 2018, Vol. 38
2018, Vol. 38文章信息
- 赵玉皓, 张艳杰, 严月, 刘玉槐, 徐燕, 刘苑秋, 郭晓敏, 娄翼来, 鲁顺保.
- ZHAO Yuhao, ZHANG Yanjie, YAN Yue, LIU Yuhuai, XU Yan, LIU Yuanqiu, GUO Xiaomin, LOU Yilai, LU Shunbao.
- 亚热带退化红壤区森林恢复类型土壤有机碳矿化对温度的响应
- Response to different temperatures under forest recovery types on soil organic carbon mineralization in subtropics
- 生态学报. 2018, 38(14): 5056-5066
- Acta Ecologica Sinica. 2018, 38(14): 5056-5066
- http://dx.doi.org/10.5846/stxb201707181295
-
文章历史
- 收稿日期: 2017-07-18
- 网络出版日期: 2018-04-04






2. 江西农业大学 林学院, 南昌 330045;
3. 中国农业科学院农业环境与可持续发展研究所, 北京 100081
2. College of Forestry, Jiangxi Agricultural University, Nanchang 330045, China;
3. Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081, China
土壤有机碳是全球陆地生态系统碳循环的重要组成部分, 全球气候变化引起气温升高加速了微生物和动物对土壤有机碳的矿化作用, 其产物通过土壤呼吸作用以CO2形式释放反馈回大气圈, 进一步加剧全球变暖[1], 因此, 土壤呼吸是陆地生态系统土壤和大气之间CO2交换的重要途径, 且对环境变化反应最为敏感[2]。
土壤温度几乎调控着所有的生物地球化学过程, 是土壤呼吸的主要控制因子[3], 土壤呼吸作用的增强和温度的增加存在着潜在的正反馈关系, 最终可能导致全球变暖加剧[4, 5], 因此土壤呼吸是全球变暖反馈环节中主要不确定因素之一[6], 这也是目前全球变化研究的热点问题, 各国学者对此研究异常活跃, 他们正尝试通过量化各种生态系统碳通量与环境因子之间的关系模型来预测未来全球变化对陆地生态系统的影响[7]。影响土壤呼吸作用的因子有很多, 包括土壤环境、土地利用方式以及植被类型等[4]。尽管森林生态系统类型的转变可能对土壤CO2的释放做出巨大贡献, 然而, 目前关于土壤呼吸对不同森林类型的响应的仍存在争议。土壤温度是影响土壤呼吸作用的又一个重要因素, Rodeghlero和Cescatti研究表明土壤矿化和温度之间具有显著的相关关系, 主要是温度影响微生物的活动所致[8], Nakajima等研究表明, 在-5℃到25℃之间, 随着温度的增高, 土壤呼吸速率表现为逐渐增强的趋势[9]。但由于温度的季节性波动使得土壤矿化具有不确定性, 温度对土壤有机碳矿化的影响尚未达成一致共识, 仍有许多争论[10]。因此, 弄清温度对特定森林土壤有机质分解的影响, 对揭示该森林生态系统碳循环至关重要。然而, 在实地测定土壤呼吸时, 容易受到其他环境因素的交互影响, 很难准确的将温度和其他因素对土壤碳矿化的影响区分开来;因此, 室内培养就被广泛地应用于土壤有机质分解的机理性研究[11-13]。目前有关土壤呼吸速率与环境因子之间的关系的报道较多[1e], 但在不同温度条件下不同森林类型林地土壤碳矿化方面的研究较少。
本文以亚热带地区典型的针叶马尾松(Pinus massoniana)、阔叶木荷(Schima superb)和枫香(Liquidambar formosana)林为研究对象, 分别将其土壤进行不同温度的室内培养, 并定期测定其土壤碳矿化速率, 分析讨论以下2个问题:1)植被类型对森林土壤碳矿化及其温度敏感性的影响;2)温度和森林类型对土壤碳矿化的交互影响。其研究结果将对研究林型和温度因素对土壤碳矿化的影响具有重要意义。
1 材料与方法 1.1 研究地区概况该试验地位于江西省中部的泰和县(26°44′N, 115°04′E), 始建于1991年, 属亚热带季风湿润性气候, 年均气温为18.6℃, 年均降水量为1726 mm, 降雨多集中在4—6月份, 大约占全年降水量的49%, 7—9月高温干旱。土壤成土母质是第四纪红粘土, 表层石砾含量较多, 属于典型的红壤低丘岗地。20世纪80年代以前, 由于长期的强度樵采、割茅、挖蔸和过牧等人为干扰, 原生常绿阔叶林已不复存在, 植被覆盖度仅30%, 且分布不均。自1991年起, 选用马尾松、木荷和枫香等树种造林, 建立了面积为133 hm2、多样化植被恢复类型的实验基地, 目前植被覆盖度已达到90%以上, 林相结构完整, 森林小气候已形成, 生态、经济和社会效应良好[15]。
