 2018, Vol. 38
2018, Vol. 38文章信息
- 王璐, 杨海军, 李昆, 李玲, 南晓飞, 张振兴.
- WANG Lu, YANG Haijun, LI Kun, LI Ling, NAN Xiaofei, ZHANG Zhenxing.
- 长白山源头溪流底栖动物群落结构季节动态
- Seasonal dynamics of macroinvertebrate community structure in a headwater stream in the Changbai Mountains
- 生态学报. 2018, 38(13): 4834-4842
- Acta Ecologica Sinica. 2018, 38(13): 4834-4842
- http://dx.doi.org/10.5846/stxb201707121259
-
文章历史
- 收稿日期: 2017-07-12
- 网络出版日期: 2018-03-16

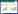




源头溪流是河湖水系的重要组成部分[1-2],其通常具有独特的地形地貌、高度异质性的生态环境,且时空变化显著[3],能为多样化的水生生物群落提供栖息地[4],对维持整个流域的结构和功能健康具有重要的意义[5-6]。底栖动物作为河流生态系统中分布最广[7]、多样性最高的生物类群[8],处于河流生态系统食物链的中间环节,在河流生态系统物质循环和能量流动中具有不可替代的作用[9-10],底栖动物的群落结构及季节动态特征更是源头溪流生物学和生态学研究的基础。
当前,我国对底栖动物的研究主要集中于各流域干流底栖动物的时空分布特征及水质生物学评价方面[11-13],涉及源头溪流底栖动物群落结构的研究鲜见报道[14-15]。同时在北方地区的冬季结冰期,取样条件的限制制约了溪流底栖动物群落结构季节动态的研究。长白山源头溪流作为松花江水系的源头,对维持松花江流域结构和功能的稳定具有重要作用,但溪流生态学研究匮乏,关于底栖动物季节动态的研究尚未见报道。
作为长白山源头溪流底栖动物长期调研工作的一部分,本研究旨在了解2016—2017年度长白山源头溪流底栖动物群落结构和物种组成,揭示底栖动物群落结构的季节动态特征及其主要的环境驱动因子,丰富水域生态学关于寒冷区的基础研究,并为深入开展相关工作奠定一定的理论基础;在应用上,有望为长白山生态系统的健康维护和管理,及松花江水系乃至全国类似水系的水生生物保护及生态修复提供基础数据和参照标准。
1 研究地概况及研究方法 1.1 研究地概况试验溪流位于吉林省通化市辉南县龙湾国家级自然保护区内(42°16′20″—42°26′57″N, 126°13′55″—126°32′02″E),地处长白山北麓龙岗山脉中段。该地区气候属于北温带大陆性季风气候,冬季寒冷漫长,历年平均气温4.1℃,最低温出现在1月份,年平均降水量704.2 mm,降水多集中在夏季。本研究所选区域内河流水系属于松花江支流,数量众多的源头溪流分别汇入南河和北河,之后交汇为后河,再依次汇入三统河、辉发河,最终汇入松花江。每年11月下旬至翌年4月上旬为溪流结冰期,约70%的水面被冰层覆盖[16]。冬季冰面下水体仍维持流动性,冰下水体温度约为0.5℃。
本研究所选河段位于上述南河小流域源头、吊水壶风景区溪流上游,该河段无人为干扰及旅游开发,能较好地反应自然状态下长白山源头溪流的生态特征。研究地海拔700—800 m,流域森林覆盖率大于75%,河岸带以色木槭(Acer mono)、蒙古栎(Quecus mongolica)、山杨(Populus davidiana)、榆树(Ulmus pumila)、紫椴(Tilia amurensis)等为主要优势植被。由于源头溪流生境异质性强,为充分反映溪流的生境条件及生物状况,本研究沿水流方向共设置5个采样断面,其中3个断面位于一级支流,两个断面位于二级溪流,断面长100 m,间距不小于500 m(图 1)。

|
| 图 1 试验断面布设图 Fig. 1 Location of experimental river reach |
分别于2016年6月30日(夏季,丰水期)、2016年10月22日(秋末,平水期)、2016年11月24日(冬季,枯水期,河面冻结)及2017年4月2日(初春,枯水期,河面开始融化)用40目网径、采样面积0.09 m2的索伯网(Surber sampler)沿每个断面采集3次。冬季由于河面冻结,先用锤子破开冰面,再进行采样。采集的样品用60目网筛过滤清洗后装入封口袋内,并加入75%酒精固定保存。在实验室内,将底栖动物挑拣出,放入50 mL的标本瓶中,加入75%酒精固定。通过体式显微镜对底栖动物进行鉴定、计数,样品尽可能鉴定至最低分类单元,并参照Barbour等[17]的分类方法划分底栖动物的功能摄食类群。
