 2018, Vol. 38
2018, Vol. 38文章信息
- 牛钰杰, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民.
- NIU Yujie, YANG Siwei, WANG Guizhen, LIU Li, DU Guozhen, HUA Limin.
- 放牧干扰下高寒草甸物种、生活型和功能群多样性与生物量的关系
- Relationship between plant species, life form, functional group diversity, and biomass under grazing disturbance for four years on an alpine meadow
- 生态学报. 2018, 38(13): 4733-4743
- Acta Ecologica Sinica. 2018, 38(13): 4733-4743
- http://dx.doi.org/10.5846/stxb201707061223
-
文章历史
- 收稿日期: 2017-07-06
- 网络出版日期: 2018-03-16





2. 毕节市畜牧兽医科学研究所, 毕节 551700;
3. 兰州大学生命科学学院, 兰州 730030
2. Institute of Animal and Veterinary Science, Bijie 551700, China;
3. School of Life Sciences, Lanzhou University, Lanzhou 730030, China
青藏高原东缘约有1900种植物分布[1], 且每平米物种丰富度最高可达50种[2]。高寒草甸作为青藏高原主体生态系统类型, 是畜牧生产的主要资源载体, 也是植物多样性保存相对完好的地域之一[3-4]。多样性与生产力之间的关系是高寒草甸最主要的研究热点之一[5-9]。Tilman等[10]创造性地提出功能多样性比物种多样性更能决定生态系统生产力, 进而将作为物种组合类群多少的功能多样性引入到多样性与生产力关系的研究之中。但Thompson等[11]和马文静等[8]部分学者也发现功能多样性与生产力无关。物种多样性和功能多样性究竟哪个更能决定生产力的大小, 一直争议很大[6-9]。
这种争议一方面是由于不同研究者选择的多样性指数不同[12]。现今多样性指数的种类多种多样[12]。按组织层次不同可以将多样性分为物种多样性和功能多样性, 而物种和功能多样性按其强调多样性组分不同, 又均可以分为丰富度[13]、优势度[14]和异质性测度[15]的统计指标。王长庭等[16]选择丰富度指数、Simpson指数和均匀度指数研究物种多样性与生物量之间的关系, 发现不同多样性指数与生物量之间相关系数差别很大。单就功能多样性而言, 由于植物功能特征不是绝对单一的, 所以对植物功能的界定就会有不同的理解和划分方法, 导致人为主观性较强[6, 17]。根据研究目的不同, 研究者选用不同的功能分类方法。而以生活型态[18]和经典的林奈生物学分类法[6, 8]为原始功能分类依据较为常见。王国杰等[19]使用不同的功能多样性指标对内蒙古草原群落进行研究发现, 水分生态类型多样性比物种和生活型多样性更能反映与生产力之间的关系, 从而得出多样性与生产力的关系受多样性指数选择的影响。因此, 在生态学研究中, 多样性与生产力关系因研究者选择不同功能多样性和物种多样性指数而异。
另一方面, 研究者在野外植物调查中普遍使用地上现存量直接表示生产力, 来研究其与多样性之间的关系[4, 8-10, 19]。但除了在受控试验条件下可以用草地生物量准确折算出草地的初级生产力外[20], 草地植物现存量极大地受放牧干扰的影响[19]。放牧作为所有草地类型最主要且不可避免的干扰方式[3], 其对植物群落多样性和生产力的影响取决于干扰的强度特征和时间累积效应[21]。而不同放牧强度对植物群落的影响是在空间尺度上进行比较, 具有瞬时性[3]。但因为草地植物本身具有补偿性生长和每年重新萌发的特性, 以放牧年份为代表的时间累积效应对植物多样性和生物量的影响就很模糊[21-22]。导致放牧干扰下多样性与生产力之间的关系更加不确定, 鉴于放牧干扰不可避免且普遍存在, 那么研究不同放牧强度长时间持续干扰下多样性与生物量之间的关系, 对多样性与生产力之间的相互预测就具有更多的科学和现实意义。