1.2 样品采集采样时间为2015年1月, 在研究区分别随机选取马尾松林、木荷林、枫香林3种植被类型, 每种类型各选择3块大小为20 m×20 m的样方作为重复, 在每个样方中以S型选取5个点, 用7.5 cm土钻在0—10 cm取土壤样品。然后将相同样点的土壤混匀, 将新鲜土壤样品过2 mm筛去除土壤动物、小石头、可见的植物残体和根系等杂质, 每份1 kg, 分成2份装入无菌自封袋, 其中一份用生物冰袋冷藏, 供土壤呼吸作用以及其他一些微生物的测定, 另一部分带回实验室进行pH值、全氮、全碳、有效磷等指标的测定。土壤基本理化性质见表 1。
| 植被类型 Forest type |
pH | 含水量 Moisture/% |
全氮 Total N/% |
全碳 Total C/% |
全磷 Total P/(mg/g) |
| 马尾松Pinus massoniana | 4.35 | 14.18 | 0.102 | 1.405 | 0.217 |
| 木荷Schima superb | 4.34 | 17.74 | 0.222 | 2.689 | 0.281 |
| 枫香Liquidambar formosana | 4.40 | 17.30 | 0.190 | 2.319 | 0.256 |
方法参照Chen等[16], 采用元素分析仪(Isoprime-EuroEA3000, 意大利)测定土壤全碳、全氮含量, DOC和DON含量采用2.0 mol/L KCl浸提, 然后用TOCN分析仪测定SOC和可溶性总氮(total soluble N, TSN), pH值用1:2.5水溶液、pH计测定, 含水量采用烘干法。土壤碳矿化测试方法:称取新鲜土壤40.0 g, 将其含水量调到田间持水量的60%, 放入4个恒温培养箱(温度分别设为5、15、25℃和35℃)里进行35 d培养。土壤碳矿化测定方法采用碱液吸收法:2个70 mL的带盖塑料瓶置于1 L的容器内, 其中一个为CO2吸收装置, 其内加入用于吸收CO2的0.1 mol/L NaOH, 另一个装有40.0 g鲜土, 在22℃条件下进行培养, 同时做5个对照试验(仅放置碱液)。培养第1、3、7、14、21、28天和35天后将盛有0.1 mol/L NaOH吸收瓶取出后, 先加入1.0 mol/L BaCl2使得土壤排放的CO2以碳酸盐的形式沉淀, 加入酚酞指示剂后搅拌, 用0.05 mol/L HCl进行滴定剩余的NaOH。对土壤碳矿化进行方程模拟:
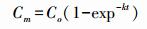
|
式中:Cm为瞬时碳排放量, Co是土壤碳排放量极大值, k为常数, t为培养时间[17], 结合温度敏感性Q10分析, 即温度每升高10℃土壤碳矿化速率所增加的倍数。
1.4 数据分析实验数据运用SPSS 17.0软件分析, 对不同处理间的差异显著性进行One-way(单因素方差分析)进行检验, 用LSD法进行差异显著性检验。采用SigmaPlot 10.0软件制图, 显著性水平设定为α=0.05。
2 结果与分析 2.1 不同温度对土壤CO2累计排放量的影响由图 1看出, 土壤CO2累计排放量随温度升高而增加, 在3种林地中, 土壤CO2累计排放量均表现为15、25、35℃之间差异显著(P < 0.05)。马尾松、木荷和枫香3种林地35℃时土壤CO2累计排放量最高, 分别为246.08、193.70 mg/kg和265.07 mg/kg土壤, 分别相对于25℃提高了44.68%、52.04%和39.21%;25℃时3种林地土壤CO2累计排放量分别为136.13、92.90 mg/kg土壤和161.13 mg/kg土壤, 分别相对于15℃时提高了60.87%、64.30%和58.67%;15℃与5℃之间土壤CO2累计排放量差异则不显著。

|
| 图 1 不同温度下土壤CO2累计排放量(平均值±标准误差) Fig. 1 Effects of substrate addition on CO2-C emission under Chinese fir in the same temperature (mean±SE) 不同小写字母a、b、c表示同一处理不同温度间差异显著(P < 0.05) |
由图 2看出, 4个温度下, 土壤的呼吸速率由高到低均表现为:枫香林﹥马尾松林﹥木荷林, 在25℃条件下, 枫香林和马尾松林地土壤呼吸速率显著高于木荷林地(P < 0.05), 3种林地土壤均表现为第一天呼吸速率最高, 而后均表现为下降趋势, 培养21 d后, 土壤呼吸速率均逐渐趋于平缓。在5℃时土壤呼吸速率表现为第一天最高, 而后骤降, 之后一直保持平缓的状态;在15℃时土壤呼吸速率总体变化差异不大;在25℃和35℃时土壤呼吸速率为第一天最高, 而后缓慢下降, 最后趋于平缓。

|
