 2018, Vol. 38
2018, Vol. 38文章信息
- 刘潇潇, 何秋月, 闫美杰, 李国庆, 王仕稳, 杜盛.
- LIU Xiaoxiao, HE Qiuyue, YAN Meijie, LI Guoqing, WANG Shiwen, DU Sheng.
- 黄土丘陵区辽东栎群落优势种和主要伴生种树干液流动态特征
- Characteristics of sap flow dynamics in dominant and companion trees in a natural secondary oak forest in the loess hilly region
- 生态学报. 2018, 38(13): 4744-4751
- Acta Ecologica Sinica. 2018, 38(13): 4744-4751
- http://dx.doi.org/10.5846/stxb201707131269
-
文章历史
- 收稿日期: 2017-07-13
- 网络出版日期: 2018-03-16





2. 西北农林科技大学水土保持研究所, 杨凌 712100;
3. 中国科学院大学, 北京 100049
2. Institute of Soil and Water Conservation, Northwest Agriculture and Forestry University, Yangling 712100, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China
蒸腾是森林水分循环的关键过程, 在一定程度上反映树木对气候因子和土壤水分条件的响应特征。在黄土高原半湿润半干旱地区, 降雨量较少, 水分供需问题十分突出, 不同植被类型和植物种形成了对水分利用的适应性策略。不同乔灌木种对水分的利用特征是合理规划和确定植被建设的生态学基础[1-2]。因此, 对主要植被类型及其建群种的蒸腾耗水特性进行研究显得尤为重要而紧迫。
测定树干木质部液流是研究树木单株和林分蒸腾耗水的重要方法。研究表明, 树干木质部边材液流量占整株树蒸腾的99.8%[3]。Granier热扩散探针法(TDP)由于其测定精度高、操作简便、可连续实时监测和成本较低等优点而被广泛应用。近年来, 众多学者采用此技术对不同树种树干液流进行了研究, 如马占相思[4]、侧柏[5]、油松[6]等, 分析了这些树种的液流动态特征和与气象因子的关系。关于黄土高原地区一些常见树种如辽东栎、油松、栓皮栎、刺槐、柠条锦鸡儿等液流特征也有报道[7-12], 这些研究多集中于对单一树种树干液流特征的探讨和林分蒸腾耗水量的测算分析。在群落内, 树木个体之间相互依存互为影响, 并存在对于资源的竞争性利用。优势种和伴生种生态位有所不同, 林木接收地上(如光照)和地下部资源也有差异, 进而会影响其蒸腾过程。这些差异可能会表现为树干液流特征的差异。关于森林优势种和伴生种树干液流特征的比较研究目前还少有报道, 特别是针对天然林群落内的研究还极为少见[13], 而这方面的研究是了解群落内个体间特别是种间相互关系的重要内容。
辽东栎(Quercus liaotungensis)是黄土丘陵区主要天然次生林群落的优势树种, 具天然更新能力, 对水土保持及维持群落稳定性和区域生态平衡具有重要作用。本研究采用热扩散探针法连续监测黄土丘陵区天然次生林群落优势种辽东栎及3个主要伴生种(山杏、细裂槭和桃叶卫矛)的树干边材液流, 通过分析液流通量密度的日变化特征及其对主要环境因子的响应, 探究优势种和伴生种树干液流动态变化特征的差异, 从而为揭示该地区主要树种蒸腾耗水对环境条件的响应机制提供理论支撑。
1 材料与方法 1.1 研究地概况及供试材料的选取研究地点位于陕西省延安市南郊公路山林地, 地理坐标为36°25.40′N, 109°31.53′E, 海拔1245—1395 m。该地区位于森林区与森林草原区的过渡地带, 为典型森林分布区的北缘[2, 14], 地貌类型属于黄土高原丘陵沟壑区, 气候为温带半干旱气候。主要土壤类型为黄绵土, pH 7.8—8.5。天然森林类型以栎类林为主, 也有少量松、柏、杨、桦林。辽东栎林为该地区顶极森林群落。
