
pH | 土壤有机质 Soil organic carbon/(g/kg) |
土层厚度 Soil thickness/m |
坡度 Slope/(°) |
土壤含水量 Soil water content/g |
| Patrick指数Patrick Index | -0.589 | -0.084 | 0.538 | 0.743* | -0.110 | -0.312 | 0.026 | -0.402 | 0.397 | 0.519 |
| Pielou指数Pielou Index | 0.105 | 0.263 | 0.499 | 0.210 | -0.098 | 0.042 | 0.393 | 0.823* | 0.733* | 0.302 |
| Shannon指数Shannon Index | 0.780* | -0.109 | 0.529 | 0.622 | -0.359 | 0.088 | 0.134 | 0.616 | 0.450 | 0.589 |
| Simpson指数Simpson Index | -0.292 | 0.270 | 0.495 | 0.819* | -0.297 | 0.065 | 0.010 | 0.672 | 0.346 | 0.736* |
| *表示显著相关(a=0.05) | ||||||||||
从表 4中可知, DCA结果显示四个排序轴的梯度长度介于3—4之间, 可以采用单峰模型排序。CCA环境排序轴与物种相关系数均在0.88以上, 表现出良好相关性。第1、2轴的特征值分别0.431, 0.327, 0.296, 0.215。四轴累计方差解释比为79.7%, 能较为完整的反映排序信息, 前两轴累计方差解释比47.6%, 表明前两轴所包含信息占已有排序信息的绝大部分。
| 因子Factor | 第一轴Axis1 | 第二轴Axis2 | 第三轴Axis3 | 第四轴Axis4 |
| 土壤含水量Soil water content/g | -0.8332** | -0.2521 | 0.1126 | 0.3745 |
| 土壤pH Soil pH | -0.272 | 0.1179 | 0.0398 | -0.1411 |
| 郁闭度Canopy density/% | -0.1436 | 0.4384* | 0.1276 | -0.5491* |
| 土壤容重Soil bulk density | -0.1267 | -0.3741 | -0.5008* | -0.4442 |
| 土壤有机质Soil organic carbon/(g/kg) | 0.0242 | 0.5128* | -0.0251 | 0.1285 |
| 海拔Elevation/m | 0.1576 | -0.3452 | -0.2949 | 0.3671 |
| 坡度Slope/° | 0.2136 | -0.7324** | 0.4888* | 0.003 |
| 土层厚度Soil thickness/m | 0.3287 | -0.6307* | 0.5667* | -0.0511 |
| 混交结构Mixed structure | 0.448* | 0.4266* | 0.1448 | -0.4679* |
| 坡向Slope aspect | 0.7658** | -0.0379 | 0.3937 | -0.0971 |
| Dca | 3.151 | 1.629 | 1.187 | 1.790 |
| Cca | 0.431 | 0.327 | 0.296 | 0.215 |
| 相关系数Correlation coefficient | 0.954 | 0.946 | 0.917 | 0.883 |
| 累计方差贡献率/% Cumulative percentage variance |
27.1 | 47.6 | 66.2 | 79.7 |
| *表示显著相关, * *表示极显著相关(a=0.05) | ||||
环境因子与第一排序轴相关性按相关系数(绝对值)的大小依次为土壤含水量、坡向、混交结构、土层厚度、土壤pH、坡度、海拔、郁闭度、土壤容重、土壤有机质, 其中土壤含水量与第一排序轴呈极显著负相关, 坡向与之呈极显著正相关。环境因子与第二排序轴相关性大小顺序为坡度>土层厚度>土壤有机质>郁闭度>混交结构>土壤容重>海拔>土壤含水量>土壤pH>坡向, 坡度与第二排序轴呈极显著负相关, 土层厚度与之呈显著负相关, 混交结构, 郁闭度, 土壤有机质与之呈显著正相关。
