 2018, Vol. 38
2018, Vol. 38文章信息
- 贺一鸣, 李青丰, 贺晓, 樊如月, 段茹晖, 刘洋.
- HE Yiming, LI Qingfeng, HE Xiao, FAN Ruyue, DUAN Ruhui, LIU Yang.
- 环境因子对蒙古莸种子萌发及幼苗建成的影响
- Effects of environmental factors on seed germination and seedling establishment of Caryopteris mongolica
- 生态学报. 2018, 38(13): 4724-4732
- Acta Ecologica Sinica. 2018, 38(13): 4724-4732
- http://dx.doi.org/10.5846/stxb201707091239
-
文章历史
- 收稿日期: 2017-07-09
- 网络出版日期: 2018-03-16





2. 内蒙古和盛生态科技研究院有限公司, 呼和浩特 011517
2. Inner Mongolia Hesheng Institute of Ecological Sciences & Technology, Hohhot 011517, China
蒙古莸(Caryopteris mongolica), 别名白蒿, 马鞭草科莸属旱生小灌木, 主要分布在蒙古高原的典型草原、荒漠化草原和荒漠区及黄土高原北部及西北部的典型草原区内。因其可药用、可提取芳香油以及可做饲料而具有较高的经济价值, 同时蒙古莸作为干旱区重要的水土保持和园林观赏植物而具有较高的生态价值[1-4]。近年来蒙古莸分布范围日渐缩小[5-7], 被《内蒙古重点保护草原野生植物名录》收录[8]。受其地理分布范围的影响, 国外对蒙古莸的研究较为少见, 国内学者对于蒙古莸的研究虽涉及领域较广, 但大多数研究主要集中在其分布范围[9]、抗旱性[5, 10-11]和生殖生物学特性[3, 5-6]等方面。这些研究展示了蒙古莸的基础生态学和生物学特性, 但却未能揭示出这一珍贵的资源植物日渐稀缺的原因。
从种子萌发到幼苗建成是高等种子植物生活史中的关键组成部分[12]。实生苗的生长和存活是影响种群空间分布和动态的决定性因素[13-14]。然而这一过程也是植物种群生活史中死亡率最高的时期之一[15-16]。植物在幼苗建植期间对外界环境条件的变化非常敏感[17], 幼苗从萌发出土后必须有较强的环境适应能力才能生存从而使种群得以延续[14]。然而, 近年来, 以高温、低温、干旱为主的极端气候事件越来越频繁[18]。不同的环境条件(如温度、水分)对植物种子萌发和幼苗生长造成显著的影响[19-20]。特别是在高温胁迫下植物产生的一系列生理变化直接影响其生长和繁殖[21-22]。目前, 对于蒙古莸种子萌发及幼苗建成的研究仍相对匮乏。郭春燕[23]曾对蒙古莸的种子萌发条件进行过探究, 其结果表明蒙古莸种子无明显休眠现象, 能够在自然条件下迅速萌发, 实验室内发芽率达到90%, 种子适宜的萌发条件为温度20—30℃, 土壤含水量7%—20%, 覆土厚度2—20 mm。谢乾瑾等[11]发现水分胁迫下的蒙古莸幼苗通过降低光合效率、蒸腾速率以减缓生长来适应干旱条件。宋鑫等[5]发现在不同供磷水平下蒙古莸的地上部分及根系形态会发生变化, 当供磷水平达到17.5 mg/kg时, 蒙古莸幼苗的生物量达到最大值。这些研究揭示出蒙古莸种子活力高、幼苗适应性强, 在自然条件下具有良好的繁殖扩散能力, 干旱、少磷等胁迫只能对其产生有限的影响。然而, 蒙古莸的分布范围正在不断缩小并成为稀缺的资源植物, 因此我们需要对蒙古莸种子萌发及幼苗生长过程中的各种限制因子进行更为系统全面的研究, 才能揭示出其分布范围缩小的原因。
本研究通过对内蒙古中西部干旱草原区蒙古莸天然种群实生苗的调查, 以及在室内模拟各种气候条件, 对蒙古莸种子萌发和幼苗建成进行研究, 旨在探究这一物种珍稀、濒危的原因。同时筛选出最优的种子萌发和幼苗生长条件, 凝练出通过以种子育苗为主的快速育苗技术方案, 为蒙古莸的人工繁育提供技术支撑。
