 2018, Vol. 38
2018, Vol. 38文章信息
- 亓琳, 李艳玲, 赵威, 王晓凌.
- QI Lin, LI Yanling, ZHAO Wei, WANG Xiaoling.
- 锶污染下燕麦对土壤酶活性和微生物群落功能多样性的影响
- Effect of Avena sativa L. on soil enzyme activity and microbe functional diversity under strontium pollution
- 生态学报. 2018, 38(13): 4888-4896
- Acta Ecologica Sinica. 2018, 38(13): 4888-4896
- http://dx.doi.org/10.5846/stxb201706261152
-
文章历史
- 收稿日期: 2017-06-26
- 网络出版日期: 2018-03-16





2. 中国科学院生态环境研究中心城市与区域生态国家重点实验室, 北京 100085
2. State key laboratory of urban and regional ecology, Research center for eco-environmental sciences, Chinese academy of sciences, Beijing 100085, China
锶-90是一种放射性金属,为铀-235和钚-239的裂变产物,形成于核武器和核反应堆爆炸[1]。放射性同位素具备重金属和放射性的双重毒性。已有研究表明,放射性核素在环境中与其稳定性同位素在环境中的行为相似。因此,观察稳定锶-88在土壤和植物中的行为,是一个预测模拟其放射性同位素锶-90的方法[2]。
修复污染土壤的最终目标不仅是清除或减少土壤中的污染物,更有价值的是恢复土壤质量[3]。通过评价土壤中活性成分的指标可以判断污染土壤植物修复的效果。土壤微生物对于外界环境的扰动相当敏感,能够有效地表征土壤环境中生物化学过程的强度、方向以及土壤微环境的改变,并对重金属污染相当敏感[4]。土壤酶在土壤中养分循环和能量流动的过程中起着至关重要的作用,与物质与能量的转化、生态环境质量以及植物生长密切相关[5]。BIOLOG技术被表征环境中土壤微生物群落的功能多样性和碳源利用动力学特征,并被广泛应用于评价重金属污染对微生物群落的影响[6]。
燕麦(Avena sativa L.)是主要的高寒作物之一,为上等杂粮。燕麦在华北、西北和青藏高原等高寒及半干旱地区广泛种植。燕麦的生物量大、根系发达、生长速度快以及耐盐胁迫等特点有利于对土壤中污染物的吸附[7]。在我们先前的研究中,比较了小麦、燕麦和大麦的锶富集能力,从26个麦类作物品种中筛选出了锶富集能力最强的燕麦品种—内蒙科艺1号[8]。在本研究中,内蒙科艺1号被选为供试植物,通过研究土壤酶(转化酶、磷酸酶、蛋白酶、脲酶和脱氢酶)活性和微生物群落功能多样性的响应以评价种植燕麦对锶污染土壤的修复效果,以期对锶污染土壤的植物修复研究提供理论依据。
1 材料与方法 1.1 试验材料燕麦(Avena sativa L.)种子购于河南省洛阳市农资市场,选择籽粒饱满、大小一致的燕麦种子,经0.2%次氯酸钠表面消毒20 min后,用水浸种10 min,以去除种子表皮上的病菌,风干。实验用土采集于河南科技大学周山校区的农场土壤表层土(0—20 cm),采用土壤农化分析方法测定其理化性质[9]。供试土壤基本理化性质为:pH值为8.05、有机质含量为12.85 g/kg、全氮含量为0.96 g/kg、速效磷含量12.52 mg/kg、速效钾含量155.13 mg/kg、阳离子交换量为19.23 cmol/kg、锶含量为25.06 mg/kg。
1.2 试验设计采用土培盆栽试验,种植于塑料盆中(高=16 cm,底径=12 cm,口径=22 cm)。每盆中装的风干土2 kg。将分析纯试剂SrCl2· 6H2O溶于水中制成溶液,然后将该溶液均匀地拌入土壤中,设置3个Sr2+浓度梯度,分别为0,75,225 mg/kg,其中Sr2+浓度为0的土壤为对照组。土壤本底值为25.06 mg/kg,将锶处理浓度设置为本底值的4倍和10倍[10-11]。因此,最终土壤中的金属锶浓度为25 mg/kg(CK),100 mg/kg(L)和250 mg/kg(H)。金属锶在土壤中“老化”处理30 d。每盆播种20粒种子,出苗后定苗,每盆12株。本试验设置6个处理包括:1)CK、2)L、3)H、4)CK+燕麦、5)L+燕麦、6)H+燕麦。所有处理设置5个重复。浇灌期间通过称重法添加自来水,保持田间持水量的60%。在锶处理30 d后采样,取燕麦根际土。用土钻将每盆中燕麦的根系从土壤中挖出,尽量保持根系的完整性,采用“抖根法”去掉和根系结合松散的土体土,取与根系紧密结合的土壤。将一盆中获取的根际土混合视为一个样品。根际土样过2 mm筛以除去杂物,然后置于4 ℃下保存,待测[12-13]。
