 2018, Vol. 38
2018, Vol. 38文章信息
- 简小枚, 税伟, 王亚楠, 王前锋, 陈毅萍, 江聪, 项子源.
- JIAN Xiaomei, SHUI Wei, WANG Ya'nan, WANG Qianfeng, CHEN Yiping, JIANG Cong, XIANG Ziyuan.
- 重度退化的喀斯特天坑草地物种多样性及群落稳定性——以云南沾益退化天坑为例
- Species diversity and stability of grassland plant community in heavily-degraded karst tiankeng: a case study of Zhanyi tiankeng in Yunnan, China
- 生态学报. 2018, 38(13): 4704-4714
- Acta Ecologica Sinica. 2018, 38(13): 4704-4714
- http://dx.doi.org/10.5846/stxb201706281163
-
文章历史
- 收稿日期: 2017-06-28
- 网络出版日期: 2018-03-16





2. 福州大学生物科学与工程学院, 福州 350116
2. College of Biological Science and Engineering, Fuzhou University, Fuzhou 350116, China
喀斯特天坑(Karst tiankeng)是21世纪初发现的一种与多数漏斗、洼地和洞穴形成机制不同且规模宏大的独特地表负地形, 其通常定义为四周岩壁陡直, 平均宽度和深度均超过100 m的大型塌陷漏斗[1], 由于受到天然屏障的围圈保护, 天坑底部的生境与外界环境相互独立, 具有特殊的水热条件, 在坑底形成了独特的生态系统[2]。中国是名副其实的“世界喀斯特天坑王国”, 截止至2010年, 全世界共发现喀斯特天坑80多个, 其中, 分布于中国的喀斯特天坑达50多个[3], 并出现了许多有代表性的喀斯特天坑群, 如云南沾益天坑群、四川兴文天坑群、广西乐业天坑群等(图 1)。喀斯特天坑群的存在显著影响着区域植物群落的生态过程, 其微生境在全球气候变化下可作为某些植物重要的避难所, 具有一定的保护价值、旅游价值及美学价值[4-6]。

|
| 图 1 全球及中国的喀斯特天坑群分布 Fig. 1 Distribution of global and Chinese karst tiankengs |
Waltham[7]以地质环境发生演化为分类依据, 将喀斯特天坑分为原生天坑、成熟(典型、活动)天坑和退化天坑。近年来对原生天坑和成熟天坑的相关研究越来越多[8-11], 如对云南沾益原生天坑的植物群落研究发现, 坑底存在植物种类丰富、植被类型特殊、区系成分异常等特点[12-13], 广西成熟天坑植物群落具有植物种类丰富、水平分布平衡丰富及生长环境多样等特点[14-15], 但对退化天坑的生物多样性特征及内在机理方面研究较薄弱, 所蕴含的巨大科学价值尚未得到足够重视。退化天坑是指失去地下河行迹或周壁的完整性遭受严重破坏的天坑, 其四周的崖壁渐渐被堆积体掩埋, 生长的树木掩映块石堆积体后形成天坑植物群落的一种天坑类型[16]。目前在全球喀斯特区域上分布的绝大多数天坑都以退化天坑的形式存在, 从天坑的破损程度及深宽比等外观形态上可将退化天坑分为轻度退化天坑、中度退化天坑、重度退化天坑及极重度退化天坑。
退化天坑底部可进入性较强, 受人类干扰较大, 坑底生态系统及其群落分布格局异于原生天坑, 具有较多元的生态类型, 刘守江等[11]对不同溶蚀洼地与退化天坑之间的植物群落物种多样性进行对比发现退化天坑植物群落物种多样性最为丰富, 且不同类型的退化天坑植物群落具有相似性和重现性低的特点[17]。退化天坑相比原生天坑在研究过程中存在较多的不确定性及多重性, 但其作为所有天坑类型中与外界连接最紧密的区域, 其植物多样性研究不仅是探索天坑植物群落特征不可或缺的一部分, 还能为脆弱的喀斯特区域生态恢复提供有力的借鉴。
云南沾益天坑群具有原生-退化天坑完美共生的特点, 且不同退化程度的喀斯特天坑分布较多, 坑底主要分布着草地植物群落。因此, 本文以重度退化的大竹菁天坑坑底草地植物群落为研究对象, 从“轻微人为干扰下自然恢复的重度退化天坑草地植物群落是否具有稳定的群落特征?”这一科学问题出发, 采用野外采样和统计分析相结合的方法, 探寻坑底草地植物群落特征及多样性变化, 从定量的角度分析坑内外植物群落的差异以及天坑植物群落的稳定程度, 完善退化天坑草地现有植被资料, 揭示坑内与坑外植物群落分布格局的差异, 为退化草地的生态恢复和生物多样性保育提供科学依据, 亦为后续原生天坑的植物群落结构和功能研究奠定研究基础。