1 研究方法 1.1 研究区概况研究区位于中国西北部的内蒙古和陕西省交界处神木县的大柳塔镇(110°19′53.58″E, 39°17′30.54″N, 海拔1218 m), 为黄土沟壑与沙地交错区。属中温带干旱、半干旱大陆性季风气候, 冬季寒冷而漫长, 夏季较为炎热而短暂。近20年的年均气温为(7.4±0.6)℃, 年均降水量为(346.5±90.0) mm。年均蒸发量为(2138.0±180.0) mm, 降水主要集中在7月到9月之间[24]。
选取的蒙古莸群落位于大柳塔镇东南部的硬梁地上, 地形起伏较大, 植被盖度约为40%, 蒙古莸为建群种, 主要伴生种有亚洲百里香(Thymus serpyllum var. asiatieus)、阿尔泰狗娃花(Heteropappus altaicus)、草木樨状黄芪(Astragalus melilotoides)等。
1.2 试验材料于2016年9月采集蒙古莸自然群落的种子带回实验室进行清选, 选出成熟饱满种子。同时在群落中取0—60 cm土层土壤带回实验室作为萌发用基质。
1.3 试验方法 1.3.1 实生苗调查在蒙古莸自然群落的不同坡面上随机设置9个4 m×4 m的样方。自2016年5月6日至2016年10月7日每隔15 d左右对样方内蒙古莸实生苗的数量进行调查, 对新发现的实生苗挂签记录, 跟踪其在整个生长季的存活情况。
1.3.2 种子萌发将试验地采集的土壤过1 mm孔径土壤筛, 之后在105℃烘箱中烘干48 h, 使土壤中没有可以萌发的植物种子, 并测定烘干后土壤的理化性质(表 1)。将烘干的土壤与蛭石按1:2比例混合后装入底部无孔的塑料桶中, 基质的饱和含水量为65.8%±2.2%。桶口径20 cm, 混合基质高约14 cm。
| pH | 碱解氮 Available N / (mg/kg) |
速效磷 Available P / (mg/kg) |
速效钾 Available K / (mg/kg) |
有机质 Organic matter / (g/kg) |
| 8.15 | 48.44 | 5.37 | 136.07 | 3.64 |
萌发条件采用3因素5水平正交试验设计(表 2), 每处理3个重复, 每重复50粒种子。将供试种子随机数出, 放入混合基质中, 在人工气候培养箱内进行萌发试验, 按照浇水周期进行补水。其中, 温度分为恒温条件和变温条件, 变温条件为每12 h温度进行一次变化。浇水周期指每两次浇水的时间间隔。每次浇水使用称重法使土壤水分含量为土壤饱和含水量的100%, 萌发实验持续21 d。试验期间每24 h统计新萌发种子数量, 以胚根突破种皮作为萌发标准, 埋入土中种子以子叶出土作为萌发标准。
| 试验号 Test number |
温度 Temperature/℃ |
浇水周期 Water cycle/d |
埋深 Burial depth/mm |
| 1 | 20 | 1 | 0 |
| 2 | 20 | 2 | 2 |
| 3 | 20 | 3 | 5 |
| 4 | 20 | 4 | 10 |
| 5 | 20 | 5 | 15 |
| 6 | 25 | 1 | 2 |
| 7 | 25 | 2 | 5 |
| 8 | 25 | 3 | 10 |
| 9 | 25 | 4 | 15 |
| 10 | 25 | 5 | 0 |
| 11 | 30 | 1 | 5 |
| 12 | 30 | 2 | 10 |
| 13 | 30 | 3 | 15 |
| 14 | 30 | 4 | 0 |
| 15 | 30 | 5 | 2 |
| 16 | 15/25 | 1 | 10 |
| 17 | 15/25 | 2 | 15 |
| 18 | 15/25 | 3 | 0 |
| 19 | 15/25 | 4 | 2 |
| 20 | 15/25 | 5 | 5 |
| 21 | 20/30 | 1 | 15 |
| 22 | 20/30 | 2 | 0 |
| 23 | 20/30 | 3 | 2 |
| 24 | 20/30 | 4 | 5 |
| 25 | 20/30 | 5 | 10 |
选取生长情况较为一致并长出第一对真叶的幼苗进行不同温度和水分条件下的幼苗建成试验。试验分温度-浇水周期组和温度-浇水量组两组进行。每处理3个重复, 每重复10株幼苗。试验设计如表 3所示。其中, 温度-浇水周期组的浇水量为土壤饱和含水量的100%。温度-浇水量组的浇水量按照土壤饱和含水量的百分比划分, 每隔1 d补水一次。除恒温条件外, 为模拟试验地土壤播种层日温度变化设置了3个高温胁迫梯度:
| 温度-浇水周期Group of temperature & water cycle | 温度-浇水量Group of temperature & water quantity | |||||
| 试验号 Test number |
温度/℃ Temperature |
浇水周期/d Water cycle |
试验号 Test number |
温度/℃ Temperature |
浇水量/% Watering quantity |
|
| 1 | 20 | 5 | 1 | 50(2 h) | 20 | |
| 2 | 20 | 10 | 2 | 50(2 h) | 40 | |
| 3 | 20 | 15 | 3 | 50(2 h) | 60 | |
| 4 | 20 | 20 | 4 | 50(2 h) | 80 | |
| 5 | 20 | 25 | 5 | 50(2 h) | 100 | |
| 6 | 25 | 5 | 6 | 50(4 h) | 20 | |
| 7 | 25 | 10 | 7 | 50(4 h) | 40 | |
| 8 | 25 | 15 | 8 | 50(4 h) | 60 | |
| 9 | 25 | 20 | 9 | 50(4 h) | 80 | |
| 10 | 25 | 25 | 10 | 50(4 h) | 100 | |
| 11 | 30 | 5 | 11 | 50(6 h) | 20 | |
| 12 | 30 | 10 | 12 | 50(6 h) | 40 | |
| 13 | 30 | 15 | 13 | 50(6 h) | 60 | |
| 14 | 30 | 20 | 14 | 50(6 h) | 80 | |
| 15 | 30 | 25 | 15 | 50(6 h) | 100 | |
| 16 | 50(2 h) | 5 | ||||
| 17 | 50(2 h) | 10 | ||||
| 18 | 50(2 h) | 15 | ||||
| 19 | 50(2 h) | 20 | ||||
| 20 | 50(2 h) | 25 | ||||
| 括号内为高温时间长度 | ||||||
20℃(8 h)/25℃(7 h)/50℃(2 h)/25℃(7 h)(下简称50℃(2 h));
20℃(8 h)/25℃(6 h)/50℃(4 h)/25℃(6 h)(下简称50℃(4 h));
20℃(8 h)/25℃(5 h)/50℃(6 h)/25℃(5 h)(下简称50℃(6 h))。
试验期间每隔5 d统计幼苗存活数量, 每隔24 d统计幼苗的株高、基径和叶龄。株高为自然高度, 叶龄为真叶对数。
1.4 数据处理
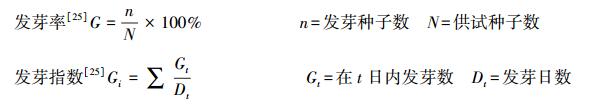
|
幼苗存活率[25]=存活幼苗数/初始幼苗数×100%
用Excel 2013软件对所得数据进行统计, 用SAS软件进行单因素和正交方差分析, 用SigmaPlot 12.2软件进行双因素方差分析并作图。
2 结果分析 2.1 自然群落实生苗更新蒙古莸自然群落内实生苗的存活情况如图 1所示。实生苗数量随着时间变化呈现出先增加后减少的趋势, 新萌发的实生苗从5月初起持续被发现, 在7月末实生苗数量达到最高值54株, 到9月中旬剩余16株后不再变化。在8月下旬实生苗死亡数量达到峰值。至10月中旬生长季结束, 累计出现81株实生苗, 死亡65株。实生苗的死亡率高达80.20%。实生苗的更新强度为0.11株/m2。

|
| 图 1 蒙古莸群落实生苗存活、死亡数量 Fig. 1 The survival and mortality number of seedlings |
蒙古莸种子在适宜条件下播种15 d内完成萌发。在覆土情况下萌发率最高可达90.7%。播种20 d左右开始长出第一对真叶, 进入幼苗期。如表 4所示, 萌发温度和覆土厚度对种子的萌发率和发芽指数都存在极显著影响。温度是影响种子萌发的重要因子(R=57.24)。种子适宜在恒温20—25℃之间及变温20℃/30℃的条件下萌发, 覆土的最适宜厚度为10—15 mm。浇水周期对种子的萌发率没有显著影响, 但对发芽指数存在显著影响。在任何浇水周期下, 种子都有着较高的萌发率, 在每隔两天浇水一次时种子的发芽指数达到最高值, 能够更快速的萌发。
