 2018, Vol. 38
2018, Vol. 38文章信息
- 张广帅, 于秀波, 张全军, 李雅, 刘宇, 段后浪.
- ZHANG Guangshuai, YU Xiubo, ZHANG Quanjun, LI Ya, LIU Yu, DUAN Houlang.
- 鄱阳湖湿地土壤微生物群落结构沿地下水位梯度分异特征
- Variation in the distribution of soil microbial community structure along ground water level gradients in the Poyang Lake Wetland
- 生态学报. 2018, 38(11): 3825-3837
- Acta Ecologica Sinica. 2018, 38(11): 3825-3837
- http://dx.doi.org/10.5846/stxb201705220946
-
文章历史
- 收稿日期: 2017-05-22
- 网络出版日期: 2018-03-02





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
湿地是陆地生态系统和水域生态系统相互作用形成的自然综合体和独特生态系统类型, 在维护区域生态平衡和生物多样性保护等方面具有重要作用[1], 故而被称为“地球之肾”和“生态超市”[2]。由于湿地生态系统独特的环境条件和丰富的物种多样性, 物质交换和能量流动过程十分复杂, 因而其生物地球化学过程相对于陆地生态系统更为复杂[3]。
土壤微生物是使土壤具有生命力的最主要成分, 与土壤肥力和土壤健康水平密切相关[4]。湿地生态系统中, 土壤微生物不仅在有机质分解、腐殖质合成以及物质转化和能量传递过程中发挥关键作用[5, 6], 还控制着沼泽湿地类型的分异与演替和土壤碳库的动态变化[7]。在洪泛平原湿地生态系统中, 土壤微生物群落结构对环境因子的变化尤其是土壤湿度、pH、质地结构和营养元素的可利用程度响应敏感[8]。水文情势的变化是湿地生态系统结构和功能的主要控制因素[9], 湿地水文状态的改变不仅制约着土壤微生物的生理和功能代谢水平, 还影响微生物功能群落的组成[10]。地下水位变化所引起的干湿交替环境通过其有氧和无氧环境的转变为土壤微生物的群落组成形成了生理压力[11]。微生物群落结构沿地下水位梯度的分异与土壤水分、透气性、有机养分和植被等因子的差异密切相关[12]。高地下水位梯度的湿润条件下, 细菌更具有生存优势[13]。
鄱阳湖是我国第一大淡水湖泊, 受流域来水和长江水位共同作用, 年内洪枯水位可达12m, 这种独特的水文节律变化下形成了大面积干湿交替的洲滩湿地[14]。沿不同的高程梯度, 地下水位变化呈明显的空间地带性分布, 进而在洲滩尺度上形成了不同的微地带性环境。相关研究表明, 水文情势的变化对鄱阳湖植被群落分布、生物量积累和物种多样性造成了不同程度的影响[15-16]。目前在鄱阳湖湿地的研究主要集中在水文过程与景观变化[17-18]、土壤微生物与植被关系[19-20]等方面, 这些研究对认识鄱阳湖湿地的变化规律奠定了基础。Coulis等[21]认为短期内生态环境因子的变化不会直接改变微生物的群落结构, 只能改变微生物活性, 枯水期内洲滩尺度上不同地下水位环境梯度是否会引起土壤微生物群落结构的变化及其与微环境因子的相关性研究尚不足。本文以鄱阳湖典型碟形子湖白沙湖洲滩湿地为研究对象, 研究比较不同地下水位梯度土壤湿地微生物生物量和群落结构的分异特征, 并对相关影响因素进行分析, 以期为深入了解枯水期鄱阳湖湿地生态系统结构稳定性的维持机制及湿地生物地球化学循环过程, 揭示土壤地下生物群系应对枯水期环境变化的响应策略提供科学依据, 进而对鄱阳湖湿地生态系统的有效保护具有重要意义。
1 材料与方法 1.1 研究区概况鄱阳湖地处江西北部, 长江中下游, 地理坐标为115°49′—116°46′E, 28°24′—29°46′N, 南北相隔170km, 东西最大跨度70km。鄱阳湖属东亚季风区, 为典型亚热带季风气候, 夏季炎热多雨, 冬季低温少雨, 年平均降水量为1387—1795mm, 年内、年际变化较大。年降水量最大2452.8mm(1954年), 最小1082.6mm(1978年), 最大6个月降水量(3—8月)占全年降水量的74.4%。年平均蒸发量800—1200mm, 约有一半集中在温度最高的7—9月。多年平均气温17.6℃, 最冷月(1月)平均气温5.1℃, 最热月(7月)平均气温29.5℃。受鄱阳湖水文节律季节性变化的影响, 呈现出河、湖、滩交错的湿地景观格局。