基于上述分析, 本研究在青藏高原东北缘高寒草甸选择6块不同放牧强度样地, 连续4 a进行物种、生活型和功能群多样性调查与生物量监测, 研究不同放牧强度作用下多样性与生物量之间的关系。以期明晰不同层次的多样性对放牧干扰在空间强度特征与时间尺度累积作用下的敏感性以及多样性和生物量之间的相互关系对放牧干扰的响应, 最终为有效评估高寒草甸在空间和时间尺度上的植物多样性变化和建立合理的放牧制度提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于青藏高原东北缘, 植被类型为高寒嵩草草甸。行政上隶属甘肃省天祝县抓喜秀龙乡(37°11′—37°14′ N, 102°40′—102°47′ E), 海拔2958 m。年均气温-0.1℃, >0℃的年积温为1380℃。年均降水量416 mm, 主要集中在7—9月, 占全年降雨量的76%。无绝对无霜期, 植物生长期120—140 d。土壤类型为高山草甸土, 土层厚度40—80 cm。优势物种为垂穗披碱草(Elymus nutans)、冷地早熟禾(Poa crymophila)、西北针茅(Stipa krylovii)、矮生嵩草(Kobresia humilis)和阴山扁蓿豆(Melissitus ruthenica)等。
1.2 试验设计在系统考察试验地植被情况和调查当地实际载畜量的基础上, 选择地势平坦、草地类型一致、优势种为垂穗披碱草和大针茅的总面积共10 hm2样地, 此样地在围栏放牧试验之前以3.69羊单位/hm2进行轻度的自由放牧管理。根据当地的草地生物量计算草地理论承载的羊单位头数为4.5—5.0羊单位/hm2, 进而设置6个放牧强度区(放牧强度=羊单位数×放牧时间/放牧面积)开展控制放牧试验(表 1), 选择体重相近, 约为45 kg的甘肃高山细毛羊为放牧家畜。以全年每天连续放牧的方式进行放牧管理。放牧作用于基础条件一致的草地, 消除了地形和气候因素的干扰。以当地平均放牧强度作为基准, 上下浮动设置放牧强度梯度, 并定义高于当地平均放牧强度(中等放牧强度)的样地4、样地5和样地6为高放牧强度样地, 低于当地平均放牧强度的样地1、样地2和样地3为低放牧强度样地。于2012年8月, 开始围栏放牧, 并连续4 a对样地植物群落进行定点调查。
| 样地 Plots |
面积/hm2 Areas |
物种丰富度 Species richness |
盖度/% Cover |
家畜头数/只 Livestock number |
放牧强度/(羊单位/hm2) Grazing intensity |
| 1 | 2.18 | 8.00±1.63b | 88.50±3.42a | 6 | 2.75 |
| 2 | 1.10 | 8.25±2.22b | 91.00±6.22a | 4 | 3.63 |
| 3 | 1.61 | 13.40±2.07a | 89.20±6.30a | 7 | 4.36 |
| 4 | 1.26 | 10.00±2.23b | 89.20±4.44a | 6 | 4.78 |
| 5 | 2.50 | 10.40±1.67b | 91.20±1.48a | 13 | 5.19 |
| 6 | 1.11 | 11.00±1.00b | 90.57±3.67a | 8 | 7.20 |
| 不同小写字母表示不同放牧强度样地各项指标差异显著(P < 0.05) | |||||
2012—2015年, 每年8月15号左右进行样地植物群落调查。采用系统随机取样法, 将每个样地规则地分为9个小区。在每个小区内随机地设置一个样方, 取样时避开样地群落边缘, 即每个样地重复取样9次。4年共调查样方216个。在2012年首次取样后将取样点固定。之后每年在固定取样点调查取样。
1.3.2 调查和计算方法群落内每一物种盖度:用0.25 m2样方, 针刺100次;物种高度:每一种植物随机测量10次;物种频度:每样方附近随机抛样圆20次, 记录每一物种频度。用最大值标准化法计算每一物种高度、盖度和频度的相对值。物种重要值计算公式为:IV=(A+B+C)/3。式中:A为相对盖度;B为相对频度;C为相对高度。用各生活型和功能群植物的重要值之和表示生活型和功能群重要值。根据植物生活形态和功能性划分方法分别将物种划分为6个生活型(丛生型、根茎型、莲座型、匍匐型、一年生、直立型)[23]和4个功能群(禾本科、莎草科、豆科、杂类草)[6, 8, 24]。在选择生活型时, 因为根茎型植物的耐牧性和一年生植物对放牧作用的敏感性, 因而将其优先从植物中筛选出来, 其余4种生活型根据茎的形态(丛生、莲座、匍匐、直立)也可进行完全划分。功能群的选择按照经典的林奈生物学分类法进行。不同组织层次(物种、生活型和功能群)的多样性指数均选取丰富度、均匀度和优势度作为测度指标[12, 25]。物种、生活型和功能群丰富度均用0.25 m2样方, 分别以其中出现的物种数、生活型种类和功能群种类表示。再用重要值计算样方群落的物种、生活型和功能群均匀度(Evenness指数)[15]和优势度(Berger-Parker指数)[14]。计算公式为:
Evenness指数:
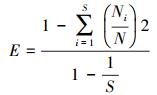
|
Berger-Parker指数:
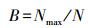
|
式中, S为每个样方总的物种数、生活型种类和功能群种类;N为S个全部物种、生活型和功能群的相对重要值之和;Ni为第i个物种、生活型和功能群的相对重要值, Nmax为相对重要值最大值的物种、生活型和功能群。
1.4 数据分析对放牧强度作用下物种、生活型和功能群多样性(丰富度、均匀度和优势度)做随放牧年份累积作用下的重复测量方差分析(LSD法进行显著性检验), 并对变量进行球形检验, 用Greenhouse-Geisser法对自由度进行校正。用Pearson相关性分析法对各多样性指数的相互关系进行分析。采用线性回归法分析物种、生活型和功能群多样性(丰富度、均匀度、优势度)与生物量之间的关系, 设定P值为0.05, 用决定系数R2, 解释生物量对各多样性的贡献大小。所有统计分析均使用SPSS 19.0软件进行。
2 结果与分析 2.1 地上生物量在放牧干扰下的变化生物量随放牧强度和放牧年份均发生显著变化。2012年的生物量在不同放牧强度区无显著变化。而2013—2015年, 生物量随放牧强度的增加均表现出显著降低的变化趋势。而不同放牧强度区, 生物量随放牧年份的整体变化规律不一致, 但2013—2015年不同放牧强度区生物量均低于围栏放牧前2012年(表 2)。
| 样地 Plots |
地上生物量Aboveground biomass /(g/m2) | |||
| 2012 | 2013 | 2014 | 2015 | |
| 1 | 310.40±76.97a(a) | 290.58±65.85a(ab) | 312.66±53.28a(a) | 241.89±64.31a(b) |
| 2 | 321.00±17.40a(a) | 321.20±80.36a(a) | 315.39±62.99a(a) | 210.31±31.39ab(b) |
| 3 | 293.60±31.70a(a) | 126.80±29.84b(d) | 232.60±36.29b(b) | 168.08±18.41bc(c) |
| 4 | 260.00±44.18a(a) | 111.87±25.22b(c) | 210.01±7.87b(b) | 126.34±37.16cd(c) |
| 5 | 310.40±76.97a(a) | 101.60±26.58bc(c) | 203.64±44.34b(b) | 121.53±46.48cd(c) |
| 6 | 298.50±55.71a(a) | 53.60±19.65c(b) | 141.75±10.13c(b) | 89.43±20.20d(b) |
| 括号内为横向比较, 括号外为纵向比较。显著水平:P < 0.05 | ||||
对于丰富度, 放牧年份对物种、生活型和功能群丰富度的影响均达到显著水平。而不同放牧强度之间丰富度无显著差异(P>0.05)。放牧年份和放牧强度对物种丰富度的变化具有交互作用, 放牧年份对物种丰富度的作用随放牧强度的不同而异, 表现为低强度放牧样地(Plot1、Plot2、Plot3)的物种丰富度高于高强度放牧样地(Plot4、Plot5、Plot6)(图 1)。对于均匀度, 放牧年份对物种、生活型和功能群均匀度的影响均达到显著水平。放牧年份和放牧强度对物种、生活型和功能群均匀度均无交互作用, 放牧年份对均匀度的作用随放牧强度无显著变化(图 1)。对于优势度, 放牧年份对物种和功能群优势度的影响达到显著水平, 而对生活型优势度无影响。放牧年份和放牧强度对物种、生活型和功能群优势度均无交互作用, 放牧年份对优势度的作用随放牧强度无显著变化(图 1)。

|
| 图 1 不同放牧强度下丰富度、均匀度和优势度随放牧年份的变化 Fig. 1 Variation of richness, evenness, Berger-parker under the grazing intensity with different grazing year |
在连续放牧干扰下, 所有多样性指数中, 仅物种丰富度与其余多样性指数均表现显著的相关关系, 其与物种均匀度、物种优势度、生活型优势度和功能群均匀度显著负相关, 与生活型丰富度、生活型均匀度、功能群丰富度和功能群优势度显著正相关。生活型均匀度与生活型优势度的相关性最高, 其相关系数为-0.818。其次为功能群均匀度和功能群优势度, 相关系数为-0.786(表 3)。
| 植物层次 Plant level |
多样性 Diversity |