| 水平Level | 萌发率Germination rate/% | 发芽指数Germination index | |||||
| 温度 Temperature |
浇水间隔 Water cycle |
埋深 Burial depth |
温度 Temperature |
浇水间隔 Water cycle |
埋深 Burial depth |
||
| 水平1 Level 1 | 69.73a | 48.67b | 31.47c | 7.52a | 4.57ab | 1.86c | |
| 水平2 Level 2 | 72.68a | 60.13a | 49.87b | 6.92a | 5.81a | 4.99b | |
| 水平3 Level 3 | 49.47b | 58.53ab | 54.13b | 4.54b | 5.70ab | 5.70ab | |
| 水平4 Level 4 | 15.47c | 50.13ab | 67.60a | 0.61c | 4.46b | 6.48a | |
| 水平5 Level 5 | 62.40a | 52.27ab | 66.67a | 5.43b | 4.48b | 6.00ab | |
| 极差Range | 57.24** | 11.46 | 36.14** | 6.90** | 1.35* | 4.62** | |
| 不同小写字母表示方差分析结果存在显著差异(P<0.05)。*表示方差分析结果存在显著差异(P<0.05), **表示方差分析结果存在极显著差异(P<0.01) | |||||||
温度-浇水周期对蒙古莸幼苗的生长存活的影响如表 5所示。根据双因素方差分析结果, 温度显著影响幼苗的存活率。在恒温25℃及每日2 h 50℃高温胁迫条件下幼苗存活率达53.33%以上, 持续的高温和低温对幼苗的存活造成非常显著的伤害, 在恒温20℃时幼苗的存活率仅为2.00%。不同的浇水周期对幼苗的存活存在显著影响, 最佳浇水周期为每隔20 d浇水一次, 幼苗存活率可达55.00%。减少浇水周期至每隔5 d浇水一次显著影响了幼苗的存活, 使其存活率降为3.33%。
| 温度 Temperature |
浇水周期Water cycle | |||||
| 5 d | 10 d | 15 d | 20 d | 25 d | 均值Mean value | |
| 20℃ | 0 | 0 | 6.67 | 3.33 | 0 | 2.07c |
| 25℃ | 6.67 | 53.33 | 76.67 | 66.67 | 63.33 | 53.33a |
| 30℃ | 0 | 20.00 | 20.00 | 63.33 | 33.33 | 27.33b |
| 50℃(2 h) | 6.67 | 53.33 | 63.33 | 86.67 | 70.00 | 56.00a |
| 均值Mean value | 3.33b | 31.67a | 41.67a | 55.10a | 41.67a | |
| 不同小写字母表示方差分析结果存在显著差异(P<0.05) | ||||||
温度-浇水量对蒙古莸幼苗的生长存活的影响如图 2所示。根据双因素方差分析结果, 随着高温胁迫时间的延长, 幼苗存活率显著降低(P<0.05)。高温胁迫2 h后幼苗存活率为25.33%, 当胁迫时间延长至6 h后幼苗存活率仅为6.67%。浇水量对幼苗存活存在显著影响, 浇水量达到田间持水量的100.0%时幼苗存活率显著高于其他处理, 可达33.33%。浇水量为田间持水量的20.0%时, 幼苗存活率低至4.44%。说明在高温胁迫条件下幼苗生长的最适宜土壤水分条件为田间持水量的100.0%。

|
| 图 2 温度-浇水量组蒙古莸幼苗存活率 Fig. 2 The seedling survival rate of temperature & water quantity group |