1.3 水体理化指标测定采样同期测定水文指标(河宽、水深、流速)、水质指标(水温、溶解氧、电导率)、河床凋落叶分布及底质类型。河宽使用米尺测定,水深使用水深探测棒测定,流速使用便携式流速分析仪测定断面0.6倍水深处的流速。水质指标使用便携式水质分析仪(哈纳HI9829T)测定。测定试验河段内凋落叶斑块的堆积面积,以斑块堆积面积占河床总面积的百分比表征河床凋落叶分布情况。溪流底质类型按照Cummins的划分方法[18]记录。
1.4 数据处理与分析采用物种数、密度、Shannon-Wiener多样性指数、改进的Shannon-Wiener多样性指数[19]、Margalef多样性指数、Pielou均匀度指数及各功能摄食类群相对丰度、密度及物种丰富度来表征底栖动物群落结构的季节变化。
选用SPSS 21.0中的One-way ANOVA对4次取样的水体理化指标及底栖动物数据进行差异显著性检验,若方差不齐,选用非参数检验方法检验差异显著性,显著性水平取0.05。各样点底栖动物物种相似性聚类分析采用PRIMER 5进行,非度量多维标度(NMDS)分析采用Past 3进行。使用SPSS 21.0对环境因子与底栖动物群落结构参数间进行相关分析,以明确不同环境因子的季节变化对底栖动物群落结构的影响。
2 结果与分析 2.1 环境参数试验期间测定的水环境参数见表 1,单因素方差分析结果显示水深在夏、秋季显著高于冬、春季节;河宽、流速和水温表现为冬季显著低于其他季节;凋落叶分布在季节间差异显著,表现为秋季>冬季>春季>夏季。溪流底质以卵石和圆石为主,底质组成季节间变化不大。
| 参数 Parameters |
夏季 Summer |
秋季 Autumn |
冬季 Winter |
春季 Spring |
| 平均水深Mean water depth/cm | 20.02±7.81a | 19.5±5.54a | 9.9±1.33b | 10.38±1.54b |
| 平均河宽Mean water width/m | 2.68±0.88a | 2.2±0.90a | 1.2±0.41b | 1.64±0.30a |
| 流速Velocity/(m/s) | 0.39±0.15a | 0.37±0.11a | 0.15±0.07b | 0.17±0.07a |
| 水温Water temperature/℃ | 9.9±1.67a | 8.84±1.11a | 0.6±0.37b | 2.02±0.31a |
| 电导率Conductivity/(μS/cm) | 0.14±0.01a | 0.14±0.01a | 0.14±0a | 0.14±0.02a |
| 溶解氧Dissolved oxygen/(mg/L) | 15.48±0.33a | 15.62±0.45a | 14.82±0.35a | 14.74±0.47a |
| pH | 7.36±0.09a | 7.54±0.11a | 7.28±0.19a | 7.48±0.04a |
| 凋落叶分布Detritus/% | <1 | 60—80 | 45—50 | 5—10 |
| 底质Substrate/% | ||||
| 细砂Sand/% | 5 | 10 | 10 | 10 |
| 沙砾Gravel/% | 10 | 15 | 20 | 15 |
| 卵石Pebble/% | 30 | 35 | 25 | 20 |
| 圆石Cobble/% | 45 | 40 | 40 | 45 |
| 漂砾Boulder/% | 10 | 10 | 5 | 10 |
| 同行不同字母表示差异显著(P<0.05) | ||||
本研究共采集并鉴定出底栖动物15395头,90个分类单元,隶属于3纲9目38科。其中,水生昆虫占绝对优势,共85属,占总分类单元的93.4%,包括蜉蝣目14属、襀翅目14属、毛翅目21属、双翅目32属,鞘翅目3属和蜻蜓目1属;寡毛类4属,占总分类单元的4.96%;甲壳类2属,占总分类单元的2.19%。
4次取样底栖动物各类群组成及数量见表 2。单因素方差分析结果显示,底栖动物物种数目和密度在不同季节间差异显著。夏季和秋季物种数目显著高于冬季和春季,秋季密度显著高于其他季节。除襀翅目外,各类群密度总体上表现为秋季最高。
| 物种 Taxa |
总计 Total |
夏季 Summer |
秋季 Autumn |
冬季 Winter |
春季 Spring |
| 蜉蝣目Ephemeroptera | 14 (1453.33) | 12 (148.15b) | 13 (891.85a) | 5 (62.96b) | 7 (350.37b) |
