 2018, Vol. 38
2018, Vol. 38文章信息
- 孙雪娇, 常顺利, 张毓涛, 李吉玫.
- SUN Xuejiao, CHANG Shunli, ZHANG Yutao, LI Jimei.
- 天山森林植物功能性状与碳库沿海拔梯度的变化
- The variations in plant functional traits and forest carbon content with altitudinal gradients in the Tianshan Mountains
- 生态学报. 2018, 38(14): 4994-5005
- Acta Ecologica Sinica. 2018, 38(14): 4994-5005
- http://dx.doi.org/10.5846/stxb201707111254
-
文章历史
- 收稿日期: 2017-07-11
- 网络出版日期: 2018-04-04






2. 新疆林科院森林生态研究所, 乌鲁木齐 830063
2. Institute of Forest Ecology, Xinjiang Academy of Forestry, Urumqi 830063, China
植物功能性状是指能够影响植物个体的生存、繁殖和生长, 反映植物对环境变化的响应, 并且能够显著影响生态系统功能的一系列植物性状[1-2]。研究植物功能性状在环境梯度上对生态系统功能的影响, 能更好地认识环境、个体和功能间的相互关系。近年来, 国内外研究者对植物功能性状的研究多是在物种水平, 着重探讨了叶片、细根和整株性状沿环境梯度的变化趋势, 认为环境因子的空间格局影响着植物功能性状的地理分布[3-5]。然而这并不能反映植物通过群落构建来适应环境变化的生态学机制, 也很难建立起植物群落的功能性状与生态系统功能的生理生态学联系。
有研究者提出以群落结构数据为基础计算功能性状的群落加权平均值(community weighted mean, CWM)来代表群落水平的功能性状[6-7]。已有研究表明, 植物功能性状的加权平均值沿着土壤养分和水分梯度[8]、光照梯度[9]、演替阶段[7]以及纬度梯度[10]表现出了明显的变化趋势。海拔差异通常导致温度、降水、光照呈现连续变化[11-12], 使植物功能性状产生复杂的适应性变化。随着海拔升高, 植物个体的叶和根的N、P含量、比根长、最大潜在高度、叶木质素含量等均有明显垂直分布特征[13-20]。但是关于群落尺度上森林植物功能性状沿海拔变化的空间格局研究仍未见报道。
同时, 通过植物功能性状来指示植被碳库的大小成为当前新的研究视角[21-22]。森林碳库在全球碳循环过程中负有重要作用[23-24]。研究显示, 树高、比叶面积、叶片N含量等对植被固碳能力有良好的指示作用[22-25]。然而, 现有的研究多集中于草地和森林植被碳库[24-25], 关于植物功能性状是如何影响森林土壤碳库还不清楚, 并且群落构建如何通过植物功能性状变化来影响碳库更是未见探讨。
天山森林对于整个天山北麓调节气候、固碳释氧、净化大气等生态系统功能有极其重要的作用。叶、根性状以及植株高度是植物对环境适应的长期结果, 其形态和结构的改变会导致植物的功能和行为变化[8], 进而对生态系统功能产生影响[8, 22]。为了探讨森林群落植物功能性状与碳库沿海拔的变化规律及二者之间关系, 为全球变化背景下的森林管理及功能预测提供依据。本文通过在沿海拔设置的系列样地上, 测定群落尺度上植株高度、叶、细根功能性状指标, 估算森林生态系统碳密度, 以期探讨下列问题:(1)群落功能性状和碳密度沿海拔梯度存在何种分异规律;(2)哪些植物功能性状可作为反映森林群落碳密度的指标;(3)影响植物功能性状在海拔梯度上连续变化的环境因子有哪些。
1 材料与方法 1.1 研究区概况本研究依托中国森林生态系统研究网络天山森林生态系统定位研究站(87°07′—87°28′E, 43°14′—43°26′N)开展工作, 该地位于新疆乌鲁木齐县板房沟林场, 属大陆温带气候, 最大积雪深度为65 cm, 年总辐射约为5.85×105 J cm-2 a-1。天山北坡受到中纬度西风带的控制, 来自大西洋和和北冰洋的湿润气流从西方或西北方吹入, 受天山阻隔, 水热条件随地形和海拔重新分配, 垂直变化明显[26-27](表 1)。本研究样地布设在海拔1700—2700 m, 土壤类型为灰褐色森林土, 雪岭云杉(Picea schrenkiana Fisch. et Mey.)是天山森林的单优乔木, 林下和林缘分布的灌木有小檗(Berberis heteropoda Schrenk及B. nummularia Bge.)、蔷薇(Rosa spinosissima L.)、栒子(Cotoneaster melanocarpus Lodd.)绣线菊(Spiraea hypericifolia L.)、方枝柏(Juniperus pseudosabina Fisch. et Mey.)、忍冬(Lonicera hispida Pall. ex Roem. et Schunt), 林下草本主要由老鹳草(Geranium rotundifolium L.)、天山羽衣草(Alchemilla tianschanica Juz.)、羊角芹(Aegopodium podagraria L.)等构成[28]。
| 海拔 Elevation/m |
年均温 Annual average temperature/℃ |
年降水量 Annual average precipitation/mm |
Kira干湿度指数 Kira humidity index |
| 1500 | 4.76 | 388 | 11.42 |