1.3 土壤酶活性的测定转化酶活性测定采用3, 5-二硝基水杨酸比色法,蛋白酶活性测定采用茚三酮比色法,磷酸酶活性测定采用对硝基苯磷酸二钠(pNPP)比色法,脲酶活性测定采用靛酚蓝比色法,脱氢酶活性测定采用氯化三苯基四氮唑(TTC)法[14]。
1.4 土壤微生物功能多样性分析称取相当于风干土10 g的土样加入到装有100 mL的0.85%无菌氯化钠(NaCl)的三角瓶中,室温震荡60 min(200 r/min),采用10倍稀释法,然后用NaCl溶液稀释至浓度10-3。将稀释液接种到Biolog生态板(ECO MicroPlate,美国Matrix Technologies Corporation生产)中,每孔加150 μL。将接种好的ECO板装入(25±1) ℃的恒温暗箱培养,每隔12 h用ELISA(Enzyme-linked Immunosorbent Assay,酶联免疫吸附测定)读取光密度值(590 nm),连续培养144 h[15]。
1) 采用孔平均颜色变化率法(Average Well Color Development,AWCD)测定土壤微生物利用单一碳源的能力。

|
式中,Ci为每个孔的光密度值,Ri为对照孔的光密度值,31为碳源总数。
2) 群落Shannon指数(H)
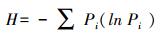
|
式中,Pi=(Ci-Ri)/ ∑(Ci-Ri)。表示有碳源的孔与对照孔A1的光密度值之差与整板总差的比值。
3) 碳源利用丰富度指数(S)
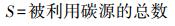
|
4) Shannon均匀度指数(E)

|
5) Shannon优势度指数(D)

|
本试验采用BIOLOG微平板培养72 h的数据分析微生物代谢多样性。
1.5 统计分析以锶添加量为固定因素(自变量),对所有参数(因变量)进行单因素方差分析。处理之间差异的显著性在P<0.05概率水平以最小显著性差异(LSD)进行判定。所有统计分析都是用SPSS软件(version 13.0;SPSS Institute Chicago,IL,USA)来进行。各图表中的数据均为5个重复的平均值。
2 结果 2.1 在锶污染下燕麦对土壤酶活性的影响如表 1所示,在L和H处理中,转化酶活性与对照相比分别减少了7%和21%,磷酸酶活性与对照相比分别减少了13%和17%,蛋白酶活性与对照相比分别减少了5%和13%,脲酶活性与对照相比分别减少了16%和29%,脱氢酶活性与对照相比分别减少了6%和22%。这一结果表明金属锶胁迫显著抑制了土壤酶活性,抑制效应随着金属锶浓度的升高而增强。
| 处理 Treatments |
转化酶Invertase / (mg g-1 h-1) | 磷酸酶Phosphatase / (μg g-1 h-1) | 蛋白酶Protease / (mg g-1 h-1) | 脲酶Urease / (mg g-1 h-1) | 脱氢酶Dehydrogenase / (mg g-1 h-1) |
| CK | 1.23±0.02a | 0.71±0.02a | 0.64±0.02a | 0.31±0.01b | 1.42±0.02b |
| L | 1.15±0.01b | 0.62±0.02c | 0.61±0.01b | 0.26±0.02c | 1.33±0.02c |
| H | 0.97±0.02c | 0.55±0.01d | 0.54±0.016c | 0.24±0.01d | 1.11±0.03e |
| CK+燕麦CK+Oat | 1.25±0.02a | 0.73±0.04a | 0.65±0.03a | 0.35±0.02a | 1.52±0.03a |
| L+燕麦L+Oat | 1.23±0.02b | 0.68±0.02b | 0.63±0.02b | 0.29±0.02b | 1.48±0.02b |
| H+燕麦H+Oat | 1.01±0.01c | 0.59±0.03d | 0.56±0.01c | 0.22±0.01c | 1.22±0.02d |