| 襀翅目Plecoptera | 14 (1800.00) | 12 (171.11c) | 11 (564.44ab) | 3 (733.33a) | 8 (331.11bc) |
| 毛翅目Trichoptera | 21 (1251.11) | 16 (466.67a) | 20 (446.67a) | 6 (292.59a) | 6 (45.19b) |
| 双翅目Diptera | 32 (1864.44) | 30 (385.19ab) | 23 (695.56a) | 19 (277.04b) | 13 (506.67ab) |
| 鞘翅目Coleoptera | 3 (1823.70) | 3 (358.52bc) | 3 (860.00a) | 3 (537.04b) | 1 (68.15c) |
| 蜻蜓目Odonata | 1 (2.22) | 1 (0.74a) | 1 (1.48a) | — | — |
| 甲壳类Crustacea | 4 (2973.33) | 1 (611.11b) | 1 (2142.22a) | 1 (154.81b) | 1 (65.19b) |
| 寡毛类Oligochaeta | 1 (235.56) | 4 (193.33a) | 3 (42.22b) | — | — |
| 总计Total | 90(2850.93) | 79(2334.81b) | 75(5644.44a) | 32(2057.78b) | 36 (1366.67b) |
季节变化对底栖动物群落结构的影响,可反映在多样性指数的变化上(图 2)。单因素方差分析结果显示除Shannon-Wiener多样性指数外,各指数在不同季节间差异显著。多样性整体表现为夏季和秋季高于冬季和春季,其中Margalef多样性指数还表现为春季显著高于冬季。而Pielou均匀度指数与多样性指数变化相反,表现为冬季和春季高于夏季和秋季。

|
| 图 2 不同季节底栖动物Shannon-Wiener多样性指数、改进的Shannon-Wiener多样性指数、Margalef多样性指数和Pielou均匀度指数 Fig. 2 The Shannon-Wiener diversity index, Improved Shannon-Wiener diversity index, Margalef diversity index and Pielou evenness index of macroinvertebrate in different seasons |
物种组成的Cluster和NMDS(stress value=0.08)见图 3和图 4,结果表明夏季和秋季底栖动物群落结构相似性较高,而冬、春季底栖动物群落结构与其差异较大。

|
| 图 3 各季节底栖动物群落聚类图 Fig. 3 The hierarchical cluster dendrogram of the macroinvertebrates communities at different seasons |

|
| 图 4 底栖动物群落结构无度量多维标定法排序 Fig. 4 NMDS distribution of the macroinvertebrate assemblages |
功能摄食类群可以进一步从生态系统功能方面揭示底栖动物的季节变化。研究发现长白山源头溪流收集者相对丰度最高,达到44.22%,其次为撕食者和捕食者,相对丰度分别为25.18%和16.71%,刮食者和滤食者相对丰度较低,仅占8.83%和1.85%。
季节变化对功能摄食类群的影响可反映在各类群的相对丰度、密度及物种丰富度的变化上。相对丰度的季节动态分析结果表明,除撕食者显著增加外,秋季各类群相对丰度与夏季无显著差异。而进入冬季后,各类群相对丰度变化显著,冬、春季撕食者及刮食者相对丰度显著降低,此外,冬季捕食者和滤食者相对丰度显著升高,而春季收集者相对丰度显著升高。
功能摄食类群密度和物种丰富度的季节动态见图 5,结果表明撕食者和刮食者密度表现为秋季最高,夏季次之,冬、春季较低;收集者密度表现为秋季>春季>夏季>冬季;捕食者密度表现为冬季最高,秋、春季次之,夏季最低;滤食者密度在各季节间差异不显著。物种丰富度方面,各功能摄食类群总体上表现为夏、秋季高于冬季和春季,其中,撕食者、收集者和捕食者3种优势功能摄食类群的物种丰富度均在冬季显著降低,并达到最低值。

|