| 图 2 不同森林类型土壤有机碳矿化速率 Fig. 2 Effects of different forest types on CO2-C evolved in different temperature |
从图 3, 随温度的增加, 不同植被类型的土壤CO2累计排放量呈现规律性的变化。在25℃和35℃条件下, 培养7 d内土壤碳排放量增加明显, 尔后呈缓慢上升趋势, 刚开始进行室内培养实验时, 土壤从冷库中取出, 微生物被激活, 此时活性最大, 随后微生物活性逐渐减弱, 因此后期呼吸速率较为平缓;在5℃与15℃时, 土壤CO2累积排放量则一直表现为缓慢上升的趋势。4个温度条件下均表现为枫香林地土壤CO2累计排放量最大, 其次是马尾松林, 木荷林地土壤CO2累计排放量最小。在5℃时, 枫香林和马尾松林与木荷林之间最大潜在碳排放差异显著, 而马尾松林与木荷林之间差异不显著;在15℃、25℃、35℃条件下, 3种林地土壤最大潜在碳排放差异均不显著(P > 0.05)。由图 3我们可以看出, 在没有干扰的条件下, 土壤呼吸作用会在一定的时间后达到平衡, 保持一个最高值。

|
| 图 3 不同森林类型土壤有机碳累计碳排放(平均值±标准误) Fig. 3 Effects of different forest types on rate of soil respiration (mean±SE) 不同小写字母a和b表示同一处理不同温度间差异显著(P < 0.05) |
在不同温度条件下, 对不同环境因子和土壤累积碳排放量进行回归分析, 得出土壤全碳和土壤含水量与土壤累积碳排放量存在一定的相关性(图 4)。在15℃和25℃条件下, 土壤全碳对土壤碳排放量均呈现抛物线变化规律, 即随土壤全碳含量增加, 土壤碳排放量呈现先增后减的变化趋势(15℃, R2=0.4865, P < 0.1;25℃, R2=0.5512, P < 0.05), 全碳的极值点分别约为1.83%和1.89%。在5℃和35℃条件下, CO2累积排放量随土壤全碳含量并无显著性差异。对不同森林类型土壤碳含水量与碳排放量进行回归分析得出(图 4)在25℃时, 不同林地土壤碳排放量随土壤含水量的增加呈现出先增后减的变化趋势, 土壤含水量的极值点为14.93%, 且差异达显著性水平(P < 0.1), 在其他温度条件下并无显著性差异。在相同的相对含水量的条件下, 土壤碳排放量随温度的升高而增加, 说明土壤含水量并不是影响土壤碳排放量的调控因子。

|
| 图 4 土壤全碳及含水量对不同森林类型土壤碳矿化的影响 Fig. 4 Effects of total carbon and moisture for different forest soil carbon minerlization |
对植被类型和培养温度与土壤碳矿化量进行分析得出, 植被类型和温度对土壤碳矿化有显著影响, 且不同温度对土壤呼吸作用更显著(P < 0.000), 但两因素间并无显著的交互效应(表 2, P>0.05)。对不同森林类型土壤生物及环境因子进行冗余分析, 其结果显示土壤P、MBN和N素与碳矿化之间基本上是呈现出负相关关系(图 5), 因此土壤P、MBN和N素对土壤碳排放有一定的限制作用。
| 组别Terms | P |
| 物种Species | 0.02* |
| 温度Temperature | 0.000*** |
| 物种×温度Species × temperature | 0.37 |
| ***, P < 0.001; *, P < 0.05 | |

|
| 图 5 环境因子和生物因子间的冗余分析排序 Fig. 5 Biplot of redundancy analysis (RDA) of the relationships between environmental factors and microbes metabolic profiles in different forest types soils |
在没有外界干扰的情况下长时间培养后土壤累积碳排放量将会达到一个峰值。对不同林型林地土壤进行35 d的室内培养实验, 运用方程Cm=Co(1-exp-kt)对土壤潜在碳排放进行模拟, 得出不同温度下不同林型林土壤最大碳排放量随温度升高而升高(表 3), 相关系数R2除5℃条件下的马尾松和木荷外, 均在0.9以上, 说明拟合效果很好。在5℃条件下, 枫香林地土壤碳矿化量显著高于马尾松和木荷林(P < 0.05), 在其余3种温度条件下, 枫香和马尾松林地土壤潜在碳排放量显著高于木荷(P < 0.05)。
| 温度 Temperature/℃ |
处理 Treatments |
潜在碳排放量 Potentially carbon mineralizable(Co) |
系数 Constants (K) |
相关系数 Relation coefficients(R2) |