研究试验样地位于江西南矶湿地国家级自然保护区内的白沙湖, 地处赣江三角洲前缘, 属典型的内陆河口湿地区。白沙湖为典型的碟形湖泊, 洲滩地势平坦, 受鄱阳湖季节性周期水文变化影响, 4—9月份保护区除南山岛和矶山岛外, 其余地方均被淹没, 白沙湖与大湖连通, 10月份湖水逐渐消退, 归入碟形洼地, 不同高程的洲滩相继出露。白沙湖洲滩发育有大面积的草洲, 优势植被为灰化苔草(Carex cinerascens)、南荻(Triarrhena lutarioriparia)、芦苇(Phragmites australis)、虉草(Phalaris arundinacea)等, 其中灰化苔草分布最为广泛, 几乎遍及整个湿地洲滩, 盖度在85%—100%。试验样地位置如图 1所示, 面积为200m×300m, 样地内优势植被为灰化苔草, 伴生种为南荻和芦苇。从湖岸向湖心按照地下水位划分为4个梯度(GT-A, GT-B, GT-C, GT-D), 梯度带之间相隔约100m。不同梯度带灰化苔草生长态势、土壤温度及地下水位分异明显(表 1)。每个地下水位梯度带内根据微地形差异设置3—6个采样点, 同一梯度带内采样点间隔50—80m, 每个采样点设置3个重复。每个采样点处埋入细口径(内径为2cm)PVC管(埋入地下约70cm)用于测定地下水位。

|
| 图 1 研究区地理位置与样地设置 Fig. 1 Location of study area GT-A, GT-B, GT-C, GT-D代表不同的地下水位梯度带(从湖岸向湖心方向依次为A, B, C, D) |
| 梯度带 Gradient zones |
生物量 Phytomass */(g/m 2) |
平均高度 Mean height/cm |
平均盖度 Vegetation coverage/% |
土壤温度 Soil temperature/℃ |
平均地下水位 Mean water table/cm |
| GT-A | 153.33±67.39 | 40.25±7.70 | 51 | 25.33±0.58 | -(33.47±7.97) |
| GT-B | 171.46±88.49 | 29.55±2.36 | 63 | 25.80±1.30 | -(21.54±3.18) |
| GT-C | 70.05±14.13 | 27.48±3.70 | 42 | 24.67±0.82 | -(7.14±4.20) |
| GT-D | 50.87±19.19 | 12.88±9.02 | 17 | 24.33±0.52 | -(0.70±2.66) |
| *采样点生物量为苔草生物量, GT-A, GT-B, GT-C, GT-D代表不同的地下水位梯度带(从湖岸向湖心方向依次为A, B, C, D) | |||||
2016年11月下旬, 利用土钻(内径5cm)对每个采样点进行五点混合取样, 采集0—20cm土壤样品约500g装入聚乙烯自封袋内, 编号后置于保鲜盒运回实验室进行后续分析。土壤样品运回实验室后, 挑拣出石块等杂物后, 每份样品分为两份, 一份置于-20℃冷藏箱中用于测定土壤微生物磷脂脂肪酸(PLFA)测定, 另一部分自然风干后磨碎风干, 分别过20目和100目筛, 用于土壤理化性质的分析。
土壤理化性质分析具体方法如下:土壤含水量采用烘干法测定, 土壤颗粒组成采样马尔文激光粒度仪测定;土壤容重(BD)采用环刀法测定;土壤pH采用水土比2.5: 1 pH计测定;土壤有机碳(SOC)采用重铬酸钾外加热法测定;土壤总氮(TN)采用半微量凯氏定氮法测定;土壤总磷(TP)采用钼锑抗比色法测定。土壤微生物量碳(MBC)和微生物量氮(MBN)的测定采用氯仿熏蒸-K2SO4浸提法[22]。