设有辽东栎天然次生林固定监测样地面积为0.04 hm2, 坡向东北, 坡度22°, 植被覆盖率约为75%。优势种辽东栎和主要伴生种山杏为主要建群种, 其他伴生树种有侧柏、桃叶卫矛、大果榆和细裂槭。林下灌木主要有小叶锦鸡儿、紫丁香、水栒子等。林分年龄约为60年。本研究用于监测树干液流的辽东栎和主要伴生树种山杏各3株, 另外2个伴生树种细裂槭和桃叶卫矛各1株, 各供试树木个体基本参数见表 1。
| 树种 Tree species |
样木号 Tree No. |
树高 Height/cm |
胸径 Diameter/cm |
树皮厚 Bark thickness/cm |
边材厚 Sapwood depth/cm |
| 辽东栎Quercus liaotungensis | 1 | 7.94 | 17.55 | 1.0 | 2.5 |
| 2 | 9.2 | 18.76 | 1.3 | 2.1 | |
| 3 | 8.35 | 17.75 | 1.19 | 2.38 | |
| 山杏Armeniaca sibirica | 4 | 5.46 | 12 | 0.92 | 1.03 |
| 5 | 6.8 | 12.7 | 0.95 | 1.04 | |
| 6 | 6.45 | 12.45 | 0.94 | 1.04 | |
| 细裂槭Acer stenolobum | 7 | 5.8 | 10.3 | 0.83 | 0.98 |
| 桃叶卫矛Euonymus bungeanus | 8 | 4.8 | 9.1 | 0.77 | 0.84 |
树干边材液流利用Granier热扩散探针进行测量。根据前期研究基础, 本研究统一采用10 mm长探针, 直径为2 mm。探针统一安装于树干北侧1.3 m高度, 每组2个探针(加热探针和参考探针)上下间隔12 cm左右。采用专用电流调节装置连接12 V直流电源, 供给加热探针以恒定电流保持0.15 W的持续加热。为避免太阳辐射所产生的测量误差, 将探针用防辐射铝铂覆盖。用数据采集器(CR1000, Campbell Scientific)每30 s读取一次数据, 并记录30 min的平均值。
树干边材液流通量密度(Fd, mL m-2 s-1)根据监测得到的两探针间的温度差, 利用Granier经验公式进行计算如下[15]:
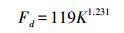
|
其中, K=(ΔTm-ΔT)/ΔT, ΔTm为液流通量为零时加热探针与参考探针间的最大温度差, ΔT为瞬时温差值。
1.3 环境因子测定在试验地林外空旷处设有主要气象要素监测装置, 对太阳辐射(Rs, W/m2)、空气温度(T, ℃)、空气相对湿度(RH, %)、降雨(P, mm)进行实时监测。其中, 太阳辐射利用光量子传感器(LI-200, Li-Cor)进行测量;空气温度和相对湿度利用温湿度计(HMP50 Vaisala, Finland)进行测量;降水量用翻斗式自计雨量计(Davis Rain collector II model 7852, Davis Instruments, USA)测量。其中, 光量子传感器和温湿度计安装于地面2 m以上处;翻斗式自计雨量计水平固定于地面上30 cm处。利用CR1000数据采集器连接各传感器, 数据记录频率同液流测定。样地内设置3个时域反射仪(TDR)水分测管, 每月测定2—3次土壤含水量。空气水汽压亏缺(VPD, kPa)是大气温度与空气相对湿度的综合反映, 根据Campbell和Norman的公式计算如下[16]:
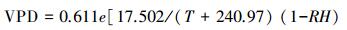
|
式中, T为空气温度(℃), RH为大气相对湿度(%)。
1.4 数据选取与分析本文分析了2011年5月所监测的数据, 两个土壤水分状况时段的选取依据为TDR测定的土壤含水量和对于降雨事件的监测。在监测期间的5月9日有一次明显的降雨事件, 选取5月7日和11日分别代表土壤水分状况相对干旱和湿润的时段, 并且为典型天气条件(晴天或多云), 因此可用于树干液流通量密度的日变化规律分析。对优势种辽东栎及伴生种每30 min的液流通量密度和气象环境因子(太阳辐射和水汽压亏缺)的关系分析涵盖5月份全月数据, 采用SPSS 20.0和SigmaPlot 12.5对数据进行统计分析和做图。