结合表 4和图 3可以看出, 第一排序轴反映了生境的干旱程度对物种多样性的影响, 从左至右含水量逐渐减小, Trasp值逐渐增加, 生境从湿润转向干旱。第二排序轴主要从坡度和土层厚度方面反映了生境的差异, 图 3样地间的距离反映了样地的相似程度, 可以看出阳坡纯林与阳坡混交林生境较为接近, 阴坡纯林与阴坡混交林较为接近, 沟底和峁顶生境差异较大。由图 3可以看出, 随生境的梯度变化, 物种分布也随之改变, 沿着排序轴物种变化幅度较大, 如42号狼毒(Stellera chamaeiasme)和43号假苇拂子茅, 与其他物种距离较远, 表明其对环境要求较为严苛, 其中假苇拂子茅更趋向于生长在水分充足有季节性流水的沟底, 而狼毒更倾向生长在较为干旱的峁顶。20号小画眉草(Eragrostis minor)等一年生草本物种间距离较少, 且集中在原点, 表明其对环境要求较低, 适生境更为广泛。

|
| 图 3 样地(物种)与环境因子的CCA排序图 Fig. 3 CCA ordination diagram of sample plots (species) and environmental factors |
在此基础上, 采用PCCA变量分离的方法来分析地形因子, 土壤因子, 和灌木层因子对沙棘林下草本物种多样性的影响, 手动检验每组变量中显著的环境因子, 并用蒙特卡洛法检验植被变化的显著性。由表 5可以看出, 所选三类环境因子可解释惯量占总惯量的80%以上, 且与第一排序轴相关性均达极显著水平, 由可解释量大小可知, 环境因子对物种的影响顺序依次为地形因子>土壤因子>灌木层因子。
| 环境变量 Factor |
可解释惯量 Variance explain |
占总惯量比 Variance explain ratio |
交互解释量 Share variance explain |
第一轴 First axis |
所有轴 All axis |
| 土壤因子Soil factor | 0.880 | 26.47 | 0.174 | 0.005** | — |
| 地形因子Ferrain factor | 1.063 | 35.58 | 0.001** | 0.004** | |
| 灌木层因子Shrub layer factor | 0.674 | 19.74 | 0.009** | 0.071 | |
| 注:*表示显著相关, * *表示极显著相关(a=0.05) | |||||
物种多样性具有丰富性和平衡性两方面的内容[10], Partick指数常被用来从丰富度视角对生境物种多样性进行描述[11], 本文研究表明, 阴坡地形和混交结构林分下物种丰富度最高, 这与周萍[12]等研究结果一致。Hillebrand等[13]认为物种均匀度的提高对于提升群落结构稳定性有积极影响, Pielou指数表现出坡面高于其他立地条件, 说明坡面生境相比沟底和峁顶生境群落在平衡性方面表现更为突出, 草本层群落结构也更为稳定。Shannon-weaner指数来源于信息论中计算熵的公式, 在均匀度差异不显著的情况下, 其对物种丰富度较为敏感[1], 本文研究显示, 沟底纯林与峁顶纯林的均匀度接近, 但沟底纯林Shannon指数为2.24显著高于峁顶纯林2.05, 也说明沟底物种出现稀有种的可能性更大, 稀有种对于Shannon指数的计算存在较大贡献[14], 实际调查情况也证实沟底生境由于其季节性流水的缘故, 生长着其他生境所不具有的芦苇(Phragmites australis)等湿生植物。本文研究发现, 不同立地条件下的沙棘林下草本物种Simpson指数没有显著差异(a=0.05), 呈现物种多样性指数不随物种丰富度变化而变化的现象, 许晴等[15]提出的Simpson指数的稀释效应很好的解释了这一现象。
CCA排序第一轴从地形角度描述了砒砂岩区水热条件对于物种分布的影响, 第二轴反映了坡度以及砒砂岩区独有的土壤性质方面差异对于物种分布的影响, 本文所选环境因子和生物因子对环境解释量基本已经涵盖了影响砒砂岩区人工沙棘林下草本物种多样性的因素, 其中地形因子解释量占有较大比重。土壤含水量虽然与物种多样性指数表现出较高的相关性, 其在第一排序轴中也处于主导地位, 但土壤因子中还包含了土壤pH、土壤有机质等与物种多样性相关性较弱的因子, 因此其所占解释量要低于地形因子。相比地形和土壤因子, 灌木层因子不具备明显的梯度特征, 因此其解释量低于二者。交互解释部分也说明了, 三类因子存在交叉影响部分, 特别是地形因子在很大程度上影响着土壤性质和植被生长, 未能解释量部分可能包含了人为干扰因素, 如造林前的整地工作和造林后期的抚育管理等因素[16-17]。