| 5 | 马尾松 | 39.875±7.964b | 0.081±0.035a | 0.829±0.013a |
| 木荷 | 39.758±0.559b | 0.048±0.007b | 0.772±0.029b | |
| 枫香 | 71.121±9.418a | 0.040±0.010b | 0.902±0.009a | |
| 15 | 马尾松 | 74.270±21.288a | 0.041±0.009b | 0.995±0.003a |
| 木荷 | 35.524±5.183b | 0.073±0.005a | 0.991±0.004a | |
| 枫香 | 80.910±14.374a | 0.047±0.002b | 0.990±0.005a | |
| 25 | 马尾松 | 202.917±63.342a | 0.036±0.008b | 0.991±0.002a |
| 木荷 | 113.081±18.772b | 0.052±0.010a | 0.995±0.002a | |
| 枫香 | 200.255±31.662a | 0.048±0.006ab | 0.995±0.000a | |
| 35 | 马尾松 | 298.407±70.212a | 0.050±0.005b | 0.993±0.000a |
| 木荷 | 232.848±30.033b | 0.049±0.004c | 0.999±0.000a | |
| 枫香 | 283.570±12.197a | 0.067±0.007a | 0.983±0.003a | |
| 同一列不同小写字母表示同一温度不同处理间差异显著(P < 0.05), 图中数据为平均值±标准差 | ||||
土壤呼吸与温度的关系还可以从温度敏感性角度来研究(Q10), 因为二氧化碳产生的温度敏感性可作为有机质降解温度敏感性的主要指标。本研究中Q10培养35d的结果见图 6, 其范围为1.797—1.971, 变化幅度较小, 不同森林林型在不同温度条件下的Q10值并未表现出显著性差异。

|
| 图 6 第35天的Q10值 Fig. 6 The Q10 value of the 35th day ns:无显著性差异 |
土壤矿化是一个非常复杂的生物学过程, 受土壤温度、含水量、C、N含量等非生物因子以及植被类型综合影响, 且土壤碳矿化的大部分变化主要是由温度和湿度引起[18-19]。由于本研究区域年均降水量为1726 mm, 说明该地区并不缺水, 并且本研究将土壤含水量调节到持水量的60%, 这时土壤呼吸作用最为活跃。因此本研究主要关注在相对湿度一致的条件下土壤异养呼吸与土壤温度的关系。结果表明在35℃以下范围内随温度升高能显著提升林地土壤碳矿化量, 在较低温度(5℃和15℃), 土壤CO2累计排放量较低, 土壤微生物并没有被激活或激活缓慢。在较高温度(25℃和35℃), 土壤CO2累计排放量较高, 其原因可能是土壤温度升高, 促使土壤中微生物、土壤中酶的活性和根的呼吸增强等从而促使CO2排放量增加[18]。此外, 本研究采集的样品是0—10cm, 土壤的营养底物主要集中在表层, 因此表层的气温对土壤呼吸的影响比较显著, 这与Nakajima[9]等人所得出的结果一致。Mi等[20]研究结果表明, 温度对土壤碳矿化的影响极显著, 这可能是不同温度条件下微生物活性不同, 导致了土壤呼吸作用强度的差异。另外, 从呼吸速率的结果来看, 土壤呼吸速率随温度的升高而增加。
3.2 不同森林类型对土壤呼吸作用的影响植被类型对土壤呼吸作用有重要的影响。同一立地条件下, 森林类型对土壤呼吸的影响主要是通过枯枝落叶的数量和分解速率、根系呼吸速率立地条件等影响土壤呼吸作用[21]。本研究结果表明3种不同林地呼吸作用强度由高到低分别为枫香>马尾松>木荷(P < 0.05), 这意味着不同植被类型对土壤碳矿化具有显著影响。有研究表明, 阔叶林地土壤呼吸速率比针叶林要高[22], Raich等对土壤呼吸速率的研究结果表明针叶林土壤呼吸速率一般比相同土壤上的阔叶林的低约10%左右[21], 本研究结果表现为枫香林呼吸速率高于马尾松林, 这与以上研究结果一致[23], 但是木荷林的土壤呼吸速率最低, 低于马尾松林的呼吸速率, 从森林调查结果分析其可能原因是针叶树种凋落物中存在大量顽抗的化合物, 导致了其较低的分解速率并且减慢了颗粒有机物向矿质土壤转化的速度[24], 使得土壤微生物呼吸速率较低。Burton等[25]通过对北美温带地区不同林型细根呼吸的研究, 得出阔叶林根系呼吸明显高于针叶林, 两种林分类型根系呼吸的差异主要来自阔叶树种细根氮含量高于针叶树种。另外, 木荷凋落物所含单宁、蜡质少, 灰分元素较多, 易于分解转化[26], 不利于林地土壤有机质的积累, 具体情况有待于进一步研究。