土壤微生物磷脂脂肪酸(PLFA)的分析测定参照Frostegård等[23]的KOH-甲醇溶液甲酯化方法进行, 以十九烷酸为内标采用GC-MS (TRACE GC Ultra ISQ)测定, 全自动进样装置, 检测磷脂脂肪酸的方法为:进样口温度280℃, 分流比20: 1, 柱温箱程序升温为70℃起始, 保持1min, 以20℃/min升温至170℃, 维持2min, 再以5℃/min升温至280℃, 维持5min, 最后以40℃/min升温至3000℃, 维持1.5min。
对不同地下水位条件下的土壤进行PLFA分析, 共检测出30种不同的脂肪酸。通常情况下每类特征标记脂肪酸仅存在于特定的微生物类群内, 且占有优势地位[24]。因此不同的特征标记脂肪酸可以表征特定的生物类群。其中, i13:0, i14:0, i15:0, a15:0, i16:0, i17:0, a17:0, a16:0, i15:1可以表征革兰氏阳性细菌;12:0, 14:1ω5c, 14:0, 16:1ω6c, 15:0, cy17:0, 16:1 2OH, i17:0 3OH, cy19:0 ω8c, i15:0 3OH, 16:1ω9c可以表征革兰氏阴性细菌;16:1ω5c, 18:1ω9c, 18:3ω6c(6, 9, 12)表征真菌;10Me17:0和10Me 18:0表征放线菌[25-27]。
1.3 数据处理采用SPSS 20.0进行数据的统计分析, 采用单因素方差分析不同地下水位梯度环境下土壤理化性质、微生物群落结构的变化, 多重比较方法选用Dunnett′s T3比较检验, 采用双变量皮尔逊相关分析研究区微生物群落结构与土壤理化性质的相关性;采用CANOCO(Canoco for Windows 5.0)软件对不同地下水位环境梯度下不同采样点土壤微生物特征PLFAs进行主成分分析(PCA), 对土壤理化性质、土壤微生物群落结构之间的关系进行冗余分析(RDA), RDA分析通过蒙特卡洛显著性检验(Monte Carlo permutation text)(P=0.02)。
根据脂肪酸含量和种类计算微生物群落Shannon多样性指数(H′)、Pielou均匀度指数(J)和Simpson优势度指数(D)来评价微生物多样性特征[28-30], 计算公式如下:
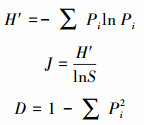
|
式中, Pi=Ni/N, Ni为第i类脂肪酸量, N为试验检测出的总脂肪酸量, S为微生物群落中总脂肪酸的种类。
2 结果与分析 2.1 土壤属性沿地下水位梯度的分异地下水位对TN以外的土壤理化性质影响显著(表 2;P < 0.05)。随着地下水位的升高, 土壤pH从4.62(GT-A)升至5.05(GT-D), 增长幅度达9.3%(P < 0.05);土壤含水量从30.8%(GT-A)增长到52.6%(GT-D), 变幅达70.7%(P < 0.05)。GT-D的粘粒含量比GT-A、GT-B和GT-C分别降低了34.6%、31.2%、22.7%, 粉粒含量分别降低了7.0%、4.6%和1.5%, 而沙粒含量则增加了4.28、2.18和1.28倍(P < 0.05)。地下水位的升高促进了土壤TOC和TP的流失, 相对GT-A, GT-D的TOC和TP含量显著降低了53.5%和26.1%(P < 0.05)。不同地下水位梯度间, 土壤TN差异性不显著(P > 0.05), 土壤碳氮比依次表现为GT-A(10.78)> GT-B(10.20)> GT-C(9.07)> GT-D(7.63)。
| 属性参数 Attribute parameter |
地下水位梯度带Ground water level gradient zones | |||
| GT-A | GT-B | GT-C | GT-D | |
| pH | 4.62±0.11a | 4.68±0.05a | 4.86±0.07b | 5.05±0.05c |
| 含水量Moisture/% | 30.80±1.61a | 31.36±2.82a | 33.93±6.24a | 52.56±10.20b |
| 沙粒Sand content/% | 3.44±1.36a | 6.77±1.68a | 11.53±2.92b | 14.74±3.45b |
| 粉粒Silt content/% | 78.74±0.88c | 76.70±1.49b | 74.33±1.38a | 73.19±1.41a |