| CK:对照,control check;L:低浓度锶,low concentration of strontium;H:高浓度锶,high concentration of strontium; 不同小写字母表示不同处理之间差异显著(P < 0.05) | |||||
在CK处理中种植燕麦后,种植燕麦处理与未种植处理相比,5种酶的活性增加。与CK相比,转化酶、磷酸酶和蛋白酶活性分别增加了2%、3%和2%,但未达到显著性差异(P>0.05);而脲酶和脱氢酶活性分别增加了13%和7%,处理之间达到显著性差异(P<0.05)。在L处理中种植燕麦后,与L处理相比,转化酶、磷酸酶、蛋白酶、脲酶和脱氢酶活性分别增加了7%、10%、3%、12%和11%;除蛋白酶活性未达到显著性差异(P>0.05),其他4种酶活性在处理之间差异显著(P<0.05)。在H处理中种植燕麦后,与H处理相比,转化酶、磷酸酶、蛋白酶、脲酶和脱氢酶活性分别增加了4%、7%、4%、9%和10%;转化酶、磷酸酶和蛋白酶活性在处理之间差异不显著(P>0.05),但脲酶和脱氢酶活性在处理之间达到显著性差异(P<0.05)。
2.2 在锶污染下燕麦对AWCD的影响光密度平均颜色变化率(AWCD)能够被用来比较土壤微生物利用总体碳源的能力,AWCD越大,表明微生物的代谢活性越强。如图 1所示,随着培育时间的延长,AWCD逐渐升高。添加金属锶处理与对照相比,其AWCD的排列顺序为:CK>L>H。种植燕麦与无种植处理相比,其AWCD的排列顺序为:CK+燕麦>CK;L+燕麦>L;H+燕麦>H。这一结果表明在锶胁迫条件下种植燕麦能显著恢复微生物对碳源的利用能力。在24h之前AWCD的值一直很小,不同处理间的AWCD值接近于零,表明在培育24 h之内碳源基本上没有被利用。在温育24 h之后,AWCD大幅度增加,表明此后碳源被大量消耗。其中CK和CK+燕麦处理AWCD增加的速度较快,而H处理AWCD增加的速度较慢。综合所有的处理的AWCD,其排序为:CK+燕麦>CK>L+燕麦>H+燕麦>L>H。

|
| 图 1 不同处理AWCD随培育时间的变化 Fig. 1 AWCD changes with incubation time in different treatments 注:CK:对照,control check;L:低浓度锶,low concentration of strontium;H:高浓度锶,high concentration of strontium |
从图 2可以看出,土壤微生物对碳源利用的Shannon多样性指数(H)在L和H处理中显著最低(P<0.05)。Shannon均匀度指数(E)在CK+燕麦处理中最高(P<0.05)。丰富度指数(S)在所有处理中的表现为:CK、CK+燕麦>L、L+燕麦>H+燕麦>H。而Shannon均匀度指数(E)与上述指数有所不同,表现为H>L>L+燕麦、H+燕麦>CK>CK+燕麦。在种植燕麦处理中,相对于L+H处理,L+燕麦和H+燕麦处理显著提高了土壤微生物群落中物种多样性和丰富度(P<0.05),而均匀度和优势度在种植和无种植处理之间无显著性差异(P>0.05)。

|
| 图 2 锶胁迫下燕麦对土壤微生物群落多样性指数的影响 Fig. 2 Effect of oats on the diversity indices of soil microbial communities under Sr stress |
如表 2所示,对培养72 h的31种碳源利用情况进行主成分分析,共提取3个主成分:第一主成分(PC1)、第二主成分(PC2)和第三主成分(PC3),分别可以解释所有变量的54.24%、24.28%和21.48%。3个主成分累积方差贡献率达到100%,可以全面地反映31个变量的特征,并据其得分来表征微生物群落碳源代谢特征。不同的处理中,CK+燕麦在PC1上得分最高,L+燕麦在PC2上得分最高,H+燕麦在PC3上得分最高。
| 处理 Treatment |
主成分1 PC1 |
主成分2 PC2 |
主成分3 PC3 |
| CK | 0.079 | 0.023 | 0.827 |
| L | -3.123 | -2.321 | -1.223 |
| H | -5.581 | -2.222 | -3.221 |
| CK+燕麦CK+Oat | 5.234 | -2.342 | -1.324 |