物种Species | 生活型Life form | 功能群Functional group | ||||||||
| 丰富度 Richness |
均匀度 Evenness |
优势度 Berger-parker |
丰富度 Richness |
均匀度 Evenness |
优势度 Berger-parker |
丰富度 Richness |
均匀度 Evenness |
优势度 Berger-parker |
||||
| 物种 | 丰富度 | 1 | ||||||||||
| Species | 均匀度 | -0.452** | 1 | |||||||||
| 优势度 | -0.435** | -0.343** | 1 | |||||||||
| 生活型 | 丰富度 | 0.535** | -0.435** | -0.153 | 1 | |||||||
| Life form | 均匀度 | 0.190* | -0.056 | -0.094 | 0.199* | 1 | ||||||
| 优势度 | -0.358** | 0.172* | 0.112 | -0.271** | -0.818** | 1 | ||||||
| 功能群 | 丰富度 | 0.449** | -0.312** | -0.290** | 0.352** | 0.160* | -0.220** | 1 | ||||
| Functional | 均匀度 | -0.252** | 0.007 | 0.158* | 0.057 | -0.199* | 0.143 | 0.153* | 1 | |||
| group | 优势度 | 0.383** | -0.135 | -0.146 | 0.072 | 0.427** | -0.425** | -0.007 | -0.786** | 1 | ||
| *P < 0.05; **P < 0.01 | ||||||||||||
低强度放牧样地的Plot2中物种丰富度随生物量增加而显著降低(R2=0.140)。其余放牧强度样地的物种、生活型和功能群丰富度均与生物量无关(图 2)。

|
| 图 2 不同放牧强度区丰富度与生物量的关系 Fig. 2 Relationship between richness and biomass in different plots |
低强度放牧样地的Plot1中, 生活型均匀度随生物量的增加而显著降低。而低强度放牧样地的Plot2中, 物种均匀度随生物量的增加而显著升高。高强度放牧样地的Plot5中, 功能群均匀度随生物量的增加而显著降低。在高强度放牧样地(Plot4、Plot5、Plot6)中, 生活型均匀度均随生物量的增加而显著降低。其余多样性指数与生物量无关(图 3)。

|
| 图 3 不同放牧强度区均匀度与生物量的关系 Fig. 3 Relationship between evenness and biomass in different plots |
在低强度放牧样地的Plot1中, 功能群优势度随生物量的增加而显著降低。在高强度放牧样地(Plot4、Plot5、Plot6)中, 生活型优势度均随生物量的增加而显著升高。其余多样性指数与生物量无关(图 4)。

|
| 图 4 不同放牧强度区优势度与生物量的关系 Fig. 4 Relationship between Berger-parker index and biomass in different plots |
植物多样性指数是研究群落物种组成结构和定量比较的重要指标[12]。生态学家为简化群落物种研究的复杂性[16]和强调具体多样性测定中的可操作性, 提出了功能多样性的概念[17]。但究竟物种多样性和功能多样性哪个多样性指标可以更好、更快捷地对群落进行定量比较仍然不甚清晰。原因是生态学研究中有很多多样性指数可供选择[12]。功能多样性作为物种组合类群的多少, 不同的研究者有不同的划分方法[68]。但以生活型态[18, 23]和经典的林奈分类学方法[6, 8, 24]进行功能分类被大多数研究者所认可。而不论是物种多样性还是功能多样性, 多样性统计量均可分为丰富度[13]、优势度[14]和均匀性[15]测度。因此, 面对复杂、种类繁多的多样性指数[12], 明晰放牧干扰下不同层次多样性之间的相互关系, 找出放牧干扰下多样性的代表性测度指标就有很重要的实践意义。本研究在青藏高原东缘的高寒草甸连续4 a进行放牧干扰下物种、生活型和功能群多样性之间相关关系的研究。结果表明, 物种丰富度与其余多样性指数之间均呈显著相关关系, 尤其与功能多样性表现极强的相关性。物种丰富度作为某一选定集聚群落内已知物种的种数, 其简单有效的定义和具体实践的可操作性[25], 可将其作为生物多样性比较的代表性测度指标。虽然物种丰富度并不意味着它是生物多样性的唯一测度, 但因为越是物种丰富的群落就存在越多的功能类群, 本研究中物种丰富度与功能多样性也表现出显著相关关系, 其对生态系统功能多样性强大的预测潜能, 也引起研究者的关注[25]。但因为研究者所采用的物种概念不同就会使丰富度的测度受到很大的影响[26]。因而, 在物种概念较为一致的情况下, 物种丰富度可以作为生物多样性测度的代表性指标。