1 研究区域和方法 1.1 区域概况沾益天坑群, 原生和退化天坑完美共生, 集中成群分布。其不仅是国内独特的景观资源, 也为地质、水文、生态和环境等众多学科提供一个重要的科研场所, 具有极高的研究价值和保护价值[14]。天坑群位于云南省沾益海峰自然保护区内, 地处滇东高原北部, 地理坐标介于103°29′—103°39′E, 25°35′—25°57′N之间(图 2)。该地区气候属于温带高原季风气候向亚热带高原季风气候过渡区, 冬春干旱多风, 夏秋湿润多雨, 年均温在13.8—14 ℃, 年降雨量1073.5—1089.7 mm, 年蒸发量在2069.1 mm, 相对湿度71%, 每年霜日集中在1月, 全年无霜期242 d, 年平均风速2.7 m/s, 风向以西南风为主[13]。该区域独特的气候类型及其特征为沾益区创造了生物多样性和发展的必备条件, 对林木的生长发育十分有利。由于倒石坡的存在, 坑坡形成了森林群落, 而坑底主要分布着草地植物群落。草地主要常见种有狗牙根(Cynodon dactylon)、竹节草(Chrysopogon aciculatus)和龙芽草(Agrimonia pilosa)等。

|
| 图 2 云南省沾益天坑群区位 Fig. 2 Location of Zhanyi tiankeng groups in Yunnan province |
大竹菁退化天坑, 坑外海拔1904 m, 深度123.6 m, 长径455.6 m, 短径达365.0 m, 口部面积1.66×105 km2, 深宽比为0.271, 规模宏大, 是沾益天坑群中典型的重度退化喀斯特天坑(图 3a)。其壁破损面积达51%—80%, 部分岩壁直立, 有大量崩塌堆积体和多个退化坡面, 圈闭化程度较差, 可进入性较强。天坑迄今已退耕40多年, 由于坑壁的遮挡作用, 现有人为干扰较小, 仅有少数羊群放牧干扰, 植被处于自然恢复阶段。由于退化坑壁形成特征及倒石坡的存在, 坑底主要草本植物有竹节草、狼尾草(Pennisetum alopecuroides)、艾(Artemisia argyi)等(图 3b);坑底边缘主要为灌草丛向森林群落的过渡带, 主要分布着密花荚蒾(Viburnum congestum)、雀梅藤(Sageretia thea)等;坑坡主要为森林群落, 其中, 南坡光照、水分等营养条件充足, 植被较为丰富, 主要分布有云南松(Pinus yunnanensis)、油杉(Keteleeria fortunei)等;北坡自然条件较为缺乏, 植被稀疏, 主要分布着密花荚蒾(Viburnum congestum)、小叶栒子(Cotoneaster franchetii)等。

|
| 图 3 大竹菁退化天坑剖面图(a)及示意图(b) Fig. 3 Profile (a) and sketch map (b) of Dazhujing |
在2016年8月, 对大竹菁退化天坑的草地植物群落进行详细调查, 由于四周倒石坡的存在, 大竹菁天坑底面积远小于坑口面积, 且坑底自西向东绵延分布的盲河将坑底部分成南北两侧, 考虑到地形和植被实际分布情况, 采用线路与典型样地调查相结合的方法, 在盲河的上中下段设置南北样线, 在上下两条样线中各布设6个草本样方(南北两侧各3个样方), 由于中段两侧的植被较为茂密, 在该样线中布设8个样方(南北两侧各4个样方), 共设置20个草本样方。此外, 选取坑外喀斯特草地进行草本样方的布设, 共20个草本样方, 用于坑内外植物群落的对比研究。详细记录每个样方内物种的种类、名称、数量、盖度等数据, 并采集部分植物标本, 进行室内植物的识别。本研究中的草本包括各杂类草、蕨类及草质藤本等。
1.3 数据处理与方法本研究采用Excel 2007和SPSS 22.0软件分别进行数据处理和统计分析, 采用t检验法检验两个变量之间的差异显著性(α=0.05), Origin 8.0软件绘图。
1.3.1 植物多样性测度在植物清查和识别的基础上, 分析退化天坑每种植物的相对密度、相对盖度、相对频度及重要值等数量特征, 并测度植物群落内部的α多样性指数[18-21]。
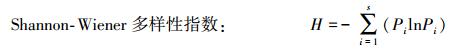
|
(1) |

|
(2) |

|
(3) |

|
(4) |
式中, Pi为第i个种在全体物种中的重要性比例, 如果以个体数量而言, Pi=Ni/N, Ni为第i个种的个体数量, N为总个体数量;S为物种总数。
其次, 本研究对天坑内外不同生境间植物群落的β多样性指数进行测算[22], 文中采用较为简便的植物群落相似性系数进行测度, 研究主要采用Jaccard系数进行计算:

|
(5) |
式中, a、b为不同生境下两植物群落的物种数, c为两植物群落的共有物种数;β范围在(0, 1), 其中, β在[0, 0.25)之间, 表明两群落极不相似;在[0.25, 0.50)之间, 表明两群落中等不相似;在[0.5, 0.75)之间, 两群落中等相似;在[0.75, 1.00)之间, 表明两群落极为相似。
1.3.2 群落总体关联性分析多植物种间的总体关联性有效地反映出退化天坑植物在不同环境中的适应程度, 并能充分的表征植物群落的结构特征。本研究采用零联结模型导出的方差比率法[23-24]定量验证天坑植物群落的总体关联性:
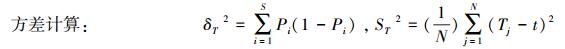
|
(6) |

|
(7) |
式中, S为总物种数, Pi=ni/N, ni为物种i出现的样方数, N为样方总数;Tj为样方j内出现的物种数, t为样方内物种的平均数;VR为群落内物种间的总体关联指数。VR>1表示物种间总体表现为正关联, VR =1, 即所有种间无关联, VR<1表示物种间总体表现为负关联。同时, 采用统计量W来检验VR偏移1的显著程度, W=VR×N, 若种间关联不显著, 则W落入χ0.052(N) < W < χ0.952(N)的概率有90%。