| L+燕麦L+Oat | 1.321 | 3.214 | -1.253 |
| H+燕麦H+Oat | -0.093 | -1.238 | 3.789 |
| 特征值Eigenvalues | 16.95 | 8.351 | 5.982 |
| 贡献率Contribution | 54.24 | 24.28 | 21.48 |
| 累计贡献率Cumulative contribution | 16.95 | 78.52 | 100 |
因子载荷反映了主成分与碳源利用的相关程度,因子载荷绝对值越接近于1,表示该碳源对主成分的影响越大。如表 3所示,与第一主成分相关性较高的碳源有14个,主要包括酚酸类化合物:4-羟基苯甲酸;胺类化合物:腐胺;聚合物:吐温40、吐温80和α-环式糊精;氨基酸:L-苯丙氨酸;羧酸类化合物:丙酮酸甲脂、D-半乳糖醛酸、γ-羟基丁酸;碳水化合物:D-半乳糖内酯、I-赤藻糖醇、D-甘露醇、N-乙酰基-D-葡萄胺、D-甘露醇。与第二主成分相关性较高的碳源有5个,主要包括酚酸类化合物:2-羟基苯甲酸;胺类化合物:苯乙基胺;聚合物:肝糖;氨基酸:L-丝氨酸;碳水化合物:D-木糖。与第三主成分相关性较高的碳源有4个,主要包括羧酸类化合物:D-苹果酸;聚合物:肝糖;羧酸类化合物:D-葡萄胺酸;碳水化合物:葡萄糖-1-磷酸盐。
| 碳源类别 Carbon source guild |
底物 Substrates |
主成分1 PC1 |
主成分2 PC2 |
主成分3 PC3 |
| 酚酸类化合物Phenolic acids | 2-羟基苯甲酸 | 0.496 | 0.763 | 0.214 |
| 4-羟基苯甲酸 | 0.962 | -0.213 | -0.092 | |
| 胺类化合物Amines/amids | 苯乙基胺 | 0.223 | 0.823 | -0.222 |
| 腐胺 | 0.982 | 0.263 | 0.223 | |
| 聚合物Polymers | 吐温40 | 0.987 | 0.341 | 0.034 |
| 吐温80 | 0.999 | 0.046 | 0.031 | |
| α-环式糊精 | 0.932 | -0.286 | -0.250 | |
| 肝糖 | -0.238 | 0.998 | 0.821 | |
| 氨基酸Amino acids | L-精氨酸 | 0.633 | 0.655 | -0.512 |
| L-天冬酰胺酸 | 0.296 | -0.763 | -0.114c | |
| L-苯基丙氨酸 | 0.962 | -0.213 | -0.092 | |
| L-丝氨酸 | 0.223 | 0.823 | -0.222 | |
| L-苏氨酸 | 0.143 | 0.321 | 0.251 | |
| 甘氨酰-L-谷氨酸 | 0.952 | 0.243 | 0.203 | |
| 羧酸类化合物Carboxylic acids | 丙酮酸甲脂 | 0.927 | 0.311 | 0.024 |
| D-半乳糖醛酸 | 0.997 | 0.046 | -0.031 | |
| γ-羟基丁酸 | 0.942 | -0.216 | -0.160 | |
| D-葡萄胺酸 | -0.134 | -0.197 | 0.912 | |
| 衣康酸 | 0.431 | 0.521 | -0.312 | |
| α-丁酮酸 | 0.369 | 0.112 | 0..104 | |
| D-苹果酸 | 0.942 | -0.153 | -0.051 | |
| 碳水化合物Carbohydrates | β-甲基-D-葡萄糖苷 | 0.396 | 0.563 | -0.114 |
| D-半乳糖内酯 | 0.954 | -0.203 | -0.029 | |
| D-木糖 | 0.312 | 0.933 | -0.122 | |
| I-赤藻糖醇 | 0.982 | 0.263 | 0.223 | |
| D-甘露醇 | 0.987 | 0.341 | 0.034 | |
| N-乙酰基-D-葡萄胺 | 0.999 | 0.046 | 0.031 | |
| D-纤维二糖 | 0.962 | -0.186 | -0.023 | |
| 葡萄糖-1-磷酸盐 | -0.138 | -3.818 | 0.921 | |
| α-D-乳糖 | 0.513 | 0.524 | -0.478 | |
| D, L-α-甘油 | 0.112 | 0.231 | 0.243 |