3.2 放牧强度对物种丰富度的影响随放牧年份有累积作用物种多样性作为一门比较性科学[12, 25]。研究者通常想知道在空间尺度上是否一个地区的多样性超过另一个地区?或特定地理区域的自然群落在时间演替过程中多样性如何变化[25]?对于放牧干扰下的草地生态系统, 研究者较多关注放牧强度或时间尺度单一因素影响下的多样性比较[3]。但对于放牧而言, 干扰的空间强度特征及其在时间尺度上的累计效应是同时存在的[12]。而植物本身又具有补偿性生长和每年重新萌发的特性, 因而研究哪些多样性指数随放牧强度在时间尺度上具有累积效应, 才能了解放牧干扰对多样性影响的持续性。本研究分析了放牧强度在放牧年份的累积作用下对物种、生活型和功能群多样性的作用大小。结果表明, 放牧年份和放牧强度仅对物种丰富度具有交互作用, 也就是说放牧强度对物种丰富度的作用随放牧年份具有累积作用。而其余层次的多样性指数虽都随放牧年份延长发生显著变化, 但没有产生随放牧强度变化的时间累积效应。这就能够解释以物种丰富度作为群落物种多样性的代表性测度指标时, 放牧强度能够造成多样性变化的原因。因为物种丰富度随放牧时间的累积在不同的放牧强度区发生明显的分异现象, 低放牧强度区的物种丰富度高于高放牧强度区。物种丰富度(即植物种类的多少)作为多样性测度中最直接的指标, 累积效应的产生意味着研究者必须用长远的眼光来看待某一时刻放牧强度的差异, 关注放牧对多样性产生影响的长时间效应, 而不只是短期的结果。
3.3 高强度放牧干扰下生活型多样性与生物量依然显著相关植物功能类型是具有确定功能性状的一系列植物的组合, 是生态学家为研究植被应对环境变化而引入的生态学概念[17]。功能多样性简化了群落物种研究的复杂性[17], 也因其对群落生产力强大的预测潜能[10], 被研究者广泛用于多样性与生产力关系的研究[6-8]。但也有研究者发现功能多样性与生产力无关[8, 11]。对草地生态系统而言, 放牧作为首要且不可避免的干扰方式[12], 扰乱了多样性与生产力之间的相互关系, 使得物种和功能多样性哪一个对生产力有较显著影响更加不确定。最主要的原因是研究者在野外试验中多采用草地植物的现存量来表示生产力水平[8-10, 19], 而未考虑草地普遍存在且不可避免的放牧强度造成的生物量差异。本研究连续4年对不同层次多样性与生物量的关系进行研究表明, 在不同的放牧强度区丰富度(物种、生活型和功能群)与生物量之间的关系均不显著。而对于均匀度和优势度而言, 仅在高强度放牧区(Plot4、Plot5、Plot6)生活型均匀度随生物量的增加而降低, 生活型优势度随生物量的增加而增加。生活型作为植物适应生态环境而具有的一定形态外貌、结构和习性[23]。在放牧干扰下, 植物群落的生活型调整决定群落能否适应放牧生境而维持生存[23]。本研究中, 在高强度放牧区, 生活型均匀度和优势度对地上生物量表现出积极作用, 更敏感的反映了生物量的变化。而丰富度对生物量的变异不敏感, 原因可能是虽然物种丰富度在放牧强度的时间累积作用下发生分异, 但丰富度(物种和物种组合类群的多少)的变化总是缓慢的。而均匀度和优势度易受放牧干扰的直接影响, 使得能够适应高强度放牧干扰的优势物种和优势类群的比例增加。高强度放牧干扰激发了这一部分植物的生存潜力, 因而高强度放牧区群落优势物种和优势类群的生存优势就越明显[12], 即优势度越高、均匀度越低, 群落在放牧干扰下的生产力也就越高。高强度放牧区生产力的增加主要通过优势物种或优势类群的生存优势来实现。同样是物种组合类群, 为什么是生活型多样性与放牧干扰下的生物量有更强烈的密切关系, 而不是功能群多样性。原因可能是同样作为物种组合类型的指标, 功能群多样性和生活型多样性均可用于简化群落物种研究的复杂性[17]。但功能群是基于生理、生活史或其它与某一生态过程相关以及与物种行为相联系的一些生物学特性来划分的, 且干扰下群落的纯生理性功能分化需在较长时期才能实现[27-28]。而群落生活型即植物的外部形态可以在较短时间内对干扰做出响应[23], 进而影响群落结构, 具有更强的操作性。生活型划分是以植物趋同适应的形态表型为基础[23], 而功能群划分以植物的生理生态特性的趋同性为特征[28]。放牧干扰下的生活型态的表型变化往往先于生理生态特性的变化, 因而生活型多样性在高强度放牧干扰下可以表现与生物量的显著相关关系。生活型多样性可作为放牧干扰下生物量变化快速预测的有效指标。