| 图 5 不同季节间各功能摄食类群密度及物种丰富度的比较 Fig. 5 Comparison of functional feeding groups density and richness among different seasons FC:滤食者Filter-Collector; GC:收集者Gather-Collector; SC:刮食者Scraper; PR:捕食者Predator; SH:撕食者Shredder |
为了解不同环境因子对底栖动物群落结构季节动态的影响,将群落结构参数分别与河宽、水深、流速、水温、电导率、溶解氧、凋落叶分布及各底质类型百分比间进行相关分析,找出显著相关的因子(表 3)。结果表明物种数、Shannon-Wiener多样性指数、改进的Shannon-Wiener多样性指数、Margalef多样性指数、均匀度指数、撕食者相对丰度、捕食者相对丰度、刮食者相对丰度及撕食者密度受温度影响显著;密度和撕食者密度与凋落叶分布显著相关;物种数、改进的Shannon-Wiener多样性指数、Margalef多样性指数、均匀度指数、撕食者相对丰度、捕食者相对丰度及撕食者密度还受流速影响显著。由此可知,水温、凋落叶分布及流速是长白山源头溪流底栖动物群落结构季节动态的主要环境驱动因子。
| 参数 Parameters |
物种数目 Species number |
密度 Diversity |
Shannon-Wiener多样性指数 Shannon-Wiener diversity index |
改进的Shannon-Wiener多样性指数 Improved Shannon-Wiener diversity index |
Margalef多样性指数 Margalef diversity index |
Pielou均匀度指数 Pielou evenness index |
撕食者 SH/% |
捕食者 PR/% |
刮食者 SC/% |
撕食者密度 Density of SH |
| 水温 Water temperature |
0.961** | — | 0.495* | 0.699** | 0.977** | -0.554* | 0.788** | -0.713** | 0.458* | 0.558* |
| 凋落叶分布 Detritus/% |
— | 0.596** | — | — | — | — | — | — | — | 0.564** |
| 流速 Velocity |
0.733** | — | — | 0.499* | 0.730** | -0.486* | 0.684** | -0.529* | — | 0.538* |
| *表示0.05水平上的相关性;**表示0.01水平上的相关性; 未列入的环境参数表示与所有的群落结构参数相关性均不显著; “—”表示不具有统计学意义 | ||||||||||
源头溪流对维持生物多样性具有重要意义,底栖动物作为源头溪流生态系统的重要组成部分,近些年受到了越来越多的关注[20-22]。本研究首次对冬季长白山源头溪流底栖动物进行了取样,并分析了底栖动物群落结构及季节动态特征,可为长白山松花江水系可持续管理及生态修复提供基础信息。
3.1 长白山溪流底栖动物群落结构特征关于长白山底栖动物群落结构的研究,目前仅见于立等[23]对长白山水生昆虫的调查研究(共鉴定出蜉蝣目14属、襀翅目12属、毛翅目19属、双翅目36属,鞘翅目1属、蜻蜓目3属及脉翅目1属),虽取样点未涵盖本研究地,但同属长白山松花江水系源头溪流,仍可作为历史数据用于分析和对比。本研究采集到的水生昆虫种类和组成(蜉蝣目14属、襀翅目14属、毛翅目21属、双翅目32属,鞘翅目3属和蜻蜓目1属)与历史数据差异不大,水生昆虫资源完整,表明近20年研究地的生境条件维持良好。
研究地处于保护区内林区溪流源头,河岸林茂盛,上游坡度大,水流湍急,河床以卵石或圆石为主,均有利于底栖动物栖息。研究地水体pH在7.2—7.8之间,属于中碱水体,适宜大多数水生昆虫代谢和发育。水体溶解氧含量高,多处于饱和状态。与历史数据[23]类似,采集到的水生昆虫也多为喜清水和喜凉的山地溪流石栖种类。试验溪流属松花江水系源头,其底栖动物物种组成与下游松花江干流[24](水生昆虫63.8%、软体动物21.6%、寡毛类12.1%、甲壳类2.6%,多为耐污种)差异显著,底栖动物密度和物种多样性也远高于干流(密度:5.3—353.3个/m2;Shannon-Wiener多样性指数:0.227—1.461;Margalef多样性指数:1.18—1.81)。物种组成及多样性的差异表明源头溪流最适宜底栖动物生存,也反映了研究地对维持松花江水系生物多样性具有重要的意义。