如表 4所示,金属锶添加处理显著降低了碳水化合物、羧酸类碳源的利用率。在L处理中,碳水化合物和羧酸类碳源的利用率分别下降了9%和34%;在H处理中,碳水化合物和羧酸类碳源的利用率分别下降了25%和47%;且达到显著性差异(P<0.05)。聚合物类、氨基酸类和酚类碳源的利用也稍有下降,但与CK相比差异不显著(P>0.05)。与未种植处理相比(CK、L和H),种植燕麦显著增加了羧酸类和聚合物类碳源的利用率。在CK处理种植燕麦后,羧酸类和聚合物类碳源的利用率分别增加了19%和31%;在L处理种植燕麦后,羧酸类和聚合物类碳源的利用率分别增加了24%和41%;在H处理种植燕麦后,羧酸类和聚合物类碳源的利用率分别增加了30%和38%,且达到显著性差异(P<0.05)。碳水化合物、聚合类、酚类和氨基酸类碳源的利用也稍有增加,但与L和H相比差异不显著(P>0.05)。综合表 3和表 4可知,碳水化合物、聚合物和羧酸类化合物是燕麦种植处理下根际土壤微生物利用的主要碳源。
| 处理 Treatment |
酚酸类化合物 Phenolic acids |
胺类 Amines/amids |
聚合物类 Polymers |
氨基酸类 Amino acids |
羧酸类 Carboxylic acids |
碳水化合物类 Phenolic acids |
| CK | 0.23±0.01a | 0. 27±0.01a | 054±0.02d | 0.74±0.02a | 0.62±0.02b | 0.32±0.01a |
| L | 0.22±0.02a | 0.24±0.02a | 0.46±0.01e | 0.68±0.03b | 0.41±0.01d | 0.29±0.02b |
| H | 0.22±0.01a | 0.22±0.01a | 0.42±0.01f | 0.58±0.03c | 0.33±0.03e | 0.24±0.02b |
| CK+燕麦CK+Oat | 0.27±0.01a | 0.28±0.02a | 0.71±0.02a | 0.79±0.03a | 0.74±0.01a | 0.33±0.02a |
| L+燕麦L+Oat | 0.26±0.01a | 0.25±0.01a | 0.65±0.01b | 0.69±0.02b | 0.51±0.03c | 0.32±0.02a |
| H+燕麦H+Oat | 0.25±0.01a | 0.23±0.02a | 0.58±0.01c | 0.63±0.03c | 0.43±0.02d | 0.26±0.01b |
为了更清晰的了解不同处理间在土壤微生物群落代谢的特征,对培养72 h的31种碳源的利用情况进行聚类分析。聚类分析根据样本的数据特征按照性质上的亲疏程度进行自动分类,类别内部的样本在特征上具有相似性,不同类别间的样本特征差异较大。从图 3可以看出,当聚类距离≤5时,L+燕麦和H+燕麦处理聚为一类,L和H处理聚为一类,CK和CK+燕麦处理聚为一类。当聚类距离≤10时,L+燕麦、H+燕麦、L和H处理聚为一类,而CK和CK+燕麦处理聚为一类。说明L+燕麦、H+燕麦、L和H处理中微生物群落代谢特性趋于一类,并且L+燕麦和H+燕麦处理在聚类关系上趋同性更强,而CK和CK+燕麦处理与其他处理间差异较大。

|
| 图 3 土壤微生物碳源利用聚类分析 Fig. 3 Cluster analysis of carbon utilization by soil microbial communities |
土壤酶系统是土壤有机体中的代谢动力,能催化土壤生态系统中生物化学过程,表征土壤的生态功能、土壤养分转化进程的强度和方向[16]。土壤转化酶、磷酸酶、蛋白酶、脲酶和脱氢酶参与土壤中营养物质转化,对土壤质量和健康状况非常敏感。土壤污染日益严重,土壤酶常用来表征污染物潜在毒性。土壤酶活性变化可以作为土壤重金属污染水平的指标[17]。重金属离子与底物竞争酶活性中心,结合于酶蛋白的必需基团巯基和咪唑基团形成了稳定的金属螯合物,或者重金属对土壤微生物的生长和代谢过程产生抑制作用[18]。本研究中土壤酶活性的降低可能有两个原因:锶与土壤酶蛋白的关键基团和配位键结合从而使酶失活,另一方面锶胁迫下微生物体内酶的合成和分泌减少。重金属通过干扰功能,使蛋白变性和破坏细胞膜完整性,以影响土壤微生物的新陈代谢。土壤酶来自动、植物和微生物的活体或残体的分泌和释放,对土壤中养分的循环至关重要[19]。种植燕麦处理显著提高了土壤脲酶和脱氢酶活性。这一结果说明种植燕麦可以显著地改善锶处理对土壤酶的抑制作用。一方面,燕麦通过富集作用,能够吸收锶进入到植物体内,并清除土壤一定比例的锶,减轻土壤中锶的污染程度。根系分泌物作用于土壤微生境,可以改善土壤环境和提高酶活性;另一方面,土壤酶活性的变化间接表明某些营养物质的转化情况,较高的土壤酶活性在一定程度上保证燕麦生长所需的营养。