物种多样性的差异是环境、生物等因子多方面协同影响下的结果, 研究尺度的不同, 影响因素也有所区别。冯建孟等[18]认为在空间大尺度水平上, 气候是制约植物种多样性差异的首要因素。许传阳等[19]认为地形是导致中等空间尺度植物多样性垂直格局变异的关键因素。杨阳和刘秉儒[20]认为土壤理化性质差异是引起小尺度范围内植物种多样性变化的主要因素。砒砂岩区典型小流域气候条件基本一致, 其地形属性有着典型黄土丘陵沟壑区的破碎化地形特征, 同时又独有砒砂岩裸露, 土层浅薄的地域特点, 与张振国等[21]结论不同的是, 土壤因子并非影响小尺度生境物种多样性的唯一因子, 本文研究区地形高差200 m, 海拔影响作用相对而言并不突出, 而坡向和坡度对砒砂岩地区的水热条件起着重新分配的作用[22], 这与朱云云等[23]的研究结果一致。本文研究证实, 砒砂岩区人工沙棘林下物种多样性与地形因素、土壤性质以及生物因素相关性均较大, 土壤因素中土壤含水量无论是对群落物种多样性还是对物种个体的空间分布均有较大影响, 值得强调的是, 土层厚度是砒砂岩区土壤性质中较为独特的指标, 在实地调查中发现该区部分区域土层厚度小于5 cm, 甚至部分地区地表砒砂岩风化物裸露, 砒砂岩的存在对于土壤持水能力有着较大影响[24], 间接影响着生物多样性的变化。生物因素中, 沙棘配置模式和栽植密度对于林下草本物种多样性均有不同程度的影响[25], 林分的混交结构对于林下植物种多样性均有一定程度的影响[26], 研究中发现, 沙棘林的郁闭度与物种多样性指数呈显著相关, 混交结构与物种多样性指数相关性虽不显著, 但是其与每个物种多样性指数的相关性更为稳定。综上所述, 砒砂岩区沙棘林下物种多样性的差异是地形、土壤和灌木层因素影响的综合体现。
4 结论(1) 砒砂岩区人工沙棘林下共发现草本植物种51种, 以旱生和中旱生为主, 大部分为禾本科和菊科植物种。
(2) 沙棘林下草本物种多样性随立地条件的变化表现出不同规律, 阴坡和沟底相对于其他立地类型物种更为丰富, 沙棘混交林下物种丰富度高于纯林;坡面物种分配更加均匀, 群落结构更加稳定;在物种分配均匀性接近的情况下, 沟底物种出现的不确定性最大;阴坡混交林物种多样性综合优势度最高。
(3) 环境因子从不同角度影响着沙棘林下草本物种多样性, 沙棘林郁闭度对物种丰富度影响较大, 坡度和土层厚度对物种分配的均匀度影响较大, 坡向则影响着物种出现的不确定程度, 土壤含水量和沙棘林郁闭度则对物种多样性的综合优势度有影响。
(4) 草本物种的分布展现出较强的水热规律, 地形因素中, 坡向和坡度是影响沙棘林下草本物种分布的主要因素;土壤因素中土壤含水量和土层厚度是影响沙棘林下草本物种分布的关键因素;土层厚度是砒砂岩区影响草本物种分布的特有因素;灌木层因素中郁闭度和混交结构是影响沙棘林下草本物种分布的次要因素。
针对砒砂岩区地形和土壤结构特点, 建议在坡面和沟谷地带营造沙棘混交林, 有助于提高林下草本生物多样性和林分稳定性, 高效发挥沙棘恢复植被、固持水土的生态功能。
| [1] | 张金屯. 数量生态学.第二版. 北京: 科学出版社, 2011: 83–105. |
| [2] | 张峰, 张金屯. 我国植被数量分类和排序研究进展. 山西大学学报:自然科学版, 2000, 23(3): 278–282. |
| [3] | 郑超超, 伊力塔, 张超, 余树全, 库伟鹏, 钱逸凡, 凌骅. 浙江江山公益林物种种间关系及CCA排序. 生态学报, 2015, 35(22): 7511–7521. |
| [4] | 范玉龙, 胡楠, 丁圣彦, 翟元杰, 柳静, 廖秉华, 卢训令. 伏牛山自然保护区森林生态系统草本植物功能群的分类. 生态学报, 2008(07): 3092–3101. |
| [5] | 李民义, 张建军, 郭宝妮, 黄明, 茹豪. 晋西黄土区不同密度油松人工林林下植物多样性及水文效应. 生态学杂志, 2013, 32(5): 1083–1090. |
| [6] | 陈彩虹, 田大伦, 方晰, 田蜜. 城郊4种人工林林下植被物种多样性、生物量与土壤养分相关性. 水土保持学报, 2010, 24(6): 213–217. |
| [7] | 李国雷, 刘勇, 于海群, 吕瑞恒, 李瑞生. 油松(Pinus tabulaeformis)人工林林下植被发育对油松生长节律的响应. 生态学报, 2009, 29(3): 1264–1275. |