2 结果与分析 2.1 测定期间气象环境因子日变化特征图 1为研究区2011年5月份逐日降水量、平均太阳总辐射、平均空气温度和水汽压亏缺。该月份总降水量为74.17 mm, 一次相对较大的降雨事件出现在5月9日, 日降水量达到40.3 mm。在此之前的3月份和4月份, 是雨水较为缺乏的月份, 两月份总降水量只有28.2 mm, 相对干旱。日均太阳总辐射与天气状况有关, 其最大值(晴天)约为23.87 MJ m-2 d-1, 符合该地区的季节变化。空气温度和水汽压亏缺也是季节变化和当日天气状况的综合体现。总体上, 日均太阳总辐射, 空气温度和水汽压亏缺保持相同的变化趋势。

|
| 图 1 测定期间(2011年5月份)研究区降水量、平均太阳总辐射、空气温度与水汽压亏缺的逐日变化 Fig. 1 Daily changes of precipitation, mean solar radiation, air temperature, and vapor pressure deficit at the study site during the study period of May 2011 |
图 2为2011年5月7日(降雨前相对干旱时期)、5月11日(降雨后相对湿润时期)优势种辽东栎及3个伴生树种(山杏、细裂槭、桃叶卫矛)树干液流通量密度以及主要环境因子的日变化曲线。在典型天气(晴天或多云), 优势种与伴生种树干液流通量密度日变化基本上呈现出与气象环境因子相同的变化趋势。在中午前后出现高峰, 夜间接近于零。优势种辽东栎液流启动时间较早, 通常在5:00左右, 伴生树种山杏、细裂槭和桃叶卫矛液流启动时间均晚于辽东栎一个小时以上。辽东栎于9:00前后达到峰值, 细裂槭、桃叶卫矛和山杏液流通量密度达到峰值的时间分别在10:00、11:00和13:00前后。与辽东栎和山杏相比, 细裂槭和桃叶卫矛液流通量密度日变化曲线峰值较窄, 可能与林内光照限制有关。

|
| 图 2 相对干旱(5月7日)和相对湿润(5月11日)条件下各样木液流通量密度日变化 Fig. 2 Diurnal variation of sap flux densities in the four species in May 7 and May 11 |
在降雨前后两种土壤水分状态下液流通量密度日变化特征表现有所不同, 不同树种间存在差异。因5月11日的太阳辐射和空气水汽压亏缺略低于5月7日, 优势种辽东栎的液流通量密度日变化曲线峰值也呈现出5月11日略低于5月7日, 但在前后两日基本上较为接近, 无明显变化。而3个伴生树种的液流通量密度日变化曲线峰值在5月11日却明显高于5月7日, 显示其在土壤水分充足的条件下液流通量密度明显增大。表明优势种辽东栎的液流通量密度对天气状况(太阳辐射和空气水汽压亏缺)响应明显, 而3个伴生树种的液流通量密度对相对干旱和湿润两种土壤水分条件的响应更为明显。
2.3 树干液流通量密度与气象环境因子的关系分析液流通量密度与2个主要气象环境因子(太阳辐射和空气水汽压亏缺)的Pearson相关分析表明, 各树种液流通量密度与太阳辐射和空气水汽压亏缺均呈显著正相关(表 2)。各树种树干液流通量密度同太阳辐射的相关系数都略大于同水汽压亏缺的相关系数。除此之外, 优势种辽东栎液流通量密度与气象因子的相关性大于几个伴生树种, 伴生树种对气象环境因子的响应相对较弱。
| 树种 Tree species |
样木号 Samples No. |
太阳辐射 Solar radiation |
水汽压亏缺 Vapor pressure deficit |
| 辽东栎 Q. liaotungensis |
1 | 0.796** | 0.687** |
| 2 | 0.834** | 0.782** | |
| 3 | 0.804** | 0.745** | |
| 山杏 A. sibirica |
4 | 0.629** | 0.516** |
| 5 | 0.693** | 0.554** | |
| 6 | 0.645** | 0.532** | |
| 细裂槭 A. stenolobum |
7 | 0.570** | 0.459** |
| 桃叶卫矛 E. bungeanus |
8 | 0.484** | 0.367** |
| **表示显著性水平为P < 0.01 | |||
本研究中优势树种辽东栎与3个伴生树种表现出基本相同的树干液流通量密度日变化趋势, 符合主要气象环境因子特别是光照因子的昼夜规律性, 这与该地区其他树种的研究报道也基本一致, 例如侧柏[5]、旱柳[17]、苹果树[18-19]等。说明在日尺度上, 作为蒸腾耗水监测指标的树干液流变化与叶片气孔开闭密切关联, 关键影响因子应该是气象环境因子。太阳辐射和空气水汽压亏缺对树干液流的影响归根到底是对蒸腾的影响, 树木蒸腾随着太阳辐射和空气水汽压亏缺的增强而增强, 树干液流通量密度也随之增大。太阳辐射和水汽压亏缺共同驱动, 二者协同影响液流通量密度的大小, 但在特殊情况下也不完全是正相关关系, 例如在光合午休期间, 虽然太阳辐射和水汽压亏较高, 但由于气孔导度的降低, 蒸腾会在短时间内下降。