蒙古莸幼苗在不同温度下存活率动态变化如图 3所示, 在恒温25℃及每日2 h 50℃高温胁迫条件下, 幼苗的存活率一直处于较高的水平, 在恒温30℃条件下, 幼苗存活率在生长期内保持稳定的下降, 没有出现明显的斜率变化。在恒温20℃条件下, 幼苗在生长初期存活率急剧下降, 并于55 d后下降到2.00%。说明幼苗对持续高温和持续低温的耐受能力较差, 特别是在持续低温环境中很快死亡。

|
| 图 3 不同温度条件下蒙古莸幼苗存活率动态变化 Fig. 3 The dynamic change of seedling survival rate for different temperature conditions |
蒙古莸幼苗在不同浇水周期下存活率动态变化如图 4所示。在10、15、20、25 d浇水周期的情况下, 幼苗前55 d存活率变化基本一致, 呈现出稳步减少的趋势。后期每隔20 d浇水一次的幼苗表现出较高的存活率。每隔5 d浇水一次的幼苗存活率较其他处理明显偏低。在幼苗生长50 d后, 曲线下降斜率明显增大, 说明从这一时期开始, 幼苗死亡率增大。120 d试验结束时幼苗存活率仅为3.33%。

|
| 图 4 不同浇水周期下蒙古莸幼苗存活率动态变化 Fig. 4 The dynamic change of seedling survival rate for different water cycle conditions |
由表 6可以看出, 温度对蒙古莸幼苗的株高、基径不存在显著影响。在恒温20℃时, 幼苗叶龄为最小值, 在恒温25℃时叶龄达到最大值, 说明恒温25℃条件是蒙古莸叶片生长的最适宜温度。在变温条件下株高与枝长比显著大于其他处理。表明变温条件有利于植株茎的直立生长和增粗。缩短浇水周期至每隔5 d一次使蒙古莸株高和基径显著增大, 而对于幼苗的叶龄没有显著影响。随着浇水周期的延长, 幼苗的基径呈现出递减的变化趋势。表明每隔5 d浇水一次有利于茎的增粗, 延长浇水间隔影响幼苗生长, 使植株趋于矮小, 但叶龄不发生变化。
| 处理 Treatment |
水平 Level |
株高 Plant height/cm |
基径 Basal stem diameter/mm |
叶龄 Leaf age |
自然高度与绝对高度比值 Ratio of natural height to absolute height |
| 温度 Temperature |
20℃ | 1.47±0.96a | 0.48±0.08a | 5.41±0.66c | 0.21±0.18b |
| 25℃ | 3.92±0.38a | 0.61±0.03a | 9.85±0.26a | 0.27±0.07b | |
| 30℃ | 3.07±0.47a | 0.59±0.04a | 9.57±0.32ab | 0.43±0.09b | |
| 50℃(2 h) | 4.22±0.38a | 0.70±0.03a | 8.91±0.26b | 0.77±0.07a | |
| 浇水周期 Water cycle |
5d | 5.72±0.96a | 0.80±0.08a | 8.30±0.66a | 0.55±0.18a |
| 10d | 2.49±0.51ab | 0.58±0.04ab | 8.70±0.36a | 0.40±0.10a | |
| 15d | 2.97±0.45ab | 0.58±0.04ab | 8.94±0.31a | 0.38±0.09a | |
| 20d | 2.68±0.43ab | 0.52±0.04b | 8.11±0.30a | 0.31±0.08a | |
| 25d | 2.00±0.52b | 0.49±0.04b | 8.13±0.36a | 0.47±0.10a | |
| 不同小写字母表示方差分析结果存在显著差异(P<0.05) | |||||
蒙古莸种子不存在休眠现象, 当年生产的种子在适宜条件下即可萌发。温度对种子的萌发影响甚大。恒温条件下, 种子的适宜萌发温度为20—25℃之间, 30℃的恒温即对种子萌发产生不利影响。这与郭春燕等的研究结果基本一致[23]。变温20℃/30℃条件下, 种子的萌发与恒温20—25℃相若。但在15℃/25℃变温下, 种子的萌发率和发芽指数显著小于其他处理。每日15℃左右的低温持续12 h会使的种子萌发率显著下降。30℃的高温对种子的萌发也有一定的抑制作用。这说明蒙古莸种子萌发的适宜温度范围较窄。