底栖动物对生境条件变化敏感,在不同水体中的季节动态也不相一致,密度和多样性的最高值可以出现在秋季、冬季[25-26],也可以出现在春季、夏季[27-28]。长白山源头溪流底栖动物群落结构也存在显著的季节差异,冬、春季底栖动物群落结构与夏、秋季差异显著,且物种丰富度和多样性显著降低,表明夏季和秋季的生境条件更适合底栖动物生存。在功能摄食类群方面,密度和物种丰富度是各功能摄食类群实际分布情况的反映,相对丰度能够反映群落功能特征的稳定性。各类群密度整体上表现为秋季最高,表明秋季生境条件最适宜底栖动物生存和繁殖;物种丰富度在冬季达到最低,指示了冬季溪流生境条件最差;相对丰度的季节动态反映了夏季和秋季底栖动物功能结构稳定,而冬、春季稳定性下降的特征。
3.2 群落结构的季节动态特征及环境驱动因素底栖动物群落结构季节上的差异,除了与底栖动物自身的生活史特征密切相关外,更主要地受到水环境条件变化的影响。溪流水文特征、水体理化因子等环境特征的季节变化会影响底栖动物的生长、繁殖及种群的演替,最终会影响到整个溪流生态系统的群落结构和功能[25, 29]。本研究结果表明,水温、凋落叶分布和流速是长白山源头溪流底栖动物群落结构季节动态的主要环境驱动因素。
底栖动物属于变温动物,在一定的水温范围内,其生长、繁殖等一切生命活动极大地受到水温的制约[30-32]。大量的研究表明,冬季由于水体低温及冻结,底栖动物的存活率会受到影响[33]。一些底栖动物会以卵的形式越冬,或通过静止、滞育及减少每年的世代数以应对冬季的低温和冻结生境[34-36],进而会降低底栖动物的数量和丰富度。
凋落叶作为源头溪流主要的能量来源[5],是溪流底栖动物生存的关键环境因子,河床分布的凋落叶一方面为底栖动物提供食物来源,另一方面也为底栖动物提供越冬的栖息地和避难所[37-38]。本研究结果也表明底栖动物密度和撕食者密度都显著地受到凋落叶季节分布的影响。底栖动物功能摄食类群中撕食者主要以堆积在河道的凋落叶为食[39-40],秋季凋落叶大量输入,能为撕食者提供充足的食物,其大量繁殖,密度及物种丰富度均达到最高值;夏季虽河床凋落叶分布少,但河岸带新鲜有机物(叶片、花等)的输入能为撕食者提供丰富的营养物质[41],因此撕食者仍能维持较高的密度;而冬季溪流低温和春季河床凋落叶的减少则是制约冬、春季节撕食者生存和繁殖的主要因素。
流速也是影响底栖动物群落结构的重要环境因子[32]。不同物种间最适流速差异很大[42],一般认为底栖动物的物种丰度和密度的最大值均出现在流速0.3—1.2 m/s范围的各种底质中[43]。冬季流速显著降低,会影响适应急流生境的底栖动物生存,进而影响底栖动物的群落结构。此外,流速降低也使得生境条件更为单一和稳定,进而影响底栖动物的均匀度,本研究结果也表明冬、春季虽底栖动物群落结构简单,但Pielou均匀度指数显著增加,指示了长白山源头溪流枯水期生境条件较丰水期和平水期更为稳定的特征。
4 结论本研究揭示了长白山源头溪流底栖动物群落结构的季节动态特征,底栖动物密度及多样性在夏、秋季显著高于冬季和春季,并在冬季达到最低值。环境因子中水温、凋落叶分布和流速是底栖动物群落结构季节动态的主要环境驱动因子。目前我国关于冬季溪流的生态学研究很少,限制了对溪流冬季生态过程的理解和认识。长白山源头溪流冬季冻结期长达近5个月,为研究冬季溪流的生态过程提供了理想的对象。长白山源头溪流底栖动物生活史的关键期(越冬期)河床凋落叶的堆积对底栖动物多样性的影响、底栖动物对冬季凋落叶分解过程的响应规律等科学问题有待进一步研究,研究成果在学科上能丰富水域生态学关于寒冷区溪流的相关理论,具有重要价值。
| [1] | Horton R E. Erosional development of streams and their drainage basins:hydrophysical approach to quantitative morphology. Geological Society of America Bulletin, 1945, 56(3): 275–370. DOI:10.1130/0016-7606(1945)56[275:EDOSAT]2.0.CO;2 |
| [2] | Benda L, Hassan M A, Church M, May C L. Geomorphology of steepland headwaters:the transition from hillslopes to channels. Journal of the American Water Resources Association, 2005, 41(4): 835–851. DOI:10.1111/jawr.2005.41.issue-4 |