1.3.3 群落稳定性分析采用M. Godron贡献定律法[25], 将退化天坑底部所调查的植物种类按频度大小依次排序, 并转化为相对频度进行逐步累积;其次, 按植物种类排序的顺序将植物种类的总和取倒数, 并依次进行累积, 用百分数表示;以植物种类累积倒数百分比为x轴, 以累积相对频度为y轴, 建立模糊散点平滑曲线;最后, 与直线x+y=100相交, 求交点坐标, 即为所求植物群落的稳定点。植物群落的植物种类累积倒数百分比与累积相对频度百分比越接近20/80, 植物群落就越稳定[26]。

|
(8) |

|
(9) |
通过公式(9)得出交点坐标(x, y), 与20/80比较, 判断退化天坑坑内与坑外植物群落的稳定程度。
2 结果与分析 2.1 退化天坑内外草地的植物群落基本特征 2.1.1 物种组成对大竹菁退化天坑样方调查发现, 从相对密度、相对盖度及相对频度出发, 除了常见的松杉等乔木外, 喀斯特天坑区域的植物组成相对简单, 天坑内外共有草本植物54种, 分属于20科, 49属。其中, 菊科12属13种, 豆科6属6种, 蔷薇科2属4种, 禾本科4属4种, 四大科合计24属27种, 占全部种数的50.0%, 表明四大科在喀斯特天坑草地群落区系中占了重要的地位。其次, 伞形科(Umbelliferae)有3种植物, 薯蓣科(Dioscoreaceae)与天南星科(Araceae)有2种植物, 酢浆草科(Oxalidaceae)、金星蕨科(Thelypteridaceae)、毛茛科(Ranunculaceae)、水龙骨科(Polypodiaceae)、川续断科(Dipsacaceae)、凤尾蕨科(Pteridaceae)、葫芦科(Cucurbitaceae)、爵床科(Acanthaceae)、木犀科(Oleaceae)、茜草科(Rubiaceae)、茄科(Solanaceae)、荨麻科(Urticaceae)、唇形科(Labiatae)等13科均只有1种植物。总体上, 天坑内外的植物群落存在一定差异, 天坑内有14科35属35种, 坑外草地群落有14科30属30种, 坑内外的植物组成表现为多数植物属于少数科, 少数植物属于多数科, 多数植物属于单一种(表 1)。
| 科名Family | 属名Genus | 物种名Species | 相对密度 Relative density |
相对盖度 Relative coverage |
相对频度 Relative frequency |
|||||
| 天坑内Inside | 天坑外Outside | 天坑内Inside | 天坑外Outside | 天坑内Inside | 天坑外Outside | |||||
| 禾本科Gramineae | 金须茅属Chrysopogon | 竹节草Chrysopogon aciculatus | 24.61 | — | 27.59 | — | 9.57 | — | ||
| 狗牙根属Cynodon | 狗牙根Cynodon dactylon | 11.71 | 0.42 | 13.22 | 0.03 | 8.51 | 1.08 | |||
| 狼尾草属Pennisetum | 狼尾草Pennisetum alopecuroides | 30.65 | 28.48 | 30.36 | 44.83 | 12.77 | 19.35 | |||
| 求米草属Oplismenus | 竹叶草Oplismenus compositus | 0.27 | — | 0.54 | — | 1.06 | — | |||
| 菊科Compositae | 白酒草属Conyza | 白酒草Conyza japonica | 0.09 | — | 0.24 | — | 1.06 | — | ||
| 毛连菜属Picris | 毛连菜Picris hieracioides | — | 0.42 | — | 0.14 | — | 1.08 | |||
| 兔儿风属Ainsliaea | 长穗兔儿风Ainsliaea henryi | — | 10.97 | — | 11.54 | — | 8.60 | |||
| 马边兔儿风Ainsliaea angustata | 0.18 | 1.90 | 0.03 | 1.71 | 1.06 | 4.30 | ||||
| 田基黄属Grangea | 田基黄Grangea maderaspatana | 0.09 | — | 0.01 | — | 1.06 | — | |||
| 天名精属Carpesium | 天名精Carpesium abrotanoides | 0.91 | — | 1.69 | — | 2.13 | — | |||
| 火绒草属Leontopodium | 火绒草Leontopodium leontopodioides | — | 1.69 | — | 0.55 | — | 2.15 | |||
| 蒿属Artemisia | 艾Artemisia argyi | 8.05 | — | 8.12 | — | 9.57 | — | |||
| 飞蓬属Erigeron | 一年蓬Erigeron annuus | 0.27 | 0.21 | 0.49 | 0.02 | 1.06 | 1.08 | |||