| 2000 | 2.282 | 543 | 18.06 |
| 2200 | 1.7 | 528 | 18.32 |
| 2600 | -0.54 | 495 | 18.79 |
| 2700 | -1.1 | 489 | 19.01 |
| Kira干湿度指数=P/(ET+20), P为年降水量, ET为月平均气温高于5℃的总和 | |||
2009年, 在研究区沿海拔1700—2700 m布设两条样带, 在每条样带内以200 m为一个海拔梯度, 在每个海拔梯度内以每50 m海拔为一个小区, 在每个小区内设置了1—2个20 m×20 m的固定样方, 共计43个固定样地。同时调查了每个样方的基本群落信息和土壤因子(包括各土层(0—20, 20—40, 40—60, 60—80, 80—100 cm)土壤有机碳含量、土壤密度、土壤有机质、速效氮、速效磷、速效钾含量和直径大于2 mm石砾所占体积)。
2013年, 对雪岭云杉的生物量进行调查:根据树龄、胸径和冠幅的信息选取30株标准木, 对30株标准木进行全株采伐解析, 将每株标准木的冠层分为上、中、下3层, 测量每层每个枝条的长度、基径, 并分别算出该层枝条长度、基径的平均值, 然后以每层枝条的平均值为依据在上、中、下各层分别选取3个标准枝, 摘取叶片, 分别对叶片和枝条称量鲜重;对树干采用分层切割收获法, 每隔1.5 m截取树木圆盘并称量鲜重;将每株标准木的根系全部挖出, 称量鲜重;然后对每株标准株的叶片、枝条和根系进行随机取样, 称量鲜重后装入样品袋内标号并将所有树木圆盘带回实验室烘干后称量干重。
2016年7—9月, 对43个固定样地进行复查, 调查样地内所有木本植物的胸径/地径、株高、冠幅等, 并在每个固定样地内沿对角线设置3个1 m×1 m的草本样方, 调查草本样方内所有物种的平均高度、个数、盖度等信息。
1.2.2 样品采集分别对43个固定样地内的木本层(包括乔木和灌木)和草本层进行采样。
对于木本层, 在每个采样点, 选定3棵采样株, 采集东、西、南、北4个方位的中部的新生叶, 将不同部位的叶片混合, 采用四分法取样, 同时采集小于2 mm细根装入样品袋标记, 带回实验室处理。
对于草本层, 采集草本样方内全部的地上及地下部分, 装入样品袋标记, 带回实验室处理;
对于出现在样地内的所有物种, 采集2—3枚当年生新鲜叶片, 放入自封袋内带回实验室用于叶绿素含量的测定。
1.3 样品测试分析方法植物功能性状指标的选取、测定及其生态学意义如表 2所示。
| 植物功能性状指标 Functional traits |
英文名称 English name |
缩写 Abbreviation |
测定方法 Measured method |
生态学意义 Ecological meaning |
| 叶片碳、氮、磷含量, 细根碳、氮、磷含量 | Leaf and fine root carbon、nitrogen、phosphorus content | Cleaf, Nleaf, Pleaf, Croot, Nroot, Proot | 重铬酸钾外加热法、硫酸-高氯酸消煮 | 参与和影响着植物的代谢过程和生长发育[18] |
| 叶片碳氮比、氮磷比 | Leaf C/N and N/P ratio | C/N, N/P | 叶片C含量/叶片N含量叶片N含量/叶片P含量 | 反映植物对养分的利用效率和限制状况[18] |
| 叶片干物质含量 | Leaf dry matter content | LDMC | 叶干重/叶鲜重 | 代表植物叶片获取资源的能力, 影响其机械强度和可燃性[29] |
| 叶木质素含量 | Leaf lignin content | LLC | 重铬酸钾氧化法 | 影响凋落叶片分解过程[5] |
| 叶绿素 | Chlorophyll | Chl | 分光光度法 | 参与植物的光合作用[13] |
| 细根干物质含量 | Fine root dry matter content | RDMC | 细根干重/细根鲜重 | 植物细根获取资源的能力, 植物对细根建成的投入[13] |
| 比根长 | Specific root length | SRL | 细根长/细根干重 | 影响植物吸收对水养等的能力[13] |
| 植株高度 | Height | H | 直尺、卷尺和树高仪 | 影响植物光竞争能力, 抗弯强度, 抗压强度等[1] |
首先计算30株雪岭云杉标准木的生物量, 以胸径的平方与树高的乘积(D2H)为自变量, 建立雪岭云杉的生物量模型。灌木的生物量模型参考在相同研究区的研究成果(表 3)。
| 树种 Species |
生物量模型 Biomass model |
R2 | 参考文献 References |
| 雪岭云杉P. Schrenkiana | W=0.7489(D2H)3.4764 | 0.9799 | — |
| 多刺蔷薇R. acicularis | W=-2.535+2.325V-0.007V3 | 0.998 | 仇瑶等[28] |
| 忍冬L. hispida | W=7.448-9(D2H)2.753 | 0.89 | 仇瑶等[28] |
| 小檗B. thunbergii | W=-3061.6+1251.5(D2H) | 0.962 | 王文栋等[30] |
| 绣线菊S. salicifolia | W=1990(D2H)0.97 | 0.892 | 王文栋等[30] |
| 黑果栒子C. melanocarpus | W=5.654-0.11V2+0.001V3 | 0.992 | 仇瑶等[28] |
| 新疆方枝柏J. pseudosabina | W=0.881+0.021(D2H) | 0.85 | 仇瑶等[28] |