| [3] | Townsend C R. The patch dynamics concept of stream community ecology. Journal of the North American Benthological Society, 1989, 8(1): 36–50. DOI:10.2307/1467400 |
| [4] | Meyer J L, Strayer D L, Wallace J B, Eggert S L, Helfman G S, Leonard N E. The contribution of headwater streams to biodiversity in river networks. Journal of the American Water Resources Association, 2007, 43(1): 86–103. DOI:10.1111/j.1752-1688.2007.00008.x |
| [5] | Vannote R, Minshall G W, Cummins K W, Sedell J R, Cushing C E. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(1): 130–137. DOI:10.1139/f80-017 |
| [6] | Wipfli M S, Richardson J S, Naiman R J. Ecological linkages between headwaters and downstream ecosystems:transport of organic matter, invertebrates, and wood down headwater channels. Journal of the American Water Resources Association, 2007, 43(1): 72–85. DOI:10.1111/j.1752-1688.2007.00007.x |
| [7] | Voelz N J, McArthur J V. An exploration of factors influencing lotic insect species richness. Biodiversity & Conservation, 2000, 9(11): 1543–1570. |
| [8] | Strayer D L. Challenges for freshwater invertebrate conservation. Journal of the North American Benthological Society, 2006, 25(2): 271–287. DOI:10.1899/0887-3593(2006)25[271:CFFIC]2.0.CO;2 |
| [9] | Covich A P, Palmer M A, Crowl T A. The role of benthic invertebrate species in freshwater ecosystems:zoobenthic species influence energy flows and nutrient cycling. BioScience, 1999, 49(2): 119–127. DOI:10.2307/1313537 |
| [10] | Thorp J H, Covich A P. Ecology and Classification of North American Freshwater Invertebrates.2nd ed. New York: Academic Press, 2001. |
| [11] | 徐梦珍, 王兆印, 潘保柱, 巩同梁, 刘乐. 雅鲁藏布江流域底栖动物多样性及生态评价. 生态学报, 2012, 32(8): 2351–2360. |
| [12] | 王备新, 杨莲芳, 胡本进, 单林娜. 应用底栖动物完整性指数B-IBI评价溪流健康. 生态学报, 2005, 25(6): 1481–1490. |
| [13] | 蒋万祥, 贾兴焕, 唐涛, 蔡庆华. 底栖动物功能摄食类群对酸性矿山废水的响应. 生态学报, 2016, 36(18): 5670–5681. |
| [14] | 江晶, 温芳妮, 顾鹏, 闫云君. 湖北清江流域胡家溪大型底栖动物群落结构及水质评价. 湖泊科学, 2009, 21(4): 547–555. |
| [15] | 蒋小明, 程建丽, 熊晶, 张鹗, 谢志才. 黄盖湖水系河源区——皤河大型无脊椎动物群落与水质评价. 长江流域资源与环境, 2011, 20(9): 1040–1046. |
| [16] | 《中国河湖大典》编纂委员会. 中国河湖大典——黑龙江、辽河卷. 北京: 中国水利水电出版社, 2014. |