| 粘粒Clay content/% | 17.30±1.12c | 16.45±0.74c | 14.65±1.84b | 11.32±0.79a |
| 容重Bulk density/(g/cm3) | 0.67±0.07c | 0.57±0.04c | 0.45±0.06b | 0.31±0.12a |
| 有机碳TOC/ (g/kg) | 10.32±3.12b | 9.73±1.85b | 6.19±1.23a | 4.80±0.60a |
| 总氮TN/(g/kg) | 0.97±0.08a | 0.98±0.19a | 0.66±0.03a | 0.58±0.17a |
| 总磷TP/(mg/kg) | 328.50±44.09b | 321.13±32.41b | 297.10±71.71b | 242.80±60.64a |
| 碳氮比C: N | 10.78±1.04b | 10.20±1.32b | 9.07±1.10ab | 7.63±1.38a |
| 不同小写字母表示同行在不同地下水位梯度带间在0.05水平上具有显著差异 | ||||
土壤生物量碳氮在不同地下水位梯度间表现为在GT-A、GT-B和GT-C之间差异不显著(图 2), GT-D显著高于其他3个梯度(P < 0.05)。GT-D的微生物量碳氮分别比GT-A增长了2.8倍和4.3倍。微生物商是指土壤微生物量碳和土壤有机碳含量的百分比(MBC/SOC)。随着地下水位梯度的升高, 微生物商逐渐升高, 在GT-A、GT-B和GT-C之间变化不显著, 相对GT-C, GT-D的微生物商升高了2.6倍。土壤微生物量氮的分配比例(MBN/TN)在不同地下水位梯度间的变化趋势与微生物商一致, GT-D土壤微生物量氮的分配比例是GT-C的3.95倍。

|
| 图 2 不同地下水位梯度土壤微生物MBC、MBN及其分配比例 Fig. 2 MBC, MBN, allocation ratio of MBC to SOC and allocation ration of MBN to TN of different ground water level gradient zones MBC:微生物量碳, Microbial biomass carbon;MBN:微生物量氮, Microbial biomass nitrogen;SOC:土壤有机碳, Soil organic carbon; TN:土壤总氮, Total nitrogen |
不同地下水位土壤微生物类群差异性显著(图 3)。GT-D显著增强了微生物总PLFAs, 细菌PLFA和放线菌PLFA的含量(P < 0.05), 与GT-A相比, 增长幅度分别为106.8%、117.2%、74.9%, 其中革兰氏阳性细菌和革兰氏阴性细菌分别增长了107.9%和207.2%。真菌PLFA随着地下水位的升高呈现出低-高-低-高的波动性变化。由GT-A到GT-D, 细菌PLFA占总PLFAs的比重分别为79.5%、80.9%、78.3%、83.5%, 真菌PLFA占总PLFAs的比重分别为12.9%、12.9%、11.3%、8.0%, 放线菌PLFA占总PLFA的比重分别为5.8%、6.2%、7.8%、4.9%。不同地下水位梯度样点土壤微生物PLFA数据PCA分析表明, 沿着第1、第2主排序轴(PC1), 采样点产生了明显的空间分离, GT-D和GT-A、GT-B、GT-C沿第1主排序轴能明显分开;而GT-A、GT-B、GTC之间则沿第2主排序轴明显分开(图 4)。PC1和PC2分别解释了空间分异的76.7%和13.6%, 累积贡献率达90.3%。在所有特征PLFA中, 对第1主成分起主要作用的是i13:0、i15:0、i17:0 3OH、i15:0 3OH、a16:0;而18:3ω6c、10Me 17:0对第2主成分贡献较大。

|
| 图 3 不同地下水位梯度土壤的总PLFA, 细菌, 真菌, 放线菌, 革兰氏阳性细菌, 革兰氏阴性细菌 Fig. 3 The amount of soil PLFAs (total PLFAs, total bacteria, gram-positive bacteria, gram-negative bacteria, Actinomycetes and Fungi of different ground water level gradient zones 不同小写字母表示在0.05水平上具有显著差异 |