| [1] | Wu G L, Du G Z, Liu Z H, Thirgood S. Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau. Plant and Soil, 2009, 319(1/2): 115–126. |
| [2] | 杜国祯, 覃光莲, 李自珍, 刘正恒, 董高生. 高寒草甸植物群落中物种丰富度与生产力的关系研究. 植物生态学报, 2003, 27(1): 125–132. DOI:10.17521/cjpe.2003.0019 |
| [3] | Qi W, Guo S Q, Chen X L, Cornelissen J H C, Bu H Y, Du G Z, Cui X L, Li W J, Liu K. Disentangling ecological, allometric and evolutionary determinants of the relationship between seed mass and elevation:insights from multiple analyses of 1355 angiosperm species on the eastern Tibetan Plateau. Oikos, 2014, 123(1): 23–32. DOI:10.1111/more.2014.123.issue-1 |
| [4] | Li W J, Knops J M H, Brassil C E, Lu J F, Qi W, Li J H, Liu M X, Chang S H, Li W L. Functional group dominance and not productivity drives species richness. Plant Ecology & Diversity, 2016, 9(2): 141–150. |
| [5] | Bai Y F, Wu J G, Pan Q M, Huang J H, Wang Q B, Li F S, Buyantuyev A, Han X G. Positive linear relationship between productivity and diversity:evidence from the Eurasian Steppe. Journal of Applied Ecology, 2007, 44(5): 1023–1034. DOI:10.1111/jpe.2007.44.issue-5 |
| [6] | 董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(5): 1472–1483. |
| [7] | Hooper D U, ChapinⅢ F S, Ewel J J, Hector A, Inchausti P, Lavorel S, Lawton J H, Lodge D M, Loreau M, Naeem S, Schmid B, Setälä H, Symstad A J, Vandermeer J, Wardle D A. Effects of biodiversity on ecosystem functioning:a consensus of current knowledge. Ecological Monographs, 2005, 75(1): 3–35. DOI:10.1890/04-0922 |
| [8] | 马文静, 张庆, 牛建明, 康萨如拉, 刘朋涛, 何欣, 杨艳, 张艳楠, 邬建国. 物种多样性和功能群多样性与生态系统生产力的关系——以内蒙古短花针茅草原为例. 植物生态学报, 2013, 37(7): 620–630. |
| [9] | 江小雷, 张卫国, 严林, 王刚. 植物群落物种多样性对生态系统生产力的影响. 草业学报, 2004, 13(6): 8–13. |
| [10] | Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300–1302. DOI:10.1126/science.277.5330.1300 |
| [11] | Thompson K, Askew A P, Grime J P, Dunnett N P, Willis A J. Biodiversity, ecosystem function and plant traits in mature and immature plant communities. Functional Ecology, 2005, 19(2): 355–358. DOI:10.1111/fec.2005.19.issue-2 |