| [17] | 巴伯. 溪流及浅河快速生物评价方案——着生藻类、大型底栖动物及鱼类. 2版. 郑丙辉, 刘录三, 李黎, 译. 北京: 中国环境出版社, 2011. |
| [18] | Cummins K W. An evaluation of some techniques for the collection and analysis of benthic samples with special emphasis on lotic waters. American Midland Naturalist, 1962, 67(2): 477–504. DOI:10.2307/2422722 |
| [19] | 王寿兵. 对传统生物多样性指数的质疑. 复旦学报:自然科学版, 2003, 42(6): 867–868. |
| [20] | Bae M J, Chun J H, Chon T S, Park Y S. Spatio-temporal variability in benthic macroinvertebrate communities in headwater streams in South Korea. Water, 2016, 8(3): 99. DOI:10.3390/w8030099 |
| [21] | Clarke A, Nally R M, Bond N, Lake P S. Macroinvertebrate diversity in headwater streams:a review. Freshwater Biology, 2008, 53(9): 1707–1721. DOI:10.1111/fwb.2008.53.issue-9 |
| [22] | 李金国, 王庆成, 严善春, 姚琴, 乔树亮, 吕跃东, 韩壮行. 凉水、帽儿山低级溪流中水生昆虫的群落特征及水质生物评价. 生态学报, 2007, 27(12): 5008–5018. DOI:10.3321/j.issn:1000-0933.2007.12.009 |
| [23] | 于力, 暴学祥, 云宝琛. 长白山水生昆虫的研究. 水生生物学报, 1997, 21(1): 31–39. |
| [24] | 霍堂斌, 刘曼红, 姜作发, 李喆, 马波, 于洪贤. 松花江干流大型底栖动物群落结构与水质生物评价. 应用生态学报, 2012, 23(1): 247–254. |
| [25] | 蒋万祥, 贾兴焕, 周淑婵, 李凤清, 唐涛, 蔡庆华. 香溪河大型底栖动物群落结构季节动态. 应用生态学报, 2009, 20(4): 923–928. |
| [26] | 闫云君, 李晓宇. 汉江流域上游支流大型底栖动物群落结构特征与生物多样性. 湖泊科学, 2007, 19(5): 585–591. DOI:10.18307/2007.0514 |
| [27] | 周晓, 葛振鸣, 施文彧, 王天厚, 曹丹, 周立晨. 长江口九段沙湿地大型底栖动物群落结构的季节变化规律. 应用生态学报, 2006, 17(11): 2079–2083. DOI:10.3321/j.issn:1001-9332.2006.11.016 |
| [28] | 张敏, 邵美玲, 蔡庆华, 徐耀阳, 王岚, 孔令惠. 丹江口水库大型底栖动物群落结构及其水质生物学评价. 湖泊科学, 2010, 22(2): 281–290. |
| [29] | Vannote R L, Sweeney B W. Geographic analysis of thermal equilibria:A conceptual model for evaluating the effect of natural and modified thermal regimes on aquatic insect communities. The American Naturalist, 1980, 115(5): 667–695. DOI:10.1086/283591 |
| [30] | 彭松耀, 李新正. 乳山近海大型底栖动物功能摄食类群. 生态学报, 2013, 33(17): 5274–5285. |
| [31] | 于婷婷, 徐奎栋. 长江口及邻近海域秋冬季小型底栖动物类群组成与分布. 生态学报, 2013, 33(15): 4556–4566. |
| [32] | 任海庆, 袁兴中, 刘红, 张跃伟, 周上博. 环境因子对河流底栖无脊椎动物群落结构的影响. 生态学报, 2015, 35(10): 3148–3156. |