|
| 图 4 PLFA指纹图谱主成分分析(A)土壤微生物群落, (B)磷脂脂肪酸 Fig. 4 Principle component analysis of PLFA profiles from soil microbial communities, (A)soil microbial community structure, (B) phospholipid fatty acids contributing to soil microbial community ordination pattern |
在地下水位梯度升高的过程中, 真菌/细菌(F/B)由0.26(GT-A)降至0.11(GT-D), 压力响应指数即支链反异构饱和脂肪酸与支链异构饱和脂肪酸的比值(SA/SI)由0.47(GT-A)升至0.79(GT-D)。4个地下水位梯度带中, GT-D均具有最低的革兰氏阳性细菌/革兰氏阴性细菌(G(+)/G(-))和单一不饱和脂肪酸/支链脂肪酸(M/B)(表 3;P < 0.05)。
| 梯度带 Gradient zones | G(+)/G(-) | F/B | M/B | SA/SI |
| GT-A | 2.12±0.22b | 0.17±0.02b | 0.26±0.02b | 0.47±0.05a |
| GT-B | 2.06±0.26b | 0.16±0.02b | 0.30±0.06b | 0.53±0.08a |
| GT-C | 2.36±0.69b | 0.14±0.03b | 0.18±0.06a | 0.53±0.08a |
| GT-D | 1.45±0.11a | 0.10±0.02a | 0.11±0.02a | 0.79±0.11b |
| 不同小写字母表示同列不同行在0.05水平上具有显著差异; F/B:真菌与细菌的比值, the ratio of fungi to bacterial; M/B:单一不饱和脂肪酸与支链脂肪酸的比值, the ratio of monounsaturated fatty acid to branched-chain fatty acid;SA/SI:支链反异构饱和脂肪酸与支链异构饱和脂肪酸的比值, the ratio of branched anti-isomeric saturated fatty acids to branched chain isomeric saturated fatty acids | ||||
不同地下水位梯度湿地土壤微生物群落的多样性分析表明, Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J)的最低值出现在GT-D, 而最高值出现在GT-B。Simpson优势度指数(D)则与之相反(图 5;P < 0.05), 表明地下水位梯度的升高降低了土壤微生物结构多样性, 而促进了优势种群的分布。

|
| 图 5 不同地下水位梯度土壤微生物群落Shannon-Wiener多样性指数(H′), Pielou均匀度指数(J)和Simpson优势度指数(D) Fig. 5 Shannon-Wiener diversity index, Pielou evenness index and Simpson dominance index of soil microbial communities under different ground water level gradient zones 不同小写字母表示在0.05水平上具有显著差异 |
相关分析表明(表 4), 湿地土壤化学性质中, pH与G(+)、G(-)、细菌、放线菌、总PLFAs和优势度指数呈极显著正相关(P<0.01), 而与多样性指数和优势度指数呈极显著负相关(P<0.01)。TOC与多样性指数和均匀度指数呈极显著正相关(P<0.01)而与优势度指数呈极显著负相关(P<0.01)。C: N与G(-)、放线菌和优势度指数呈极显著负相关(P<0.01), 而与多样性指数和均匀度指数呈极显著正相关(P<0.01)。湿地土壤物理结构中, 容重粘粒和粉粒与微生物群落结构表现出相同的关系, 即与真菌以外的微生物群落和优势度指数呈极显著负相关(P<0.01), 而与多样性指数和均匀度指数呈极显著正相关(P<0.01), 土壤沙粒含量则表现出相反的趋势(P<0.01)。MBC与G(+)、G(-)、细菌、放线菌和总PLFA均呈极显著正相关(P<0.01), 与优势度指数呈显著正相关(P<0.05), 而与多样性指数呈极显著负相关(P<0.01), MBN则与G(+)、G(-)、细菌、总PLFA和优势度指数呈极显著正相关(P<0.01), 与放线菌呈显著正相关(P<0.05), 与多样性指数呈极显著负相关(P<0.01)。