蒙古莸自然群落中实生苗的成活率较低, 仅为19.8%。从幼苗建成对温度的反应推测, 持续的高温和低温会影响幼苗的存活。为探究蒙古莸幼苗对高温的耐受能力, 我们在实验室模拟了这一时期内每日地表温度的变化。结果发现短时间(2 h)的高温(50℃)胁迫下幼苗的存活率较高, 且相比其他恒温条件, 幼苗的株高更高, 基径更粗, 株高与枝长比的值最大。而长时间的高温胁迫(4 h以上)使幼苗死亡率显著增大。说明蒙古莸比较适应变温的环境, 对短时间高温的耐受能力较强。
大柳塔地区5月到9月土壤0—3 cm深度温度变化如图 5所示。蒙古莸在最低温明显低于15℃的4—6月少有萌发。在西北干旱地区, 春季常有大风和倒春寒的发生, 降低在此时的萌发率能够提高萌发后幼苗存活率。之后的温度条件越来越适合种子的萌发, 在温度最高的7月蒙古莸达到萌发高峰, 至此土壤种子库中能够萌发的种子大量萌发。蒙古莸幼苗在7月中旬到8月下旬之间死亡数量明显增多的原因可能与此时段的温度过高有关。图 5显示, 从6月中旬开始, 土壤中种子萌发层(0—3 cm)的温度即可达到50℃。7月中旬是西北黄土丘陵地区年内气温最高时期[26]。地温超过40℃(甚至50℃)的日数很多。蒙古莸的生长环境中, 地表裸露, 在夏季太阳直射时地表温度迅速升高。而蒙古莸幼苗只能够适应短期的高温胁迫, 随着每日高温胁迫时间的延长, 幼苗死亡率逐渐增高。这样的高温环境可持续至8月中旬。8月下旬后, 温度逐渐降低, 但土壤种子库中在此时已耗尽了原有的种子储备, 从而表现新生幼苗明显减少的现象。

|
| 图 5 研究区5—9月土壤表面温度 Fig. 5 Soil surface temperature in study site from May to September |
我国干旱半干旱地区限制植物存活生长的主要因素就是水分[19]。郭春燕等[23]的研究表明蒙古莸种子对水分条件非常敏感。在温度条件适宜, 且水分充足时迅速完成吸胀过程, 时间约为1.5 h。水分充足条件下, 不同的浇水周期对种子的萌发率没有显著的影响, 但能够提高发芽指数, 即使更多的种子快速萌发。研究区的土壤水分主要来自于降雨, 降雨多集中在7—9月, 占年降水量的69%, 并大多以暴雨的形式出现[24, 27]。这一时期的地表蒸发强烈, 水分在浅层土壤中存留时间较短[23]。蒙古莸能够适应研究区特殊的水分条件, 种子迅速吸胀, 并在3—5 d萌发出土, 响应夏季有限的降雨, 从而使幼苗尽早的生长。
不同的水分条件对幼苗的存活有显著影响[19], 室内试验结果显示频繁的浇水会增大蒙古莸幼苗的死亡率, 而与此同时在高温胁迫条件下将土壤水分维持在田间持水量的最大值有助于幼苗的生长存活。因此, 蒙古莸幼苗的生长对土壤的保水能力要求较高。自然群落实生苗调查结果显示, 在降雨集中的7、8月份是幼苗死亡的高峰期。这一时期的降雨量虽然多, 但由于降雨强度过大加之黄土丘陵地区疏松的土质使得水分快速的流失或入渗[23, 28]。这一水量能够供给蒙古莸种子萌发却不能够长时间储存在浅层土壤中保证幼苗进一步生长。同时在夏季正午近乎50℃的地温加之较为频繁的降水使得地表蒸发作用强烈, 形成了类似“蒸笼”的地表环境, 使得幼苗难以存活。萌发出土后的幼苗无法适应外界环境, 容易出现较高的死亡率。
3.3 埋深对种子萌发的影响埋藏深度影响种子在土壤中贮藏的生物和非生物条件, 最终影响种子的萌发和幼苗的生长[29], 干旱区种子的沙埋深度在0—20 mm[30], 浅层埋藏为种子提供了较为适宜的温度和水分[31]。蒙古莸种子在埋深10—15 mm时萌发率最高。此处利于种子充分吸收土壤水分并顺利出苗, 且种子萌发后根系能够较容易地固定并吸收水分, 从而增加了幼苗的存活率。造成埋深为0的种子不能萌发的主要原因是种子裸露在空气中, 难以有效吸收和保存水分, 影响种子正常萌发。
4 结论虽然蒙古莸种子不存在休眠现象, 萌发对水分的要求较宽范, 但适宜其种子萌发的温度范围较窄。而且, 适宜于种子萌发的条件与适宜于幼苗存活的条件有较大差异, 致使其从种子萌发到完成幼苗建成全过程的环境“窗口”进一步缩小。其生境地的自然条件往往难以满足从种子萌发到幼苗建成全过程的要求。这是影响野生蒙古莸种群通过有性繁殖, 扩大种群规模的主要原因之一。