根据生物量模型及对43个样地的草本调查, 计算每一样地内各层生物量, 以式(1)计算植被碳密度:

|
(1) |
式中, VCD为植被碳密度(Mg/hm2), BECi为各植被层(乔木层、灌木层、草本层)生物量(Mg), VCi为各植被层含碳率, 其中乔木层含碳率为49%[31], 灌木层含碳率为50%[32], 草本层含碳率为42%[31]。
土壤碳密度计算如式(2)所示:
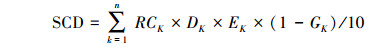
|
(2) |
式中, SCD为土壤有机碳密度(Mg/hm2), K为土壤层次, SCK为第K层土壤有机碳含量(g/kg), DK为第K层土壤密度(g/cm3), EK为第K层土层厚度(cm), GK为第K层土层中直径大于2 mm石砾所占体积百分比(%)。
群落总碳密度(TCD)=植被碳密度(VCD)+土壤碳密度(SCD)
1.4.2 群落植物功能性状的计算
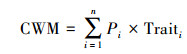
|
(3) |
式中, CWM为群落尺度上的植物功能性状, Pi为相对生物量, Traiti为各层植物功能性状。
草本层面Chl的计算, 首先以草本调查信息计算各草本物种的重要值(重要值=相对高度+相对多度+相对盖度)以重要值加权计算草本层面Chl含量, 再以(3)式计算群落尺度上Chl含量。
下文中所提及本研究测定的性状指标均为群落尺度上的植物功能性状。
1.4.3 数据分析方法在Excel 2013(Microsoft Corporation)中对各指标进行均值化、去除离异值处理, 在SPSS 17.0(IBM)中, 对指标进行单样本K-S检验, 所有指标均服从正态分布(P > 0.05), 以单因素方差分析(one-way anova)中最小显著差数(LSD)法进行多重比较;环境因子与植物功能性状的之间的关系采用相关性分析。
用Origin pro 9.0(Originlab Corporation)绘制各指标在不同海拔梯度上的箱线图。
以Canoco5.0 (Biometris, Plant Research International)中的DCA分析和RDA分析探讨碳密度与植物功能性状的关系;用R-3.2.0 (The R Foundation for Statistical Computing)对各组分碳密度与植物功能性状指标进行相关性分析, 选择P < 0.05的指标以leaps包对各组分碳密度与植物功能性状进行全子集回归, 筛选调整后的R2最高的5—15个指标组合模式以lm函数对其进行多元线性回归, 按照下述标准选择最优模型:(1)模型中各变量的显著水平较高(2)模型整体的AIC值最小(3)在满足前两个原则下, 模型中的变量个数越少越好。
2 结果分析 2.1 群落植物功能性状和碳密度的垂直分异规律 2.1.1 群落叶功能性状的垂直分异规律群落尺度上叶片性状沿海拔的变化规律见图 1。Cleaf、Pleaf均随海拔升高至2100—2300 m梯度内达到最大值而后有所下降, 其在2100—2300 m和2300—2500 m海拔内显著高于其余梯度(P < 0.05)。LLC随海拔表现为先升后降, 但各海拔段间无显著性差异(P > 0.05)。Nleaf、Chl、N/P随海拔升高逐渐降低, Nleaf在各海拔段间无显著性差异(P > 0.05)。LDMC在1700—2700 m海拔内处于0.33—0.64 g/g之间, 先升高后趋于平缓, 各海拔段间无显著差异(P > 0.05)。C/N随海拔逐渐上升, 并且在高海拔区域2100—2700 m显著高于1700—1900 m(P < 0.05)。

|
| 图 1 群落叶功能性状随海拔的变化规律 Fig. 1 Community functional traits of leaf change with altitude Cleaf:叶片碳含量, Leaf carbon content;Nleaf:叶片氮含量, Leaf nitrogen content;Pleaf:叶片磷含量, Leaf phosphorus content; C/N:叶碳氮比, Leaf C/N ratio;N/P:叶片氮磷比, Leaf N/P ratio;LDMC:叶片干物质含量, Leaf dry matter content; LLC:叶片木质素含量, Leaf lignin content; Chl:叶绿素Chlorophyll;图中不同小写字母a、b、c表示P < 0.05水平下差异显著, 相同表示不显著, NF表示各海拔梯度间无显著差异 |
群落尺度上细根性状和植株高度沿海拔的变化规律见图 2。结果表明, 在海拔1700—2700 m, Croot处于37.14%—83.25%的范围内, 并随着海拔的升高逐渐升高, 其中在2100—2300 m梯度显著高于1700—1900 m处(P < 0.05);Nroot在1.25%—1.61%范围内, 随海拔先降低后升高;海拔的升高导致Proot逐渐降低, 但除在2300—2500 m时显著(P < 0.05)低于1700—1900 m外, 其余各海拔段间未有显著差异(P > 0.05);RDMC、SRL和H随海拔均表现先升高后降低, 但RDMC在各海拔段间无显著差异(P > 0.05)。

|
| 图 2 群落细根性状和植株高度随海拔的变化规律 Fig. 2 Community functional traits of fine root change with altitude Croot:细根碳含量, Fine root carbon content;Nroot:细根氮含量, Fine root nitrogen content;Proot:细根磷含量, Fine root phosphorus content;SRL:比根长, Specific root length; RDMC:细根干物质含量, Fine root dry matter content;H:株高, Height |