| [12] | 牛钰杰, 杨思维, 王贵珍, 刘丽, 花立民. 放牧干扰下高寒草甸物种多样性指数评价与选择. 应用生态学报, 2017, 28(6): 1824–1832. |
| [13] | McIntosh R P. An index of diversity and the relation of certain concepts to diversity. Ecology, 1967, 48(3): 392–404. DOI:10.2307/1932674 |
| [14] | Tavares-Dias M, Oliveira M S B. Structure of parasites community in Chaetobranchopsis orbicularis (Cichlidae), a host from the Amazon River system in northern Brazil. Parasitology Research, 2017, 116(8): 2313–2319. DOI:10.1007/s00436-017-5539-2 |
| [15] | Smith B, Wilson J B. A consumer's guide to evenness indices. Oikos, 1996, 76(1): 70–82. DOI:10.2307/3545749 |
| [16] | 王长庭, 龙瑞军, 王启基, 景增春, 丁路明. 高寒草甸不同草地群落物种多样性与生产力关系研究. 生态学杂志, 2005, 24(5): 483–487. |
| [17] | 胡楠, 范玉龙, 丁圣彦, 廖秉华. 陆地生态系统植物功能群研究进展. 生态学报, 2008, 28(7): 3302–3311. |
| [18] | Raunkiær C, Egerton F N, Carter H G. The Life Forms of Plants and Statistical Plant Geography. Oxford: The Clarendon Press, 1934. |
| [19] | 王国杰, 汪诗平, 郝彦宾, 蔡学彩. 水分梯度上放牧对内蒙古主要草原群落功能群多样性与生产力关系的影响. 生态学报, 2005, 25(7): 1649–1656. |
| [20] | 贺金生, 方精云, 马克平, 黄建辉. 生物多样性与生态系统生产力:为什么野外观测和受控实验结果不一致?. 植物生态学报, 2003, 27(6): 835–843. |
| [21] | Collins S L, Barber S C. Effects of disturbance on diversity in mixed-grass prairie. Vegetatio, 1986, 64(2/3): 87–94. |
| [22] | Mackenzie A, Virdee S R, Ball A S. Instant Notes in Ecology. Singapore: Springer, 1998. |
| [23] | Whittaker R H. Communities and Ecosystems. New York: Macmillan Company, 1970. |
| [24] | Klein J A, Harte J, Zhao X Q. Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau. Ecology Letters, 2004, 7(12): 1170–1179. DOI:10.1111/ele.2004.7.issue-12 |
| [25] | Magurran A E. Ecological Diversity and Its Measurement. New York: Springer, 1988. |
| [26] | 张德兴. 为什么在物种概念上难以达成共识?. 生物多样性, 2016, 24(9): 1009–1013. DOI:10.17520/biods.2016223 |
| [27] | 鲍雅静, 李政海. 基于能量属性的植物功能群划分方法探索——以内蒙古锡林河流域草原植物群落为例. 生态学报, 2008, 28(9): 4540–4546. |
| [28] | 郭屹立, 卢训令, 丁圣彦. 伊洛河河岸带生态系统草本植物功能群划分. 生态学报, 2012, 32(14): 4434–4442. |