| 属性参数 Attribute parameter |
G (+) | G (-) | 细菌 Bacteria |
真菌 Fungi |
放线菌 Actinomycetes |
总PLFA Total PLFA |
H′ | J | D |
| pH | 0.74** | 0.72** | 0.69** | -0.01 | 0.69** | 0.68** | -0.74** | -0.85** | 0.73** |
| 含水量Moisture | 0.79** | 0.77** | 0.84** | 0.43 | 0.24 | 0.83** | -0.42 | -0.48* | 0.41 |
| 容重Bulk density | -0.78 | -0.74** | -0.71** | -0.38 | -0.65** | -0.73** | 0.73** | 0.80** | -0.73** |
| 粘粒Clay | -0.76** | -0.76** | -0.68** | -0.13 | -0.61** | -0.68** | 0.77** | 0.84** | -0.77** |
| 粉粒Silt | -0.55* | -0.51* | -0.47* | 0.02 | -0.64** | -0.47* | 0.61** | 0.70** | -0.61** |
| 沙粒Sand | 0.57** | 0.56* | 0.52* | 0.11 | 0.63** | 0.53* | -0.61** | -0.70** | 0.60** |
| SOC | -0.39 | -0.43 | -0.33 | 0.15 | -0.49* | -0.34 | 0.60** | 0.69** | -0.59** |
| TN | -0.19 | -0.25 | -0.14 | 0.19 | -0.30 | -0.14 | 0.44 | 0.51* | -0.43 |
| C: N | -0.56* | -0.57** | -0.50* | -0.08 | -0.59** | -0.50* | 0.58** | 0.66** | -0.58** |
| TP | -0.34 | -0.39 | -0.34 | 0.17 | -0.21 | -0.33 | 0.40 | 0.47* | -0.39 |
| MBC | 0.84** | 0.87** | 0.86** | 0.27 | 0.67** | 0.85** | -0.55* | -0.66** | 0.55* |
| MBN | 0.80** | 0.85** | 0.81** | 0.25 | 0.54* | 0.80** | -0.62** | -0.69** | 0.61** |
| **表示在0.01水平上具有极显著相关性;*表示在0.05水平上具有显著相关性 | |||||||||
土壤微生物商和土壤环境因子间具有显著的线性关系(图 6), MBC/SOC在研究区范围内随着pH和土壤含水量的增大而增大, 随着容重和粘粒含量的增大而减小。但是与pH、容重和粘粒含量相比, MBC/SOC与含水量间的回归拟合效果明显较差(R2=0.20, P=0.03)。MBC/SOC对pH变化的响应最敏感(R2=0.72, P < 0.001), 其次为粘粒含量。

|
| 图 6 土壤微生物商与土壤环境因子的关系 Fig. 6 Relationship between microbial quotient and soil environmental factors |