3.2 在锶污染下燕麦对土壤微生物群落功能多样性的影响基于微生物代谢的Biolog微平板法,在环境微生物群落研究应用广泛。Biolog技术在污染物对微生物生存环境的扰动评价以及环境的修复效果评价等方面发挥着重要作用。研究表明微生物群落结构和代谢活性与污染物的种类和程度密切相关,重金属暴露浓度与微生物代谢活性之间呈现负相关关系,并能降低微生物群落功能多样性[20]。由于Biolog-ECO板在培育过程中颜色的变化可能是处理土壤之间微生物群落组成的差异造成的,也可能是由于微生物群落大小的差异引起的[21]。因此,比较Shannon多样性指数对于评价种植燕麦对锶污染土壤的修复效果非常有必要。土壤微生物对碳源利用的Shannon多样性指数(H)在L和H处理中最低(P<0.05)。Shannon均匀度指数(E)在CK+燕麦处理中最高(P<0.05)。丰富度指数(S)在所有处理中的表现为:CK、CK+燕麦>L、L+燕麦>H+燕麦>H。这一结果说明重度锶污染显著抑制了土壤微生物群落的丰富度、多样性及均匀度。而Shannon优势度指数(D)与上述指数有所不同,表现为H>L>L+燕麦、H+燕麦>CK>CK+燕麦。这一结果表明在重度锶污染条件下,土壤中生物群落分异演化出优势种以适应金属锶胁迫。在种植燕麦处理中,相对于L+H处理,L+燕麦和H+燕麦处理显著提高了土壤微生物群落中物种多样性和丰富度(P<0.05),而均匀度和优势度在种植和无种植处理之间无显著性差异(P>0.05)。这一结果表明种植燕麦显著提高了土壤微生物群落的丰富度和多样性。种植燕麦在一方面清除了土壤中一定量的金属锶,减轻了土壤污染程度;另一方面通过凋落物和释放分泌物为土壤微生物提供碳源,改善土壤微环境,增加微生物碳源的可利用性,由此增加土壤微生物群落多样性指数[22]。
主成分分析采用降维的思路,分析在不同锶浓度处理下,种植燕麦对土壤微生物碳源利用的影响是否存在差异。郭星亮等[23]研究了重金属污染对铜川煤矿矿区微生物群落代谢的影响,主成分分析的结果表明重金属污染显著抑制了微生物的代谢活性,主要体现在糖类和氨基酸类碳源的利用上,这与本文的研究结果较为一致。不同的锶浓度对土壤微生物群落的影响不同。主成分分析表明种植燕麦对金属锶污染土壤中微生物的碳源底物的利用能力相关。在第一主成分上,解释了大部分的变异,CK+燕麦处理对土壤微生物群落功能影响最大。在第二主成分上,L+燕麦处理得分最高,说明高浓度的锶处理影响土壤微生物群落功能。在第三主成分上,H+燕麦处理得分最高,说明在金属锶污染土壤中种植燕麦能够影响土壤微环境,改变其微生物群落结构,最终使锶污染土壤中的微生物碳源代谢发生改变。重金属锶进入土壤后,改变了土壤微环境,为了适应重金属锶的胁迫环境,微生物群落的组成和结构发生了分异。在重金属污染条件下,土壤微生物群落功能多样性显著降低,能够代谢相关碳源作为底物的微生物数量相对减少,最终微生物对单一碳源底物的利用能力被减弱[24]。因子载荷反映了主成分与碳源利用的相关程度,因子载荷绝对值越接近于1,表示该碳源对主成分的影响越大。碳水化合物、聚合物和羧酸类化合物是燕麦处理下根际土壤微生物利用的主要碳源。这一结果与碳源利用率中的结果相一致。锶添加处理显著降低了碳水化合物和羧酸类碳源的利用率,且达到了显著性差异。聚合物类、氨基酸类和酚类碳源的利用率也稍有下降,但与CK相比差异不显著。燕麦通过其根系分泌和释放代谢产物至土壤中为微生物的生长和繁殖提供了营养和能源,促进微生物大量繁殖并增强微生物代谢活性。燕麦显著增加了羧酸类和聚合物类碳源的利用率。其他类碳源的利用率也稍有增加,但与L和H相比差异不显著。在锶污染土壤中种植燕麦,土壤中微生物群落代谢活性增强,微生物群落利用碳源的能力增加。在聚类分析中,CK和CK+燕麦分为一类,L和H分为一类,L+燕麦和H+燕麦分为一类。CK和CK+燕麦类别和L+燕麦和H+燕麦类别的欧式距离较小,且与L和H类别的欧式距离较大。这一结果说明土壤锶污染与土壤微生物群落的代谢特性关系密切,而种植燕麦显著影响了土壤微环境,与土壤酶活性和主成分分析的结果相一致。