目前, 室内培养方法广泛应用于土壤潜在碳排放的研究, 由于土壤矿化在没有根系或气候因子等干扰的情况下, 土壤矿化在长时间培养后终将达到平衡。本研究采用单库模式一级动力学方程Cm=Co(1-exp-kt)对土壤矿化潜在碳排放进行模拟, 得出相关系数R2均在0.9以上, 说明拟合效果很好。在5℃条件下, 枫香林地土壤碳矿化量显著高于马尾松和木荷林(P < 0.05), 在其余3种温度条件下, 枫香和马尾松林地土壤潜在碳排放量显著高于木荷(P < 0.05)。
3.3 环境和生物因子对土壤呼吸作用的影响在森林生态系统中, 土壤碳矿化是土壤碳库主要输出途径, 因此, 土壤矿化强度主要受土壤碳库含量的的影响。从土壤全碳与土壤碳矿化的关系来看, 在15℃和25℃条件下, 随土壤全碳含量增加, 土壤碳排放量呈现先增后减的变化趋势(P < 0.05), 在5℃和35℃条件下, CO2累积排放量随土壤全碳含量变化并无显著性差异, 说明土壤全碳对土壤碳矿化的影响同时也受温度调控。土壤矿化除受土壤碳库影响外, 可能还与土壤含水量密切相关, 往往同时取决于温度的配置状况。在一定温度条件下, 土壤含水量适度提高可以增强微生物的活性及微生物的数量, 从而加快土壤碳的矿化速度。本研究结果表明, 在25℃时, 不同林型土壤碳排放量随土壤含水量先增后减的变化趋势, 土壤含水量的极值点为14.93%, 且差异达显著性水平(P < 0.1), 在相同的相对含水量的条件下, 土壤碳排放量随温度的升高而增加, 这一结论与Hartley等研究的结果一致[27], 说明土壤含水量并不是影响土壤碳排放量的调控因子。植被类型和培养温度对土壤碳矿化量有显著影响, 说明植被类型和温度能够对土壤呼吸产生重要影响, 且不同温度对土壤呼吸作用更显著(P < 0.000), 但无交互作用其原因可能是植被类型主要是通过影响根系的密度及分布、土壤生物栖息的环境温度和湿度等来影响土壤碳矿化强度。
3.4 Q10及其主要影响因素土壤呼吸与温度的关系可以从温度敏感性角度来研究, 在没有外界干扰的情况下长时间培养后土壤累积碳排放量将会达到一个峰值, 但土壤呼吸与温度的关系研究比较复杂。本研究从温度敏感性角度来分析土壤碳排放量受温度的影响, 得出不同林型Q10值大小依次为木荷(1.971)>马尾松(1.957)>枫香(1.797), 木荷林地土壤具有较高的Q10值, 表明其土壤呼吸对土壤温度的升高更为敏感, 全球气候变暖可能会强烈地影响其土壤呼吸[28]。不同林型会影响土壤碳矿化及其对温度变化的敏感程度, 主要是对土壤有机碳的数量与结构产生影响, 从而导致Q10值的巨大变异[29]。本研究中4种不同温度变化下, 在15—25℃间的Q10值最大, 在15—25℃这一温度环境下, 土壤微生物活性更强。在5—15℃条件下, 土壤微生物活性没有得到激发导致土壤碳排放量较少, 使得土壤Q10值偏低。在25—35℃较高的温度条件下, 土壤异养呼吸速率受到土壤微生物的影响[30], 表明土壤呼吸敏感性随着升温是降低的, 土壤微生物活性随着土壤温度的升高而降低[31], 也有可能是土壤温度升高改变微生物的种群结构[32], 使得土壤Q10值降低。在不同培养温度条件下不同林型土壤Q10培养范围为1.797—1.971, 变异比较小, 说明不同森林林型在不同温度条件下的Q10值并未表现出显著性差异, 这与王红等研究的结果不一致[33], 其原因可能是林地土壤养分对温度的不同反应所致, Q10值除受温度的影响外, 还可能受其他因素如土壤湿度、根生物量及其他因子的综合结果[34], 而本研究所测试时间为35d, 培养时间较短, 没有全年数据, 对Q10值估算也会产生影响。因此为了更准确地估算不同林型土壤碳矿化对温度的响应得出来的Q10值, 在继续的研究中应尽可能的扩大测定期间, 以减小对Q10值估算的误差[35]。未来气候变化对亚热带森林生态系统碳循环的影响及其机理会更为复杂, 这将是今后的主要研究方向之一。
4 结论不同森林类型土壤CO2累计排放量随温度升高而增加, 且在25℃与35℃的CO2累计排放量显著高于5℃和15℃(P < 0.05)。4种不同温度条件下的林地土壤呼吸速率及土壤CO2累计排放量均表现为:枫香林﹥马尾松林﹥木荷林, 且在25℃与35℃条件下土壤呼吸强度更高。在15℃、25℃、35℃条件下, 3种林地土壤最大潜在碳排放差异均不显著。
在15℃和25℃条件下, 土壤碳排放量随土壤全碳含量呈现出先增后减的变化趋势, 在25℃条件下影响显著(P < 0.05), 全碳的极值点分别为1.83%和1.89%。从不同植被类型和培养温度来看, 它们对土壤碳矿化量均有显著影响, 说明植被类型和温度能够对土壤呼吸产生重要影响, 且不同温度对土壤呼吸作用更显著, 但两因素间无显著交互作用。在25℃时, 不同林型土壤碳排放量随土壤含水量先增后减的变化趋势, 说明土壤含水量并非是影响土壤碳排放量的调控因子。土壤P、MBN和N素对土壤碳排放存在限制作用。
| [1] | Batjes N H. Total carbon and nitrogen in the soils of the world. European Journal of Soil Science, 1996, 47(2): 151–163. DOI:10.1111/ejs.1996.47.issue-2 |