| 泽兰属Eupatorium | 异叶泽兰Eupatorium heterophyllum | 0.55 | 5.91 | 0.38 | 7.29 | 1.06 | 10.75 | |||
| 橐吾属Ligularia | 橐吾属Ligularia sp. | 0.27 | 2.95 | 0.08 | 1.90 | 1.06 | 4.30 | |||
| 苦荬菜属Ixeris | 苦荬菜Ixeris polycephala | — | 1.27 | — | 1.41 | — | 2.15 | |||
| 豆科Leguminosae | 两型豆属Amphicarpaea | 两型豆Amphicarpaea edgeworthii | 1.19 | — | 1.54 | — | 3.19 | — | ||
| 山蚂蝗属Desmodium | 异叶山蚂蝗Desmodium heterophyllum | 2.01 | 5.91 | 1.57 | 5.30 | 4.26 | 6.45 | |||
| 含羞草属Mimosa | 含羞草Mimosa pudica | 1.01 | — | 0.13 | — | 2.13 | — | |||
| 黄耆属Astragalus | 背扁黄耆Astragalus complanatus | — | 2.32 | — | 0.36 | — | 1.08 | |||
| 决明属CassiaMedicago | 含羞草决明Cassia mimosoides | — | 0.21 | — | 0.02 | — | 1.08 | |||
| 蔷薇科Rosaceae | 龙芽草Agrimonia pilosa | 9.42 | 1.90 | 4.43 | 1.40 | 10.64 | 2.15 | |||
| 龙芽草属Agrimonia | 纤细草莓Fragaria gracilis | — | 9.07 | — | 4.28 | — | 4.30 | |||
| 草莓属Fragaria | 野草莓Fragaria vesca | 0.73 | 0.63 | 0.29 | 0.29 | 1.06 | 1.08 | |||
| 伞形科Umbelliferae | 积雪草属Centella | 积雪草Centella asiatica | 1.65 | — | 1.57 | — | 1.06 | — | ||
| 柴胡属Bupleurum | 马尾柴胡Bupleurum microcephalum | 0.09 | — | 0.10 | — | 1.06 | — | |||
| 天胡荽属Hydrocotyle | 天胡荽Hydrocotyle sibthorpioides | 0.18 | 1.05 | 0.03 | 0.40 | 1.06 | 1.08 | |||
| 酢浆草科Oxalidaceae | 酢浆草属Oxalis | 酢浆草Oxalis corniculata | 1.28 | 1.05 | 0.27 | 0.56 | 3.19 | 2.15 | ||
| 金星蕨科Thelypteridaceae | 毛蕨属Cyclosorus | 毛蕨Cyclosorus interruptus | 1.74 | 0.84 | 5.10 | 0.83 | 5.32 | 2.15 | ||
| 毛茛科Ranunculaceae | 铁线莲属Clematis | 女萎Clematis apiifolia | 0.27 | 1.48 | 0.21 | 0.21 | 2.13 | 1.08 | ||
| 薯蓣科Dioscoreaceae | 薯蓣属Dioscorea | 高山薯蓣Dioscorea henryi | — | 5.70 | — | 2.63 | — | 3.23 | ||
| 云南薯蓣Dioscorea yunnanensis | 0.46 | — | 0.16 | — | 2.13 | — | ||||
| 天南星科Araceae | 天南星属Arisaema | 一把伞南星Arisaema erubescens | — | 0.42 | — | 0.30 | — | 1.08 | ||
| 半夏属Pinellia | 半夏Pinellia ternata | — | 0.42 | — | 0.60 | — | 1.08 | |||
| 水龙骨科Polypodiaceae | 瓦韦属Lepisorus | 扭瓦韦Lepisorus contortus | 0.46 | — | 0.24 | — | 1.06 | — | ||
| 川续断科Dipsacaceae | 川续断属Dipsacus | 川续断Dipsacus asperoides | — | 5.27 | — | 4.15 | — | 5.38 | ||
| 凤尾蕨科Pteridaceae | 凤尾蕨Pteris | 凤尾蕨Pteris cretica | — | 1.69 | — | 3.36 | — | 2.15 | ||
| 葫芦科Cucurbitaceae | 茅瓜属Solena | 茅瓜Solena amplexicaulis | — | 2.53 | — | 0.73 | — | 1.08 | ||