各组分碳密度沿海拔的变化规律见图 3。结果表明, 在1900—2300 m, 随着海拔升高, VCD显著大幅增加, 最小仅为19.57 Mg/hm2, 最大为61.89 Mg/hm2, 随后在2300—2700 m范围又有所降低。SCD随着海拔逐渐递增, 各样地最大SCD为555 Mg/hm2;各样地VCD均远小于SCD, SCD是生态系统碳库的主要贮存形式。随着海拔升高群落TCD先大幅增加, 随后涨势趋于平缓, 并在1700—1900 m处显著(P < 0.05)低于其他海拔梯度。

|
| 图 3 各组分碳密度随海拔的分布规律 Fig. 3 Carbon density change with altitude VCD:植被碳密度, vegetation carbon density; SCD:土壤碳密度, soil carbon density;TCD:总碳密度, Total carbon density |
以冗余分析(RDA)方法探讨各组分碳密度与群落植物功能性状之间的关系。研究表明, 四个排序轴对海拔梯度上各组分碳密度的累积解释解释量达93.8%, 可解释100%的碳密度-植物功能性状之间的相关性。RDA排序的第一、二轴的特征值分别为0.743和0.047, 解释了各组分碳密度变化的77.5%。根据前两个排序轴做出二维排序图(图 4)。第一轴随着H、SRL、Cleaf的增加, VCD、SCD和TCD均逐渐增大。第二轴与Pleaf、SDMC、Proot和Chl有较强的负相关性, 与叶片N/P有较强的负相关关系;VCD与H、Cleaf、Pleaf呈正相关。SCD和TCD与H、SRL、Croot、Cleaf成正相关关系, 而与Chl、LLC和Proot呈负相关。对各植物功能性状指标对排序的贡献进行检验, H对排序解释量最大, 为64%(P < 0.01)。Cleaf、SRL、Croot、RDMC对排序的贡献率其次, 分别为17%、10%、5%和4%(P < 0.05)。

|
| 图 4 碳密度与植物功能性状的二维排序图 Fig. 4 Two dimensional RDA ordination diagram of carbon density with plant functional traits 箭头及排序轴之间的夹角表示各指标及排序轴之间相关性的正负, 小于90°为正相关关系, 等于90°, 二者之间无相关性, 大于90°, 为负相关。箭头线段在排序轴和另一箭头线段及其延长线上的投影表示二者之间相关性的大小, 投影越大, 相关性越强 |
各组分碳密度与群落植物功能性状做相关性分析见表 4, VCD与H、Cleaf、Pleaf、SRL和C/N极显著(P < 0.01)正相关, 与RDMC和N/P(P < 0.05)显著负相关, SCD和TCD与H、Croot、SRL显著(P < 0.05)正相关而与LLC、Chl、Proot显著(P < 0.05)负相关。
| 碳密度 Carbon density |
H | Cleaf | Nleaf | Pleaf | LLC | LDMC | Chl | Proot | Nroot | Croot | SRL | RDMC | C/N | N/P |
| VCD | 0.703** | 0.591** | 0.158 | 0.400** | 0.081 | 0.188 | -0.102 | -0.152 | -0.132 | 0.168 | 0.425** | -0.330* | 0.436** | -0.306* |
| SCD | 0.505** | 0.186 | 0.004 | -0.014 | -0.445** | -0.066 | -0.370* | -0.359* | -0.044 | 0.488** | 0.316* | -0.057 | -0.027 | 0.059 |
| TCD | 0.646** | 0.312* | -0.037 | 0.089 | -0.405* | -0.013 | -0.368* | -0.363* | -0.076 | 0.497** | 0.386* | -0.126 | 0.085 | -0.024 |
| ***代表P < 0.001显著水平, **代表P < 0.01显著水平, *代表P < 0.05显著水平, 下同。VCD:植被碳密度, vegetation carbon density; SCD:土壤碳密度, soil carbon density;TCD:总碳密度, Total carbon density | ||||||||||||||
应用多元线性回归模型对各组分碳密度与植物功能性状指标间的关系进行建模, 结果见表 5。VCD模型中纳入H、Cleaf两个指标以及常数项, 其中Cleaf及常数项未达到显著水平, 但其P值处于0.05—0.1之间, 植株高度达到P < 0.001的显著水平, 总体上, 模型调整后的R2为0.51(P < 0.001)。土壤碳密度模型中包含H、LLC, Chl、Proot及常数项(P < 0.05), 模型调整后的R2为0.43(P < 0.001)。总碳密度模型纳入指标H、SRL、Chl及Proot, 除SRL的P值为0.52外, 其余指标和常数项均达P < 0.05的显著水平, 模型调整后R2为0.55(P < 0.001)。
| 碳密度/(Mg/hm2) Carbon density |
性状 Trait |