根据从种子萌发到幼苗建成各时期的存活特征, 可将蒙古莸从种子萌发到幼苗建成历程分为3个时期, 第一个是种子萌发期, 第二个是幼苗死亡多发期, 第三个是稳定生长期。在蒙古莸自然群落生境中萌发期从5月持续到7月初, 这一时期大量的种子萌发出土。然而随即到来的高温、多雨环境将幼苗直接带入死亡多发期, 这一时期的幼苗大量死亡。直至9月, 侥幸存活的幼苗进入稳定生长期。存活的幼苗作为种群自然更新的主要成分为种群的扩散做出贡献。
| [1] | 王晓江, 李爱平, 宁明世, 张纪刚. 生态灌木蒙古莸的生物生态学特性及其经济价值评价. 干旱区资源与环境, 2006, 20(2): 191–194. |
| [2] | 李玉俊, 李新荣, 杨喜林. 芳香植物——蒙古莸的生态生理特性及其栽培技术. 中国沙漠, 1991, 11(3): 50–56. |
| [3] | 高建平, 贺晓, 郭春燕, 石小兰. 蒙古莸小孢子发生和雄配子体发育的研究. 西北植物学报, 2009, 29(6): 1149–1154. |
| [4] | Mole E. Recent progress with the Caryopteris collection. Plant Heritage, 2004, 11(2): 14–15. |
| [5] | 宋鑫, 张东梅, 张丽静, 胡晓炜, 陈晓龙, 张燕慧. 施磷对蒙古莸(Caryopteris mongholica)形态和酸性磷酸酶活性的影响. 中国沙漠, 2015, 35(4): 917–922. DOI:10.7522/j.issn.1000-694X.2014.00089 |
| [6] | 郭春燕, 贺晓, 高建平, 刘果厚. 蒙古莸的开花物候与生殖特征. 西北植物学报, 2012, 32(10): 2040–2046. DOI:10.3969/j.issn.1000-4025.2012.10.017 |
| [7] | 高建平. 蒙古莸的胚胎学研究[D]. 呼和浩特: 内蒙古农业大学, 2010. |
| [8] | 武向良. 内蒙古重点保护草原野生植物名录. 呼和浩特: 内蒙古农牧业厅, 2009. |
| [9] | 赵一之. 蒙古莸的植物区系地理分布研究. 内蒙古大学学报:自然科学版, 1995, 26(2): 195–197. |
| [10] | 韩磊, 贺康宁, 芦新建, 王占林. 青海高寒半干旱区蒙古莸叶水势变化及其与环境因素的关系. 水土保持通报, 2008, 28(6): 1–5. |
| [11] | 谢乾瑾, 夏新莉, 刘超, 尹伟伦. 水分胁迫对不同种源蒙古莸光合特性与生长的影响. 林业科学研究, 2010, 23(4): 567–573. |
| [12] | Wilbur H M. Life history evolution in seven milkweeds of the genus Asclepias. Journal of Ecology, 1976, 64(1): 223–240. DOI:10.2307/2258693 |
| [13] | Baker P J. Seedling establishment and growth across forest types in an evergreen/deciduous forest mosaic in western Thailand. Natural History Bulletin of the Siam Society, 1997, 45(1): 17–41. |
| [14] | 苏嫄, 焦菊英, 王志杰. 陕北黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征. 植物生态学报, 2014, 38(7): 694–709. |
| [15] | 武高林, 杜国祯. 植物种子大小与幼苗生长策略研究进展. 应用生态学报, 2008, 19(1): 191–197. |
| [16] | 李雪华, 蒋德明, 刘志民, 李晓兰. 温带半干旱地区一年生植物种子的萌发特性. 生态学报, 2006, 26(4): 1194–1199. |