| [8] | 高阳, 程积民, 赵钰, 苏纪帅. 黄土区典型人工林草本层生态恢复效应. 草地学报, 2013, 21(1): 79–86. DOI:10.11733/j.issn.1007-0435.2013.01.012 |
| [9] | 尤业明, 徐佳玉, 蔡道雄, 刘世荣, 朱宏光, 温远光. 广西凭祥不同年龄红椎林林下植物物种多样性及其环境解释. 生态学报, 2016, 36(1): 164–172. |
| [10] | Wilsey B J, Potvin C. Biodiversity and ecosystem functioning:importance of species evenness in an old field. Ecology, 2000, 81(4): 887–892. DOI:10.1890/0012-9658(2000)081[0887:BAEFIO]2.0.CO;2 |
| [11] | 董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472–1483. |
| [12] | 周萍, 刘国彬, 侯喜禄. 黄土丘陵区不同坡向及坡位草本群落生物量及多样性研究. 中国水土保持科学, 2009, 7(1): 67–73. |
| [13] | Hillebrand H, Bennett D M, Cadotte M W. Consequences of dominance:a review of evenness effects on local and regional ecosystem processes. Ecology, 2008, 89(6): 1510–1520. DOI:10.1890/07-1053.1 |
| [14] | 王晶, 焦燕, 任一平, 薛莹, 纪毓鹏, 徐宾铎. Shannon-Wiener多样性指数两种计算方法的比较研究. 水产学报, 2015, 39(8): 1257–1263. |
| [15] | 许晴, 张放, 许中旗, 贾彦龙, 尤建民. Simpson指数和Shannon-Wiener指数若干特征的分析及"稀释效应". 草业科学, 2011, 28(4): 527–531. |
| [16] | 王顺霞, 武高林, 王占军. 宁南山区不同造林整地方式对草地植物群落特征的影响. 草业科学, 2009, 26(12): 87–91. |
| [17] | 段劼, 马履一, 贾黎明, 贾忠奎, 公宁宁, 车文瑞. 抚育间伐对侧柏人工林及林下植被生长的影响. 生态学报, 2010, 30(6): 1431–1441. |
| [18] | 冯建孟, 徐成东. 中国种子植物物种丰富度的大尺度分布格局及其与地理因子的关系. 生态环境学报, 2009, 18(1): 249–254. |
| [19] | 许传阳, 陈志超, 郝成元, 丁效东. 暖温带和北亚热带过渡区白云山植物物种多样性与环境因子相关性. 生态环境学报, 2014, 23(3): 371–376. |
| [20] | 杨阳, 刘秉儒. 宁夏荒漠草原不同群落生物多样性与生物量关系及影响因子分析. 草业学报, 2015, 24(10): 48–57. DOI:10.11686/cyxb2014438 |
| [21] | 张振国, 焦菊英, 贾燕锋, 王宁. 黄土丘陵沟壑区不同立地环境因子对植被变化的解释比例分析. 中国水土保持科学, 2010, 8(2): 59–67. |
| [22] | 邱扬, 傅伯杰, 王军, 陈利顶. 黄土丘陵小流域土壤水分的空间异质性及其影响因子. 应用生态学报, 2001, 12(5): 715–720. |
| [23] | 朱云云, 王孝安, 王贤, 邓美皎. 坡向因子对黄土高原草地群落功能多样性的影响. 生态学报, 2016, 36(21): 6823–6833. |
| [24] | 杨振奇, 秦富仓, 李晓琴, 刘力川, 牛晓乐. 砒砂岩区主要造林树种枯落物及林下土壤持水特性. 水土保持学报, 2017, 31(3): 118–122. |
| [25] | 李玉新, 赵忠, 陈金泉. 不同林龄人工沙棘林结构与林下物种多样性研究. 西北植物学报, 2010, 30(4): 776–785. |
| [26] | 曲红, 王百田, 王棣, 高海平. 黄土区不同配置人工林物种多样性研究. 生态环境学报, 2010, 19(4): 843–848. |