| 爵床科Acanthaceae | 叉花草属Diflugossa | 叉花草Diflugossa colorata | 0.18 | — | 0.53 | — | 2.13 | — | ||
| 木犀科Oleaceae | 素馨属Jasminum | 扭肚藤Jasminum elongatum | — | 0.63 | — | 0.46 | — | 3.23 | ||
| 茜草科Rubiaceae | 拉拉藤属Galium | 猪殃殃Galium aparine | 0.27 | — | 0.03 | — | 1.06 | — | ||
| 茄科Solanaceae | 茄属Solanum | 龙葵Solanum nigrum | 0.09 | — | 0.06 | — | 1.06 | — | ||
| “—”表示物种不存在 | ||||||||||
群落物种组成是反映群落结构变化的重要定量指标, 是群落最重要的基本特征之一。物种重要值研究显示(图 4), 坑内优势种重要值在1.76%—73.77%变化, 依次为狼尾草(73.77%)>竹节草(61.78%)>狗牙根(33.45%)>艾(25.74%)>龙芽草(24.49%)>毛蕨(12.16%)>异叶山蚂蝗(7.84%)等, 而坑外优势种重要值在2%—92.66%变动, 重要值排序依次为狼尾草(92.66%)>长穗兔儿风(31.11%)>异叶泽兰(23.95%)>异叶山蚂蝗(17.65%)>纤细草莓(17.65%)>川续断(14.80%)>高山薯蓣(11.55%)>马边兔儿风(7.91%)>凤尾蕨(7.20%)等。说明退化天坑内外植物群落的建群种和优势种存在明显的差异, 在坑内生境中, 狼尾草、竹节草、狗牙根、艾以及龙芽草等物种是植物群落的优势种或建群种;在坑外生境中, 植物群落的优势种或建群种是狼尾草、长穗兔儿风、异叶泽兰、异叶山蚂蝗以及纤细草莓等物种。可以看出, 天坑内外植物群落的优势种差异较大, 仅狼尾草为坑内外共优势种。

|
| 图 4 天坑内(a)和天坑外部(b)主要物种重要值 Fig. 4 Important value of main species inside (a) and outside (b) tiankeng |
比较天坑内外植物群落多样性指数(图 5), Shannon-Wiener物种多样性指数常被用于研究异质性问题, 坑内的物种多样性指数(1.0302)低于坑外的物种多样性指数(1.2364), 但差异不显著;Pielou群落均匀度指数由Shannon-Wiener指数推导而来, 表示群落均匀度情况, 可看出, 坑外群落均匀度(0.8241)显著高于坑内群落均匀度(0.6981);Simpson优势度指数是最著名的优势度指数之一, 主要用于度量调查群落的集中性, 其常与多样性指数的测度内容相反, 从图 5可以看出, 坑内优势度指数(0.5343)低于坑外群落优势度指数(0.6385), 但差异不显著, 说明坑外群落集中性较高, 异质性较小;Margalef物种丰富度指数表示一个群落或环境中物种数目的多寡, 坑内物种丰富度(0.9401)低于坑外物种丰富度(1.1685), 但差异亦不显著。此外, 本研究还采用Jaccard相似性原理作为不同生境之间相似性的度量标准, 分析坑内外植物群落相似程度。研究发现坑内外的Jaccard系数为0.2037, 说明两群落处于极不相似水平, 坑内外植物群落之间具有较大的生境差异性, 结合天坑的独特负地形考虑, 塌陷型天坑的四周坑壁具有一定隔离作用, 形成的内部独特小气候能为坑内植物群落生长提供异于坑外的生长环境。

|
| 图 5 天坑内部和外部的植物多样性 Fig. 5 Plant diversity inside and outside tiankeng |
采用方差比率法对大竹菁退化天坑内外植物群落的总体关联性进行分析, 结果如表 2所示, 坑外植物群落VR=0.71 < 1, 表明物种总体为负关联, 但W=14.25, 落在(χ0.952 20, χ0.052 20)之间, 并未达到显著性水平, 坑外优势种群处于动态演替的不稳定阶段;坑内植物群落VR=1.18>1, 物种总体为正相关, W=23.64亦落在(χ0.952 20, χ20.05 20)之间, 未达到显著性水平, 说明坑内种间联结较松散, 具有一定的独立性, 正处于不断完善阶段。
| 群落 Community |
方差比率 Variance ratio |
检验统计量W Test statistics W |
χ2临界值 χ2 critical value |
总体关联性 Overall association |
|
| χ0.952 20 | χ0.052 20 | ||||
| 坑外植物群落 Outside-tiankeng community |
0.71 | 14.25 | 10.85 | 31.41 | 不显著负关联 |
| 坑内植物群落Inside-tiankeng community | 1.18 | 23.64 | 不显著正关联 | ||
为进一步判断退化天坑植物群落的稳定程度, 根据M. Godron稳定性测定方法, 将植物群落所有植物进行累计并绘图。得到坑内外植物群落的稳定性图解(图 6)。以二次函数做趋势线模拟, 并结合函数x+y=100求解, 得出坑内植物群落稳定性曲线类型为y=-0.0099x2+1.6887x+23.3972, R2=0.9406, 其交点坐标为32.34/67.66;坑外群落稳定性曲线类型为y=-0.0092x2+1.6489x+22.7768, R2=0.9828, 其交点坐标为32.92/67.08;坑内外整体群落的种类倒数百分率与累积相对频度比值均远离20/80, 表明坑内外群落均处于不稳定状态。