估计值 Estimated value |
误差平方和 Std. Error |
t | P |
| VCD | H/m | 4.17 | 0.88 | 4.73 | 2.83×10-5*** |
| Cleaf/% | 0.85 | 0.45 | 1.92 | 0.06 | |
| 常数项 | -35.85 | 20.00 | -1.79 | 0.08 | |
| 模型 | VCD=4.17H+0.85Cleaf-35.85 F=23.21 adjr2=0.51 P=2.032×10-7 | ||||
| SCD | H/m | 8.77 | 3.18 | 2.75 | 0.009** |
| LLC/(g/kg) | -0.77 | 0.34 | -2.28 | 0.028* | |
| Chl/(mg/g) | -99.40 | 41.15 | -2.42 | 0.021* | |
| Proot/% | -545.26 | 202.43 | -2.69 | 0.01* | |
| 常数项 | 639.99 | 104.66 | 6.12 | 3.96×10-7*** | |
| 模型 | SCD=8.77H-0.77LLC-99.4Chl-545.26Proot-639.99 F=8.93 adjr2=0.43 P=3.49×10-5 | ||||
| TCD | H/m | 12.50 | 3.26 | 3.83 | 0.00047*** |
| SRL/(m/g) | 239.34 | 119.11 | 2.01 | 0.052 | |
| Chl/(mg/g) | -112.95 | 39.81 | -2.84 | 0.0073** | |
| Proot/% | -564.12 | 194.42 | -2.90 | 0.0061** | |
| 常数项 | 303.62 | 130.02 | 2.34 | 0.025* | |
| 模型 | TCD=12.5H+239.34SRL-112.95Chl-564.12Proot+303.62 F=13.91 adjr2=0.55 P=4.454×10-7 | ||||
植物功能性状的环境因子的相关性分析见表 6。H和Croot与土壤有机质和Kira干湿度指数均显著(P < 0.05)正相关, Cleaf与有机质显著(P < 0.05)正相关, Pleaf和SRL与土壤速效磷显著(P < 0.05)正相关, LLC与年降水量显著(P < 0.05)正相关, LDMC和C/N与Kira干湿度指数显著(P < 0.05)正相关, Chl与有机质极显著(P < 0.05)负相关而与年均温显著(P < 0.05)正相关, Proot与年均温显著(P < 0.05)正相关, Nroot和RDMC与土壤速效氮和年降水量相关性显著(P < 0.05)。N/P与土壤速效磷和Kira干湿度指数均显著(P < 0.05)负相关。
| 性状 Trait |
土壤有机质 Soil organic matter/ (g/kg) (n=43) |
土壤速效氮 Soil available nitrogen/ (mg/kg) (n=43) |
土壤速效磷 Soil available phosphorus/ (mg/kg) (n=43) |
年均温 Annual average temperature/℃ (n=5) |
年降水量 Annual average precipitation/mm (n=5) |
Kira干湿度指数 Kira humidity index (n=5) |
| H | 0.319* | 0.223 | 0.298 | -0.787 | 0.481 | 0.949* |
| Cleaf | 0.374* | 0.109 | 0.306 | -0.375 | 0.593 | 0.692 |
| Nleaf | -0.108 | -0.099 | -0.073 | 0.796 | -0.265 | -0.796 |
| Pleaf | 0.208 | 0.269 | 0.480** | -0.140 | 0.348 | 0.375 |
| LLC | -0.216 | -0.004 | -0.037 | 0.083 | 0.957* | 0.484 |
| LDMC | 0.024 | 0.011 | 0.213 | -0.845 | 0.502 | 0.984** |
| Chl | -0.529** | -0.099 | 0.239 | 0.929* | 0.357 | -0.568 |
| Proot | -0.061 | -0.141 | 0.029 | 0.932* | -0.132 | -0.851 |
| Nroot | 0.242 | 0.333* | -0.313 | -0.007 | -0.960** | -0.541 |
| Croot | 0.325* | 0.020 | 0.053 | -0.805 | 0.395 | 0.922* |
| SRL | -0.058 | -0.038 | 0.352* | -0.076 | 0.720 | 0.517 |
| RDMC | -0.293 | 0.321* | -0.158 | 0.298 | 0.902* | 0.259 |
| C/N | 0.203 | 0.202 | 0.098 | -0.860 | 0.305 | 0.901* |
| N/P | -0.206 | 0.219 | -0.377* | 0.808 | -0.503 | -0.974** |
群落功能性状的变化除了受到遗产基因调控外, 群落物种组成变化、植物功能性状间的影响、气候土壤等环境因子也是导致其适应性变化的主要因素。