| [17] | Jalili A, Hamzeh'ee B, Asri Y, Shirvany A, Yazdani S, Khoshnevis M, Zarrinkamar F, Ghahramani M A, Safavi R, Shaw S, Hodgson J G, Thompson K, Akbarzadeh M, Pakparvar M. Soil seed banks in the Arasbaran Protected Area of Iran and their significance for conservation management. Biological Conservation, 2003, 109(3): 425–431. DOI:10.1016/S0006-3207(02)00170-2 |
| [18] | 张凯, 慕小倩, 孙晓玉, 汪梦竹, 胡胜武. 温度变化对油菜及其伴生杂草种苗生长和幼苗生理特性的影响. 植物生态学报, 2013, 37(12): 1132–1141. |
| [19] | 马赟花, 张铜会, 刘新平, 毛伟, 岳祥飞. 春季小降雨事件对科尔沁沙地尖头叶藜萌发的影响. 生态学报, 2015, 35(12): 4063–4070. |
| [20] | 李文婷, 张超, 王飞, 郑明清, 郑元润, 张峰. 沙埋与供水对毛乌素沙地两种重要沙生植物幼苗生长的影响. 生态学报, 2010, 30(5): 1192–1199. |
| [21] | 李胜利, 夏亚真, 刘金, 师晓丹, 孙治强. 高温胁迫下番茄幼苗对冷激的响应. 应用生态学报, 2014, 25(10): 2927–2934. |
| [22] | Lobell D B, Schlenker W, Costa-Roberts J. Climate trends and global crop production since 1980. Science, 2011, 333(6042): 616–620. DOI:10.1126/science.1204531 |
| [23] | 郭春燕. 蒙古莸生殖生物学研究[D]. 呼和浩特: 内蒙古农业大学, 2009. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y1474298 |
| [24] | 贺一鸣, 贺晓, 包丽颖, 郭春燕, 高建平, 李琪. 蒙古莸(Caryopteris mongolica)种群空间格局及枝系构型. 中国沙漠, 2016, 36(4): 1007–1013. DOI:10.7522/j.issn.1000-694X.2015.00133 |
| [25] | 田璐, 张沛东, 牛淑娜, 张秀梅, 张凌宇. 不同处理对大叶藻种子萌发和幼苗建成的影响. 生态学杂志, 2014, 33(9): 2408–2413. |
| [26] | 王若安. 平凉黄土高原沟壑区最高最低气温对气候变暖的响应. 干旱区资源与环境, 2010, 24(6): 72–76. |
| [27] | 杨毅. 神木县矿山地质环境影响评价研究[D]. 西安: 西安科技大学, 2008. http://jz.docin.com/p-148745776.html |
| [28] | 尹秋龙, 焦菊英, 寇萌. 极端强降雨条件下黄土丘陵沟壑区不同植被类型土壤水分特征. 自然资源学报, 2015, 30(3): 459–469. DOI:10.11849/zrzyxb.2015.03.009 |
| [29] | 苏延桂, 李新荣, 贾荣亮, 冯丽. 沙埋对六种沙生植物种子萌发和幼苗生长的影响. 中国沙漠, 2007, 27(6): 968–971. |
| [30] | 王国华, 赵文智. 埋藏深度对梭梭(Haloxylon ammodendron)种子萌发及幼苗生长的影响. 中国沙漠, 2015, 35(2): 338–344. DOI:10.7522/j.issn.1000-694X.2014.00013 |
| [31] | 王国华, 赵文智. 不同沙埋深度对当年生柠条锦鸡儿种子萌发及幼苗生长的影响. 干旱区地理, 2016, 39(1): 95–103. |