| [2] | Zerva A, Mencuccini M. Short-term effects of clearfelling on soil CO2, CH4, and N2O fluxes in a Sitka spruce plantation. Soil Biology and Biochemistry, 2005, 37(11): 2025–2036. DOI:10.1016/j.soilbio.2005.03.004 |
| [3] | Melillo J M, Steudler P A, Aber J D, Newkirk K, Lux H, Bowles F P, Catricala C, Magill A, Ahrens T, Morrisseau S. Soil warming and carbon-cycle feedbacks to the climate system. Science, 2002, 298(5601): 2173–2176. DOI:10.1126/science.1074153 |
| [4] | Schlesinger W H, Andrews J A. Soil respiration and the global carbon cycle. Biogeochemistry, 2000, 48(1): 7–20. DOI:10.1023/A:1006247623877 |
| [5] | Grace J, Rayment M. Respiration in the balance. Nature, 1999, 404(6780): 819–820. |
| [6] | Cox P M, Betts R A, Jones C D, Spall S A, Totterdell I J. Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model. Nature, 2000, 408(6809): 184–187. DOI:10.1038/35041539 |
| [7] | Li S G, Asanuma J, Eugster W, Kotani A, Liu J J, Urano T, Oikawa T, Davaa G, Oyunbaatar D, Sugita M. Net ecosystem carbon dioxide exchange over grazed steppe in central Mongolia. Global Change Biology, 2005, 11(11): 1941–1955. |
| [8] | Rodeghiero M, Cescatti A. Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps. Global Change Biology, 2005, 11(7): 1024–1041. DOI:10.1111/gcb.2005.11.issue-7 |
| [9] | Nakajima M, Cheng W G, Tang S R, Hori Y, Yaginuma E, Hattori S, Hanayama S, Tawaraya K, Xu X K. Modeling aerobic decomposition of rice straw during the off-rice season in an Andisol paddy soil in a cold temperate region of Japan:effects of soil temperature and moisture. Soil Science and Plant Nutrition, 2016, 62(1): 90–98. DOI:10.1080/00380768.2015.1121116 |
| [10] | Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165–173. DOI:10.1038/nature04514 |
| [11] | Reichstein M, Bednorz F, Broll G, Kätterer T. Temperature dependence of carbon mineralisation:conclusions from a long-term incubation of subalpine soil samples. Soil Biology and Biochemistry, 2000, 32(7): 947–958. DOI:10.1016/S0038-0717(00)00002-X |