植株高度会受到温度和降水的双重影响, 因此与kira干湿度指数显著正相关, 有研究证明植株高度可指示森林带温度降水特征[27]。除此之外, 植株高度还受到土壤有机质的影响, 这是因为土壤有机质为植物生长提供碳源, 对植物的生存发育至关重要。
土壤环境中可利用营养元素的百分比会直接影响植物器官内C、N、P元素的含量[18], 但植物体内元素在各器官中的运输、分配存在异速增长[33]。Cleaf、Croot与土壤有机质含量均显著正相关。同时温度降水条件会影响土壤中C元素的分解转化过程[34], 使得Croot和C/N也受到干湿度指数的影响。C是构成LLC的主要元素, 因此LLC与Cleaf随海拔的变化趋势表现一致, 整体表现为先升高后降低。另外, 与刘林等的研究中相同[35], LLC还受到降水量的显著影响。
随海拔升高, 温度降低, 会减缓土壤中磷酸钙的溶解[36], 导致Proot随海拔逐渐降低。P作为核酸、磷脂及部分酶的组分, 在植物的生理代谢中起关键作用[18], 随海拔升高辐射增强, 植物将更多的P用于构建光合器官以增强光合作用, 但在高海拔地区, P含量的供给不足导致Pleaf逐渐下降。谢锦等的研究显示, 天山森林主要受到磷元素的限制[18], 因此叶片N/P主要受到土壤速效磷含量的影响。
叶绿素是植物最为重要的光合色素[37], 主要由N组成, 因此Chl与Nleaf呈正比, 在海拔上的变化均表现为逐渐降低, 同时随海拔升高, 光合辐射、紫外线均升高, 较低的Chl可减少植物对光能的吸收, 这也是植物在高光强胁迫下的一种生活对策。与樊江文等对草地植物根系的研究结果相同[38], 本文Nroot除受土壤速效氮的影响外, 与降水量呈显著负相关, 在海拔梯度上表现为先降低后增加, 这可能是由于降水的增加降低了土壤中可利用N元素的浓度所致。
LDMC在各海拔梯度间无显著变化, 可能是LDMC主要受到物种间基因的调控, 而天山森林群落结构较为单一。另一方面, 随海拔升高, 年降水量先增后减, 在1700—1900 m海拔范围内, 群落主要以小檗等耐旱灌木优势种为主, LDMC和RDMC较高海拔以雪岭云杉为主的针叶森林来说稍低, 因此LDMC与Kira干湿度指数呈极显著正相关。细根作为吸收水分和养分的器官, 细根的形态指标SRL和RDMC与植物的生理生长状态以及土壤中水分、养分含量密切相关[39], 而降水会影响土壤中营养元素的浓度和可利用性, 因此在天山森林, SRL和RDMC主要受到土壤速效氮、磷以及降水量的影响。
3.1.2 植物功能性状与生态系统碳密度之间的联系机制生态系统碳库的累积在于碳固定和碳释放的平衡, 植被的光合作用是碳进入有机体环境固定最为重要的一环[24]。SRL反映了植物吸收营养物质的能力[13], 而C、N、P等化学计量特征会通过养分限制对光合作用进行调控[33], C/N和N/P则表征植物对养分的利用效率, 这些植物功能性状指标通过调控光合作用的大小而与VCD显著相关。
土壤中的碳来自植物分泌物及残体的归还, 其储量主要受到植被光合利用率、物种分布、气候条件、土壤环境等因素的影响[40]。因此H和SRL等同样会影响到土壤碳密度的大小。而Cleaf、Pleaf等影响VCD的因素则反映在了与土壤环境更为密切的Croot和Proot上。SCD的高低还与植物残体的分解有关[24], 高LLC降低了植物残体的分解速率, 导致土壤中碳的累积变慢。
与同类研究结论相近[24], 本文测算得到的天山森林SCD为VCD的4倍左右, 这表明生态系统的碳主要以土壤碳库的形式贮存。在低海拔处, H、SRL、Cleaf、Pleaf等性状均较低, 光合作用速率相对较慢, 导致植被和土壤碳库较小, 而在水热条件最为适中的2100 m海拔上下, 腐殖质得到累积, 随着H、Cleaf、SRL的升高土壤碳库和植被碳库均有所增加。在高海拔处由于低温的环境, 土壤微生物活性较低, 生态系统C循环周转较慢, 因而植被具有低H、低Cleaf、低Pleaf的特点, 导致土壤碳库增加而植被碳库降低。
本研究由于实验条件的限制, 在植物功能性状指标上的选取还不完备, 各地区因气候条件、地形、土壤类型的不同而具有不同的植被类型, 因此关于碳库与植物功能性状间的关系还需在更大尺度上及生态系统中进行深入研究。
3.2 结论综上, 群落的物种组成结构、气候、土壤等环境因子、性状间的相互影响等导致了天山森林功能性状在群落尺度上具有明显的垂直分布特征;群落植株高度、比根长以及叶C、P含量通过影响资源的获取、营养元素的限制及利用, 进而影响碳固定过程;叶片木质素含量则通过参与植物残体的分解而影响碳释放过程, 对森林碳库具有良好的指示作用。
| [1] | 孟婷婷, 倪健, 王国宏. 植物功能性状与环境和生态系统功能. 植物生态学报, 2007, 31(1): 150–165. |
| [2] | Kumar J, Pratap A, Kumar S. Phenomics in Crop Plants: Trends, Options and Limitations. India: Springer, 2015. |
| [3] | Li R H, Zhu S D, Chen H Y H, John R, Zhou G Y, Zhang D Q, Zhang Q M, Ye Q. Are functional traits a good predictor of global change impacts on tree species abundance dynamics in a subtropical forest?. Ecology Letters, 2015, 18(11): 1181–1189. DOI:10.1111/ele.12497 |