| [33] | Olsson T I. Overwintering of benthic macroinvertebrates in ice and frozen sediment in a North Swedish river. Ecography, 1981, 4(3): 161–166. DOI:10.1111/eco.1981.4.issue-3 |
| [34] | Danks H V. Life cycles in polar arthropods-flexible or programmed?. European Journal of Entomology, 1999, 96(2): 83–102. |
| [35] | Danks H V. Insect cold hardiness:a Canadian perspective. Cryo Letters, 2000, 21(5): 297–308. |
| [36] | Hanski I. Four kinds of extra long diapause in insects:a review of theory and observations. Annales Zoologici Fennici, 1988, 25(1): 37–53. |
| [37] | Petersen R C, Cummins K W. Leaf processing in a woodland stream. Freshwater Biology, 1974, 4(4): 343–368. DOI:10.1111/fwb.1974.4.issue-4 |
| [38] | Laitung B, Chauvet E. Vegetation diversity increases species richness of leaf-decaying fungal communities in woodland streams. Archiv Für Hydrobiologie, 2005, 164(2): 217–235. DOI:10.1127/0003-9136/2005/0164-0217 |
| [39] | Cummins K W, Wilzbach M A, Gates D M, Perry J B, Taliaferro W B. Shredders and Riparian Vegetation:leaf litter that falls into streams influences communities of stream invertebrates. BioScience, 1989, 39(1): 24–30. DOI:10.2307/1310804 |
| [40] | Haapala A, Muotka T, Laasonen P. Distribution of benthic macroinvertebrates and leaf litter in relation to streambed retentivity:implications for headwater stream restoration. Boreal Environment Research, 2003, 8(1): 19–30. |
| [41] | Kochi K, Asaeda T, Chibana T, Fujino T. Physical factors affecting the distribution of leaf litter patches in streams:comparison of green and senescent leaves in a step-pool streambed. Hydrobiologia, 2009, 628(1): 191–201. DOI:10.1007/s10750-009-9756-2 |
| [42] | 郑文浩, 渠晓东, 张远, 孟伟. 太子河流域大型底栖动物栖境适宜性. 环境科学研究, 2011, 24(12): 1355–1363. |
| [43] | Beauger A, Lair N, Reyes-Marchant P, Peiry J L. The distribution of macroinvertebrate assemblages in a reach of the River Allier (France), in relation to riverbed characteristics. Hydrobiologia, 2006, 571(1): 63–76. DOI:10.1007/s10750-006-0217-x |