|
| 图 6 天坑内和天坑外草本植物群落的稳定性 Fig. 6 The stability of herbaceous plant community inside and outside tiankeng |
大竹菁是沾益天坑群中较为典型的重度退化天坑, 根据样方调查数据显示, 坑内草地群落有14科35属35种, 坑外有14科30属30种, 坑内物种组成多样, 与坑外草地群落存在一定差异, 说明四周破损坑壁仍能起到一定的隔离作用, 正如刘守江[12]对兴文小岩湾区的洼地和退化天坑的植物多样性比较发现, 退化天坑相比于溶蚀洼地物种组成较丰富, 多样性指数较高。其次, 坑内优势种虽少于坑外植物优势种, 但坑底植物种类较多, 生态系统较稳定, 表明天坑生境为植物提供了较丰富的资源生态位, 具有一定的多样性和复杂性[27-28]。退化天坑相比于其他喀斯特负地形具有其独特的优势:第一, 相比于洼地及漏斗, 四周坑壁起到一定隔离作用, 有效保护了坑底植物群落[10];第二, 相比于落水洞, 退化天坑所受光照及水分较多, 较利于植物生长;如税伟等[17]对兴文小岩湾天坑、飞雾洞等植物特征对比研究发现, 小岩湾天坑植物群落结构较为复杂, 在全球气候变化下, 天坑、漏斗等喀斯特负地形能成为某些物种重要的避难所[29-31]。对天坑植物群落多样性特征研究也发现, 坑内外植物群落分异多样, 坑内相较于坑外多样性及异质性较低, 根据已有的文献资料发现, 大竹菁天坑进行自然生态恢复的年限大于40年, 但目前仅受到轻度的放牧干扰, 而坑外植物群落一直受到适度干扰, 根据中度干扰假说[32], 中等程度干扰能维持较高的物种多样性, 与本研究结果一致。基于相似性原理调查发现, 天坑内外群落相似性系数为0.2037, 处于极不相似水平, 表明生境差异能对物种的分布格局、群落类型产生影响[33]。总的来说, 重度退化天坑的围圈环境在一定程度上产生了地理隔离, 天坑底部独特的微气候能保护相当数量的植物种类, 有利于特有植物生长。
3.2 退化天坑植物种间联结性及稳定性现状退化天坑植物群落总体关联性是种间关系的总体格局, 能够反映天坑植物群落的稳定性和演替方向[34]。总体关联强度越大, 表明天坑植物种间总体互利作用越强, 群落越稳定[35];反之, 种间竞争越强, 越不利于植物生存。通过植物群落种间总体关联性分析, 发现坑内群落总体表现为正相关, 但未达到显著性水平, 说明坑内种间联结较松散, 具有一定的独立性, 正处于不断完善阶段, 说明天坑内植物群落经过40多年的自然恢复, 其植物群落结构仍不稳定。在一定意义上来说, 植物群落的稳定性是种群关联性的体现方式, 植物群落的稳定程度最终是以植物群落中各种群之间的变化关系的表达[36]。本研究采用M. Godron稳定性分析方法, 发现坑内的群落离稳定状态还有一定差距, 坑内植物群落整体还处于相对不稳定状态, 植物群落处于动态调节中, 没有形成多物种共存复杂而稳定的群落。退化天坑虽已弃耕40多年, 但由于退化天坑的可进入性较高, 在恢复的过程中受到少量放牧干扰, 在人类参变作用及自然因素的叠加作用下会导致天坑景观格局恢复困难, 因此未来仍需加强对脆弱天坑群的保育与治理[37]。另一方面, 根据M. Godron稳定性测定, 坑内植物群落稳定性大于坑外植物群落, 说明微生境是影响植物群落稳定程度的因素之一, 塌陷的负地形犹如一个个岛屿, 在坑壁的隔离的作用下, 独特微环境供特有植物群落在坑内生长。种群总体关联性到植物群落稳定状态是一个复杂的生态过程, 本研究将退化天坑的种群关联性和植物群落稳定性放在一起, 二者得出的结论可以相互补充与支撑, 对评价退化天坑现状有很大的实际意义。
4 结论本研究对重度退化天坑的植物群落的多样性特征及稳定性分析发现, 退化天坑内草地群落现有14科35属35种, 坑外有14科30属30种, 物种组成相对多样, 坑内优势种重要值在1.76%—73.77%之间变化, 坑外优势种重要值在2%—92.66%之间变动, 坑内外优势种相差较大, 仅狼尾草一种共优种;从多样性特征分析发现坑外植物多样性指数高于坑内植物多样性指数, 但坑内外差异不显著, 此外, 坑内外植物群落处于极不相似水平, 结构分异多样, 群落之间具有较大的生境差异性;最后, 从种间关联性分析发现, 坑内植物群落呈正关联, 种间联结较松散, 具有一定的独立性, 正处于不断完善阶段, 而坑外优势种群呈负关联, 处于动态演替的不稳定阶段。采用稳定性分析进一步阐释得出, 坑内群落离稳定状态仍有一定差距, 其坑底生境仍较脆弱。
| [1] | 朱学稳, 陈伟海. 中国的喀斯特天坑. 中国岩溶, 2006, 25(S1): 7–24. |
| [2] | Whitten T. Applying ecology for cave management in china and neighbouring countries. Journal of Applied Ecology, 2009, 46(3): 520–523. DOI:10.1111/jpe.2009.46.issue-3 |