树干液流的启动是树干内水分传输的启动, 主要由气孔蒸腾驱动。清晨, 随着太阳辐射的升高, 气孔导度相应升高, 光合和蒸腾速率逐渐增强, 水分通过根部以被动方式吸入体内, 产生的蒸腾拉力驱动液流启动。但优势树种和伴生树种在树干液流启动时间上存在明显的差异, 优势种明显早于伴生种一个小时以上, 达到峰值的时间也较其他树种早, 说明优势种在空间上占据优越位置, 具有对光能利用的明显优势, 气孔反应等生理过程与太阳辐射的时滞小, 树干液流启动和达到峰值的时间也相对较早。由于群落对光能进行了合理的限制性再分配, 对树干液流的变化节律产生一定的驱动作用[9-10]。伴生种通常在群落内优势度低, 有的个体甚至为被压木, 在光能利用方面处于弱势, 体现在响应特征方面相关性较低(表 2)。另外, 优势种与伴生种的液流动态差异也可能与树种本身的生理特性有关。于萌萌等[20]对紫椴、色木槭和红松树干液流进行研究发现, 树木之间液流不同的原因与木质部解剖结构的差异及植物本身的生理特性有关, 这些因素会导致不同植物在水分消耗能力上的差异。此外, 液流通量密度在同一树种之间也存在差异, 这与树木个体之间的差异有关, 例如树木的直径、树高、叶面积以及所处的的生长阶段。
作为树木蒸腾的水分来源, 土壤水分状况对树干液流应该具有一定程度的影响。降雨补充土壤水分后同一个体的树干液流通量密度有所升高, 符合一般规律。但伴生树种表现出较大的差异, 可能与伴生树种所处的林内微气象环境有关。伴生种在群落内的空间劣势和在相应的微气象环境下叶部较少启动气孔调节机制, 较好的土壤水分条件对蒸腾作用促进明显。而优势种在昼间会随气象因子的变化启动气孔调节功能, 随土壤水分条件的变化不太剧烈。已有研究表明, 乡土树种辽东栎对土壤水分条件的变化敏感度低, 通过气孔调节实现对水分的保守性利用[21]。优势种对于群落的稳定性应该具有主导作用, 辽东栎对于水分的保守性利用有利于伴生树种的生存和群落物种多样性的维持。伴生树种的水分利用特点应该是其适应空间结构特征的综合反应, 特别是对光能的合理利用。
4 结论本文通过对黄土丘陵区辽东栎天然次生林群落优势种和3个主要伴生种树干液流特征对主要气象因子和土壤水分状况的响应分析, 得到以下结论:
(1) 辽东栎、山杏、细裂槭和桃叶卫矛液流通量密度日变化动态特征总体上与气象环境因子变化的昼夜规律性保持一致, 呈单峰或双峰型曲线。
(2) 优势种辽东栎与伴生种液流通量密度日变化规律存在一定的差异。辽东栎液流启动时间较早, 而伴生种液流启动时间均晚于辽东栎1 h以上。辽东栎达到峰值的时间也较3个伴生种早。伴生种细裂槭和桃叶卫矛液流通量密度日变化曲线峰值较窄。
(3) 优势种辽东栎树干液流通量密度随土壤水分条件的变化不太剧烈, 而伴生种随土壤水分条件的变化受到的影响程度相对较大。
(4) 相关分析表明, 树干液流通量密度与太阳辐射和空气水汽压亏缺均呈极显著正相关, 优势种的相关性大于伴生种。
| [1] | 吴钦孝, 杨文治. 黄土高原植被建设与持续发展. 北京: 科学出版社, 1998. |
| [2] | 杜盛, 刘国斌. 黄土高原植被恢复的生态功能. 北京: 科学出版社, 2015. |
| [3] | Granier A, Bobay V, Gash J H C, Gelpe J, Saugier B, Shuttleworth W J. Vapour flux density and transpiration rate comparisons in a stand of Maritime pine (Pinus pinaster Ait.) in Les Landes forest. Agricultural and Forest Meteorology, 1990, 51(3/4): 309–319. |
| [4] | 马玲, 饶兴权, 赵平, 陆平, 蔡锡安, 曾小平. 马占相思整树蒸腾的日变化和季节变化特征. 北京林业大学学报, 2007, 29(1): 67–73. |