| [12] | Reichstein M, Subke J A, Angeli A C, Tenhunen J D. Does the temperature sensitivity of decomposition of soil organic matter depend upon water content, soil horizon, or incubation time?. Global Change Biology, 2005, 11(10): 1754–1767. DOI:10.1111/gcb.2005.11.issue-10 |
| [13] | Fang C, Moncrieff J B. The dependence of soil CO2 efflux on temperature. Soil Biology and Biochemistry, 2001, 33(2): 155–165. DOI:10.1016/S0038-0717(00)00125-5 |
| [14] | 杨玉盛, 董彬, 谢锦升, 陈光水, 高人, 李灵, 王小国, 郭剑芬. 森林土壤呼吸及其对全球变化的响应. 生态学报, 2004, 24(3): 583–591. |
| [15] | 刘苑秋, 杜天真, 郭晓敏, 杨国平, 罗良兴. 退化第四纪红粘土重建森林模式及其土壤恢复研究. 水土保持学报, 2004, 18(6): 34–37. |
| [16] | Chen C R, Xu Z H, Mathers N J. Soil carbon pools in adjacent natural and plantation forests of subtropical Australia. Soil Science Society of America Journal, 2004, 68(1): 282–291. DOI:10.2136/sssaj2004.2820 |
| [17] | Smith J L, Schnabel R R, McNeal B L, Campbell G S. Potential errors in the first-order model for estimating soil nitrogen mineralization potentials. Soil Science Society of America Journal, 1980, 44(5): 996–1000. DOI:10.2136/sssaj1980.03615995004400050025x |
| [18] | 刘颖, 韩士杰. 长白山四种森林土壤呼吸的影响因素. 生态环境学报, 2009, 18(3): 1061–1065. |
| [19] | Jia B, Zhou G, Wang Y, Wang F, Wang X. Effects of temperature and soil water-content on soil respiration of grazed and ungrazed Leymus chinensis steppes, inner Mongolia. Journal of Arid Environments, 2006, 67(1): 60–76. DOI:10.1016/j.jaridenv.2006.02.002 |
| [20] | Mi J, Li J J, Chen D M, Xie Y C, Bai Y F. Predominant control of moisture on soil organic carbon mineralization across a broad range of arid and semiarid ecosystems on the Mongolia plateau. Landscape Ecology, 2015, 30(9): 1683–1699. DOI:10.1007/s10980-014-0040-0 |
| [21] | Raich J W, Tufekciogul A. Vegetation and soil respiration:correlations and controls. Biogeochemistry, 2000, 48(1): 71–90. DOI:10.1023/A:1006112000616 |
| [22] | Hudgens D E, Yavitt J B. Land-use effects on soil methane and carbon dioxide fluxes in forests near Ithaca, New York. Écoscience, 1997, 4(2): 214–222. DOI:10.1080/11956860.1997.11682398 |