| [3] | 税伟, 陈毅萍, 王雅文, 苏正安, 张素. 中国喀斯特天坑研究起源、进展与展望. 地理学报, 2015, 70(3): 431–446. |
| [4] | Vilisics F, Sólymos P, Nagy A, Farkas R, Kemencei Z, Hornung E. Small scale gradient effects on isopods (Crustacea:Oniscidea) in karstic sinkholes. Biologia, 2011, 66(3): 499–505. |
| [5] | Redžić S, Barudanović S, Trakić S, Kulijer D. Vascular plant biodiversity richness and endemo relictness of the karst mountains Prenj-čvrsnica-čabulja in Bosnia and Herzegovina (W. Balkan). Acta Carsologica, 2011, 40(3): 527–555. |
| [6] | Bátori Z, Vojtkó A, Farkas T, Szabó A, Havadtȍi K, Vojtkó A E, Tölgyesi C, Cseh V, Erdȍs L, Maák I E, Keppel G. Large-and small-scale environmental factors drive distributions of cool-adapted plants in karstic microrefugia. Annals of Botany, 2017, 119(2): 301–309. DOI:10.1093/aob/mcw233 |
| [7] | Waltham T. Collapse processes at the tiankengs of Xingwen. Cave and Karst Science, 2005, 32(2/3): 107–110. |
| [8] | 范蓓蓓. 广西大石围天坑群天坑植物群落特征及演替研究[D]. 桂林: 广西师范大学, 2014. |
| [9] | 林宇. 广西大石围天坑群天坑森林物种多样性研究[D]. 桂林: 广西师范大学, 2005. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y784724 |
| [10] | 税伟. 中国兴文世界地质公园绿色发展分析与创新对策. 北京: 光明日报出版社, 2011: 101–118. |
| [11] | 刘守江, 张斌, 杨清伟, 胡翠华, 舒成强. 四川兴文石海小岩湾园区植物群落物种组成与多样性初探. 亚热带植物科学, 2009, 38(1): 37–40. |
| [12] | 李伟云, 向艳辉, 杜宇. 云南沾益海峰自然保护区地下森林群落特征分析. 西南林学院学报, 2001, 21(1): 8–13. |
| [13] | 李伟云, 向艳辉, 杜宇, 吴霞. 云南省沾益天坑内的地下森林. 四川林勘设计, 2001(2): 20–25. |
| [14] | 苏仕林, 黄珂, 马博. 广西乐业大石围天坑群区蕨类植物多样性研究. 湖北农业科学, 2012, 51(5): 948–950. |
| [15] | 苏宇乔, 薛跃规, 范蓓蓓, 莫佛艳, 冯慧喆. 广西流星天坑植物群落结构与多样性. 西北植物学报, 2016, 36(11): 2300–2306. DOI:10.7606/j.issn.1000-4025.2016.11.2300 |
| [16] | 朱学稳, Waltham T. 天坑释义. 中国岩溶, 2006, 25(S1): 35–42. |
| [17] | 税伟, 王兴贵, 张斌, 舒成强, 魏雅丽, 李铁松, 张启春, 程金屏. 兴文侵蚀型天坑的形成演化过程研究. 四川地质学报, 2009, 29(S1): 11–16. |
| [18] | 马克平. 生物群落多样性的测度方法——Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. DOI:10.17520/biods.1994027 |
| [19] | Shannon C E, Weaver W, Wiener N. The Mathematical Theory of Communication. University of Illinois Press, 1998: 623–656. |
| [20] | Margalef R. Information theory in ecology. General Systems, 1958, 3: 36–71. |
| [21] | Pielou E C. The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 1966, 13: 131–144. DOI:10.1016/0022-5193(66)90013-0 |