| [4] | Lebrija-Trejos E, Pérez-García E A, Meave J A, Bongers F, Poorter L. Functional traits and environmental filtering drive community assembly in a species-rich tropical system. Ecology, 2010, 91(2): 386–398. DOI:10.1890/08-1449.1 |
| [5] | 李丹, 康萨如拉, 赵梦颖, 张庆, 任海娟, 任婧, 周俊梅, 王珍, 吴仁吉, 牛建明. 内蒙古羊草草原不同退化阶段土壤养分与植物功能性状的关系. 植物生态学报, 2016, 40(10): 991–1002. DOI:10.17521/cjpe.2015.0465 |
| [6] | 龚时慧, 温仲明, 施宇. 延河流域植物群落功能性状对环境梯度的响应. 生态学报, 2011, 31(20): 6088–6097. |
| [7] | 卜文圣, 臧润国, 丁易, 张俊艳, 阮云泽. 海南岛热带低地雨林植物群落水平功能性状与环境因子相关性随演替阶段的变化. 生物多样性, 2013, 21(3): 278–287. |
| [8] | Vile D, Shipley B, Garnier E. Ecosystem productivity can be predicted from potential relative growth rate and species abundance. Ecology Letters, 2006, 9(9): 1061–1067. DOI:10.1111/j.1461-0248.2006.00958.x |
| [9] | Pérez-Ramos I M, Roumet C, Cruz P, Blanchard A, Autran P, Garnier E. Evidence for a 'plant community economics spectrum' driven by nutrient and water limitations in a Mediterranean rangeland of southern France. Journal of Ecology, 2012, 100(6): 1315–1327. DOI:10.1111/jec.2012.100.issue-6 |
| [10] | 王瑞丽, 于贵瑞, 何念鹏, 王秋凤, 赵宁, 徐志伟. 中国森林叶片功能属性的纬度格局及其影响因素. 地理学报, 2015, 70(11): 1735–1746. |
| [11] | 李艳忠, 罗格平, 许文强, 尹昌应, 韩其飞, 冯异星. 天山北坡三工河流域中山带森林发育与气候土壤的关系. 山地学报, 2011, 29(1): 33–42. |
| [12] | 潘学鹏, 吴喜芳, 沈彦俊, 刘峰贵, 张存桂. 三江并流河源区植被覆盖度对气候要素的响应. 山地学报, 2015, 33(2): 218–226. |
| [13] | 辛福梅, 刘济铭, 杨小林, 赵垦田. 色季拉山急尖长苞冷杉叶片及细根性状随海拔的变异特征. 生态学报, 2017, 37(8): 2719–2728. |
| [14] | 胡启鹏, 郭志华, 孙玲玲, 王彬. 长白山林线树种岳桦幼树叶功能型性状随海拔梯度的变化. 生态学报, 2013, 33(12): 3594–3601. |
| [15] | 马闯, 张文辉, 吴敏, 马莉薇, 周建云, 薛瑶芹. 栓皮栎幼苗根系发育对环境异质性的响应. 林业科学, 2013, 49(10): 58–65. |
| [16] | 刘玉平, 刘贵峰, 达福白乙拉, 程伟燕, 陈志婧, 姜丽丽. 地形因子对大青沟自然保护区不同森林群落叶性状的影响. 林业科学, 2017, 53(3): 154–162. DOI:10.11707/j.1001-7488.20170317 |
| [17] | Loiola P P, Scherer-Lorenzen M, Batalha M A. The role of environmental filters and functional traits in predicting the root biomass and productivity in savannas and tropical seasonal forests. Forest Ecology and Management, 2015, 342: 49–55. DOI:10.1016/j.foreco.2015.01.014 |
| [18] | 谢锦, 常顺利, 张毓涛, 王慧杰, 宋成程, 何平, 孙雪娇. 天山北坡植物土壤生态化学计量特征的垂直地带性. 生态学报, 2016, 36(14): 4363–4372. |
| [19] | 张慧文, 马剑英, 孙伟, 陈发虎. 不同海拔天山云杉叶功能性状及其与土壤因子的关系. 生态学报, 2010, 30(21): 5747–5758. |
| [20] | 杨锐, 张博睿, 王玲玲, 杨建军, 苏文华, 张光飞, 杨波, 周睿. 元谋干热河谷植物功能性状组合的海拔梯度响应. 生态环境学报, 2015, 24(1): 49–56. |
| [21] | 徐琨, 李芳兰, 苟水燕, 包维楷. 岷江干旱河谷25种植物一年生植株根系功能性状及相互关系. 生态学报, 2012, 32(1): 215–225. |