| [5] | 于占辉, 陈云明, 杜盛. 黄土高原半干旱区侧柏(Platycladus orientalis)树干液流动态. 生态学报, 2009, 29(7): 3970–3976. |
| [6] | 张建国, 久米朋宣, 大规恭一, 山中典和, 杜盛. 黄土高原半干旱区辽东栎的树干液流动态. 林业科学, 2011, 47(4): 63–69. DOI:10.11707/j.1001-7488.20110411 |
| [7] | 聂立水, 李吉跃, 翟洪波. 油松、栓皮栎树干液流速率比较. 生态学报, 2005, 25(8): 1934–1940. |
| [8] | 吴芳, 陈云明, 于占辉. 黄土高原半干旱区刺槐生长盛期树干液流动态. 植物生态学报, 2010, 34(4): 469–476. |
| [9] | 于占辉, 陈云明, 杜盛. 黄土高原半干旱区人工林刺槐展叶期树干液流动态分析. 林业科学, 2009, 45(4): 53–59. DOI:10.11707/j.1001-7488.20090409 |
| [10] | 夏永秋, 邵明安. 黄土高原半干旱区柠条(Caragana korshinskii)树干液流动态及其影响因子. 生态学报, 2008, 28(4): 1376–1382. |
| [11] | Yan M J, Zhang J G, He Q Y, Shi W Y, Otsuki K, Yamanaka N, Du S. Sapflow-based stand transpiration in a semiarid natural Oak forest on China's Loess Plateau. Forests, 2016, 7(10): 227. |
| [12] | 刘潇潇, 李国庆, 闫美杰, 杜盛. 黄土高原主要树种树干液流研究进展. 水土保持研究, 2017, 24(3): 369–373. |
| [13] | 万艳芳, 于澎涛, 刘贤德, 王顺利, 王彦辉, 熊伟. 祁连山青海云杉树干液流密度的优势度差异. 生态学报, 2017, 37(9): 3106–3114. |
| [14] | Tsunekawa A, Liu G B, Yamanaka N, Du S. Restoration and Development of the Degraded Loess Plateau, China. Tokyo: Springer, 2014. |
| [15] | Granier A. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 1987, 3(4): 309–320. DOI:10.1093/treephys/3.4.309 |
| [16] | Campbell G S, Norman J M. An Introduction to Environmental Biophysics. New York: Springer Science & Business Media, 1998. |
| [17] | 彭小平. 水蚀风蚀交错区不同立地条件下柳树树干液流差异研究[D]. 杨凌: 西北农林科技大学, 2014. |
| [18] | 孟秦倩, 王健, 张青峰, 吴发启. 黄土山地苹果树树体不同方位液流速率分析. 生态学报, 2013, 33(11): 3555–3561. |
| [19] | 王力, 王艳萍. 黄土塬区苹果树干液流特征. 农业机械学报, 2013, 44(10): 152-158, 151-151. DOI:10.6041/j.issn.1000-1298.2013.10.024 |
| [20] | 于萌萌, 张新建, 袁凤辉, 何秀, 关德新, 王安志, 吴家兵, 金昌杰. 长白山阔叶红松林三种树种树干液流特征及其与环境因子的关系. 生态学杂志, 2014, 33(7): 1707–1714. |
| [21] | Du S, Wang Y L, Kume T, Zheng J G, Otsuki K, Yamanaka N, Liu G B. Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agricultural and Forest Meteorology, 2011, 151(1): 1–10. DOI:10.1016/j.agrformet.2010.08.011 |