| [22] | 马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法Ⅱβ多样性的测度方法. 生物多样性, 1995, 3(1): 38–43. |
| [23] | McCulloch C E. Variance tests for species association. Ecology, 1985, 66(5): 1676–1681. DOI:10.2307/1938030 |
| [24] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998–1005. DOI:10.2307/1938071 |
| [25] | Godron M, Daget P, Poissonet J, Poissonet P. Some aspects of heterogeneity in grasslands of Cantal//Patil G P, Pielou E C, Waters W E, eds. Statistical Ecology. Volume 3: Many Species Populations, Ecosystems, and Systems Analysis. University Park, PA: Pennsylvania State University Press, 1971: 397-415. |
| [26] | 黄祥童, 王绍先, 黄炳军, 尹航, 崔凯峰, 赵伟, 范宇光, 顾德峰. 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性. 生态学报, 2015, 35(1): 80–90. |
| [27] | 宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001: 162–186. |
| [28] | 徐斌, 赵哈林, 徐浩, 刘寅虓. 沙地草场放牧试验植物群落的TWINSPAN数量分析. 植物生态学报, 2000, 24(2): 252–256. |
| [29] | Bátori Z, Gallé R, Erdȍs L, Körmöczi L. Ecological conditions, flora and vegetation of a large doline in the Mecsek Mountains (South Hungary). Acta Botanica Croatica, 2011, 70(2): 147–155. DOI:10.2478/v10184-010-0018-1 |
| [30] | Bátori Z, Farkas T, Erdȍs L, Tölgyesi C, Körmöczi L, Vojtkó A. A comparison of the vegetation of forested and non-forested solution dolines in Hungary:a preliminary study. Biologia, 2014, 69(10): 1339–1348. |
| [31] | Bátori Z, Körmöczi L, Erdös L, Zalatnai M, Csiky J. Importance of karst sinkholes in preserving relict, mountain, and wet-woodland plant species under sub-Mediterranean climate:a case study from southern Hungary. Journal of Cave and Karst Studies, 2012, 74(1): 127–134. DOI:10.4311/2011LSC0216 |
| [32] | Roxburgh S H, Shea K, Wilson J B. The intermediate disturbance hypothesis:patch dynamics and mechanisms of species coexistence. Ecology, 2004, 85(2): 359–371. DOI:10.1890/03-0266 |
| [33] | 郝文芳, 杜峰, 陈小燕, 梁宗锁. 黄土丘陵区天然群落的植物组成、植物多样性及其与环境因子的关系. 草地学报, 2012, 20(4): 609–615. DOI:10.11733/j.issn.1007-0435.2012.04.002 |
| [34] | 房飞, 胡玉昆, 张伟, 公延明, 柳妍妍, 杨秀娟. 高寒草原植物群落种间关系的数量分析. 生态学报, 2012, 32(6): 1898–1907. |
| [35] | 奇凯, 张春雨, 侯继华, 赵秀海. 赤峰市沙地油松林草本植物多样性及种间关联动态. 生态学报, 2010, 30(18): 5106–5112. |
| [36] | 张继义, 赵哈林. 植被(植物群落)稳定性研究评述. 生态学杂志, 2003, 22(4): 42–48. |
| [37] | 沈利娜. 峰丛洼地生态系统恢复演替过程中植物群落研究——以广西马山弄拉峰丛洼地为例[D]. 桂林: 广西师范大学, 2008. |