| [22] | 黄海侠. 植物功能性状对森林生态系统服务功能的指示[D]. 上海: 华东师范大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10269-1014325719.htm |
| [23] | Woodwell G M, Whittaker R H, Reiners W A, Likens G E, Delwiche C C, Botkin D B. The biota and the world carbon budget. Science, 1978, 199(4325): 141–146. DOI:10.1126/science.199.4325.141 |
| [24] | 王兴昌, 王传宽. 森林生态系统碳循环的基本概念和野外测定方法评述. 生态学报, 2015, 35(13): 4241–4256. |
| [25] | Manning P, de Vries F T, Tallowin J R B, Smith R, Mortimer S R, Pilgrim E S, Harrison K A, Wright D G, Quirk H, Benson J, Shipley B, Cornelissen J H C, Kattge J, Bönisch G, Wirth C, Bardgett R D. Simple measures of climate, soil properties and plant traits predict national-scale grassland soil carbon stocks. Journal of Applied Ecology, 2015, 52(5): 1188–1196. DOI:10.1111/1365-2664.12478 |
| [26] | 陈曦, 许文强, 罗格平, 蔺卿, 肖鲁湘. 天山北坡不同环境条件下雪岭云杉(Picea schrenkiana)林限土壤属性. 生态学报, 2008, 28(1): 53–61. |
| [27] | 阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平. 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素. 生态学报, 2014, 34(7): 1626–1634. |
| [28] | 仇瑶, 常顺利, 张毓涛, 王文栋, 何平, 王慧杰, 谢锦. 天山林区六种灌木生物量的建模及其器官分配的适应性. 生态学报, 2015, 35(23): 7842–7851. |
| [29] | Moretti M, Dias A T C, de Bello F, Altermatt F, Chown S L, Azcárate F M, Bell J R, Fournier B, Hedde M, Hortal J, Ibanez S, Öckinger E, Sousa J P, Ellers J, Berg M P. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Functional Ecology, 2016, 31(3): 558–567. |
| [30] | 王文栋, 白志强, 阿里木·买买提, 刘端, 郭忠军. 天山林区6种优势种灌木林生物量比较及估测模型. 生态学报, 2016, 36(9): 2695–2704. |
| [31] | 许文强, 杨辽, 陈曦, 高亚琪, 王蕾. 天山森林生态系统碳储量格局及其影响因素. 植物生态学报, 2016, 40(4): 364–373. DOI:10.17521/cjpe.2015.0235 |
| [32] | 方精云, 刘国华, 朱彪, 王效科, 刘绍辉. 北京东灵山三种温带森林生态系统的碳循环. 中国科学D辑地球科学, 2006, 36(6): 533–543. |
| [33] | 陈婵, 王光军, 赵月, 周国新, 李栎, 高吉权. 会同杉木器官间C、N、P化学计量比的季节动态与异速生长关系. 生态学报, 2016, 36(23): 7614–7623. |
| [34] | 周晓宇, 张称意, 郭广芬. 气候变化对森林土壤有机碳贮藏影响的研究进展. 应用生态学报, 2010, 21(7): 1867–1874. |
| [35] | 刘林. 模拟增加降水对干旱河谷区云南松人工林凋落叶分解及养分释放的影响[D]. 成都: 四川农业大学, 2015. http://cdmd.cnki.com.cn/Article/CDMD-10626-1016048529.htm |
| [36] | 张巧明, 王得祥, 龚明贵, 张丽楠. 秦岭火地塘林区不同海拔森林土壤理化性质. 水土保持学报, 2011, 25(5): 69–73. |
| [37] | 王海珍, 韩路, 徐雅丽, 牛建龙, 于军. 土壤水分梯度对灰胡杨光合作用与抗逆性的影响. 生态学报, 2017, 37(2): 432–442. |
| [38] | 樊江文, 张良侠, 张文彦, 钟华平. 中国草地样带植物根系N、P元素特征及其与地理气候因子的关系. 草业学报, 2014, 23(5): 69–76. DOI:10.11686/cyxb20140508 |
| [39] | 韦兰英, 上官周平. 黄土高原白羊草、沙棘和辽东栎细根比根长特性. 生态学报, 2006, 26(12): 4164–4170. DOI:10.3321/j.issn:1000-0933.2006.12.032 |
| [40] | 张全军, 于秀波, 钱建鑫, 熊挺. 鄱阳湖南矶湿地优势植物群落及土壤有机质和营养元素分布特征. 生态学报, 2012, 32(12): 3656–3669. |