4 结论种植燕麦显著提高锶污染土壤中脲酶和脱氢酶活性,并显著提高锶污染土壤微生物对31种碳源的平均利用率(AWCD)、锶污染土壤微生物群落的Shannon多样性指数(H)、Shannon均匀度指数(E)和碳源利用丰富度指数(S)。主成分分析表明种植燕麦影响了锶污染土壤微生物群落功能多样性,增加了羧酸类和聚合类碳源的利用率。基于以上结果,在锶污染土壤中,燕麦显著提高了酶活性,改善了微生物功能多样性。
| [1] | Wang X, Chen C, Wang J L. Phytoremediation of strontium contaminated soil by Sorghum bicolor (L.) Moench and soil microbial community-level physiological profiles (CLPPs). Environmental Science and Pollution Research, 2017, 24(8): 7668–7678. DOI:10.1007/s11356-017-8432-8 |
| [2] | Kato M, Okada Y, Hirai S, Minai Y, Saito S, Shibukawa M. Comparative analysis of distributions of radioactive cesium and potassium and stable cesium, potassium, and strontium in brown rice grains contaminated with radioactive materials released by the Fukushima Daiichi Nuclear Power Plant accident. Journal of Radioanalytical and Nuclear Chemistry, 2016, 310(1): 247–252. DOI:10.1007/s10967-016-4824-3 |
| [3] | Margon A, Mondini C, Valentini M, Ritota M, Leita L. Soil microbial biomass influence on strontium availability in mine soil. Chemical Speciation & Bioavailability, 2013, 25(2): 119–124. |
| [4] | 李韵诗, 冯冲凌, 吴晓芙, 石润. 重金属污染土壤植物修复中的微生物功能研究进展. 生态学报, 2015, 35(20): 6881–6890. |
| [5] | 徐磊, 周静, 梁家妮, 崔红标, 陶美娟, 陶志慧, 祝振球, 黄林. 巨菌草对Cu、Cd污染土壤的修复潜力. 生态学报, 2014, 34(18): 5342–5348. |
| [6] | 宋收, 陈晓明, 肖伟, 伍迪, 郝希超, 张祥辉, 张倩, 罗学刚. 基于BIOLOG指纹解析土壤可培微生物对铀污染的响应. 核农学报, 2016, 30(6): 1169–1177. DOI:10.11869/j.issn.100-8551.2016.06.1169 |
| [7] | 刘慧军, 刘景辉, 于健, 徐胜涛, 史吉刚. 土壤改良剂对燕麦土壤理化性状及微生物量碳的影响. 水土保持学报, 2012, 26(5): 68-72, 77-77. |
| [8] | Qi L, Qin X L, Li F M, Siddique K H M, Brandl H, Xu J Z, Li X G. Uptake and distribution of stable strontium in 26 cultivars of three crop species:oats, wheat, and barley for their potential use in phytoremediation. International Journal of Phytoremediation, 2015, 17(3): 264–271. DOI:10.1080/15226514.2014.898016 |
| [9] | 鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2001. |
| [10] | 杨叶, 陈珂, 朱靖. 施加钙对锶胁迫下麻疯树生长及生理生化的影响. 核农学报, 2015, 29(2): 405–411. DOI:10.11869/j.issn.100-8551.2015.02.0405 |