| [23] | 易志刚, 蚁伟民, 周国逸, 周丽霞, 张德强, 丁明懋. 鼎湖山三种主要植被类型土壤碳释放研究. 生态学报, 2003, 23(8): 1673–1678. |
| [24] | Leckie S E, Prescott C E, Grayston S J. Forest floor microbial community response to tree species and fertilization of regenerating coniferous forests. Canadian Journal of Forest Research, 2004, 34(7): 1426–1435. DOI:10.1139/x04-028 |
| [25] | Burton A J, Pregitzer K S, Ruess R W, Hendrick R L, Allen M F. Root respiration in North American forests:effects of nitrogen concentration and temperature across biomes. Oecologia, 2002, 131(4): 559–568. DOI:10.1007/s00442-002-0931-7 |
| [26] | 曹汉洋. 杉木、马尾松、木荷纯林及其混交林的土壤养分状况. 南京林业大学学报, 1998, 22(2): 45–48. |
| [27] | Hartley I P, Hopkins D W, Sommerkorn M, Wookey P A. The response of organic matter mineralisation to nutrient and substrate additions in sub-arctic soils. Soil Biology and Biochemistry, 2010, 42(1): 92–100. DOI:10.1016/j.soilbio.2009.10.004 |
| [28] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B:Chemical and Physical Meteorology, 1992, 44(2): 81–99. DOI:10.3402/tellusb.v44i2.15428 |
| [29] | 张金波, 宋长春, 杨文燕. 不同土地利用下土壤呼吸温度敏感性差异及影响因素分析. 环境科学学报, 2005, 25(11): 1537–1542. DOI:10.3321/j.issn:0253-2468.2005.11.017 |
| [30] | 盛浩, 杨玉盛, 陈光水, 高人, 曾宏达, 杜紫贤, 张静. 土壤异养呼吸温度敏感性(Q10)的影响因子. 亚热带资源与环境学报, 2006, 1(1): 74–83. |
| [31] | 王文立, 孔维栋, 曾辉. 土壤微生物对增温响应的Meta分析. 农业环境科学学报, 2015, 34(11): 2169–2175. DOI:10.11654/jaes.2015.11.019 |
| [32] | Zhang W, Parker K M, Luo Y, Wan S, Wallace L L, Hu S. Soil microbial responses to experimental warming and clipping in a tallgrass prairie. Global Change Biology, 2005, 11(2): 266–277. DOI:10.1111/gcb.2005.11.issue-2 |
| [33] | 王红, 范志平, 邓东周, 陈德龙, 孙学凯, 高俊刚, 曾德慧. 不同环境因子对樟子松人工林土壤有机碳矿化的影响. 生态学杂志, 2008, 27(9): 1469–1475. |
| [34] | Wang C K, Yang J Y, Zhang Q Z. Soil respiration in six temperate forests in China. Global Change Biology, 2006, 12(11): 2103–2114. DOI:10.1111/gcb.2006.12.issue-11 |
| [35] | Graf A, Weihermüller L, Huisman J A, Herbst M, Bauer J, Vereecken H. Measurement depth effects on the apparent temperature sensitivity of soil respiration in field studies. Biogeosciences, 2008, 5(4): 1175–1188. DOI:10.5194/bg-5-1175-2008 |