| [11] | 曾峰. 高浓度核素污染土壤修复植物筛选及肥料对植物修复的影响[D]. 绵阳: 西南科技大学, 2015. http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=D651919 |
| [12] | 金凌波, 周峰, 姚涓, 姜大刚, 梅曼彤, 穆虹. 磷高效转基因大豆对根际微生物群落的影响. 生态学报, 2012, 32(7): 2082–2090. |
| [13] | 冯晓敏, 杨永, 任长忠, 胡跃高, 曾昭海. 燕麦/大豆和燕麦/花生间作对根际土壤固氮细菌多样性与群落结构的影响. 中国农业大学学报, 2016, 21(1): 22–32. |
| [14] | 关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986. |
| [15] | 孙凯宁, 杨宁, 王克安, 吕晓惠, 柴秀乾. 山药连作对土壤微生物群落及土壤酶活性的影响. 水土保持学报, 2015, 22(6): 95–98. |
| [16] | 张涪平, 曹凑贵, 李苹, 次仁央金, 高超, 通乐嘎, 李成芳. 藏中矿区重金属污染土壤的微生物活性变化. 生态学报, 2010, 30(16): 4452–4459. |
| [17] | Xian Y, Wang M, Chen W P. Quantitative assessment on soil enzyme activities of heavy metal contaminated soils with various soil properties. Chemosphere, 2015, 139: 604–608. DOI:10.1016/j.chemosphere.2014.12.060 |
| [18] | 韩桂琪, 王彬, 徐卫红, 陈贵青, 王慧先, 张海波, 张晓璟, 熊治庭. 重金属Cd、Zn、Cu、Pb复合污染对土壤微生物和酶活性的影响. 水土保持学报, 2010, 24(5): 238–242. |
| [19] | 段德超, 于明革, 施积炎. 植物对铅的吸收、转运、累积和解毒机制研究进展. 应用生态学报, 2014, 25(1): 287–296. |
| [20] | 张旭龙, 马淼, 吴振振, 张志政, 高睿, 石灵玉. 不同油葵品种对盐碱地根际土壤酶活性及微生物群落功能多样性的影响. 生态学报, 2017, 37(5): 1659–1666. |
| [21] | 鲁顺保, 张艳杰, 陈成榕, 徐志红, 郭晓敏. 基于BIOLOG指纹解析三种不同森林类型土壤细菌群落功能差异. 土壤学报, 2013, 50(3): 618–623. |
| [22] | Sumi H, Kunito T, Ishikawa Y, Sato T, Park H D, Nagaoka K, Aikawa Y. Plant roots influence microbial activities as well as cadmium and zinc fractions in metal-contaminated soil. Chemistry and Ecology, 2015, 31(2): 105–110. DOI:10.1080/02757540.2014.950570 |
| [23] | 郭星亮, 谷洁, 陈智学, 高华, 秦清军, 孙薇, 张卫娟. 铜川煤矿区重金属污染对土壤微生物群落代谢和酶活性的影响. 应用生态学报, 2012, 23(3): 798–806. |
| [24] | Ding Z L, Wu J P, You A Q, Huang B Q, Cao C G. Effects of heavy metals on soil microbial community structure and diversity in the rice (Oryza sativa L. subsp. Japonica, Food Crops Institute of Jiangsu Academy of Agricultural Sciences) rhizosphere. Soil Science and Plant Nutrition, 2017, 63(1): 75–83. DOI:10.1080/00380768.2016.1247385 |