对所有地下水位带湿地土壤微生物群落结构进行冗余分析发现, 前2个排序轴累积贡献值为94.8%, 其中第1排序轴解释了82.9%的变异(图 7)。在所有土壤环境因子中, Monte Carlo检验表明, 土壤微生物群落结构与pH(F=17.3, P=0.002), 土壤含水量(F=3.8, P=0.04), 和C: N(F=2.7, P=0.05)显著相关。pH和土壤含水量与第1轴呈显著正相关, 而C: N与第1轴呈显著负相关。通过单个PLFA与环境因子相关性排序可知, 革兰氏阴性细菌PLFA标记(i15:0 3OH、i17:0 3OH、15:1ω6c、15:0、cy17:0)和革兰氏阳性细菌PLFA标记(i13:0、i14:0、a16:0)与土壤含水量呈显著正相关, 即这些PLFAs的相对丰度值随着土壤含水量的升高而升高。真菌特征PLFA(18:3ω6c和18:1ω9c)与C: N呈显著正相关。革兰氏阳性细菌PLFA标记(i15:0、14:0)、革兰氏阴性细菌PLFA标记(14:1ω5c)和真菌PLFA标记(16:1ω5c)与pH呈显著正相关。革兰氏阳性细菌PLFA标记(a17:0)、革兰氏阴性细菌PLFA标记(12:0)与沙粒含量呈显著正相关。放线菌PLFA标记(10Me18:0、10Me16:0)与各土壤环境因子相关性均不显著。

|
| 图 7 土壤微生物优势群落与环境因子的RDA排序 Fig. 7 Redundancy analysis (RDA) of the microbial distribution patterns constrained by soil physicochemical properties along ground water level gradients |
水文条件是湿地形成和发育的驱动因子。当水淹没洲滩土壤时, 进入土壤的氧气减少, 起分解作用的好氧微生物活动减弱, 有机质分解速率低, 土壤有机碳以积累为主[31]。而在湿地土壤干湿交替地条件下, 会引起土壤收缩和膨胀, 使土壤团聚体中有机质暴露, 对土壤微生物代谢具有激发效应, 促进有机质的分解[32]。本研究发现不同地下水位梯度间土壤理化性质差异明显, 湿地土壤机械组成的分异主要是因为水体波动过程中的冲刷和筛选作用导致细粒物质被水流带走而使得比重较大的粗颗粒物积累下来[33]。此外, 地下水位波动引起的干湿交替过程会引起土壤胶体的形态变化, 破坏土壤团聚体结构。土壤pH, 颗粒组成和容重的变化使原来不能被分解的有机质因团聚体的崩溃而加速分解, 所以土壤有机碳的含量显著降低[34]。张全军等[35]研究发现高盖度、长势旺盛的苔草群落有利于养分保留和有机碳的固持, 由于靠近水体的梯度带苔草盖度、株高、生物量等生物性状显著低于其他梯度, 故而不利于土壤有机碳的积累。土壤全磷含量主要受成土母质、气候环境条件影响, 受植物群落影响不大[36], 高地下水位梯度土壤磷含量显著降低主要是因为可溶性磷酸盐被淋溶分解[37], 而研究表明可溶性有机碳中的芳香类物质能够保护土壤粘粒抵抗外界的侵蚀[38], 进而又促进了粘粒含量的减少。
土壤微生物量可以反映土壤同化和矿化能力的大小, 是土壤活性大小的标志[21]。本研究中靠近水体的高地下水位梯度具有最高的生物量碳氮, 可能是因为pH和土壤含水量综合作用的结果。枯水季节期间, 洲滩土壤含水量在30%—55%之间, 土壤含水量的升高有利于微生物活性的增强。由于枯水期洲滩土壤pH在4.5—5.5之间, 高地下水位梯度pH和微生物量碳氮均高于其他梯度, 说明研究区土壤微生物更喜欢弱酸性的环境。
湿地土壤地下水位改变了土壤微生物群落组成。随着地下水位梯度空间位置的升高, PLFA总量、细菌群落、放线菌群落的PLFA含量显著增加, 最高值均出现在地下水位梯度最高的GT-D。Mentzer等研究发现湿地水文情势的变化能够促进土壤微生物群落结构的持久性变化[39]。湿地水文波动是影响土壤氧气浓度的驱动型因素, 但是在枯水季节, 好氧环境和厌氧环境会根据降水或湖泊水位变化同时出现在高程梯度较低的区域[40], 因此在高程低, 靠近水体的地带, 水位会在地面表层上下波动, 进而导致干湿交替过程。本研究中的结果表明高地下水位区域频繁的干湿交替能够促进细菌和放线菌群落的丰度, 与Foulquier等的研究结果一致[41]。真菌群落对酸性的耐受程度高于细菌[42], 因此本研究中真菌群落并没有随pH产生明显变化。
由于土壤微生物量碳氮的周转快, 所以它们的分配比例越大, 土壤有机碳等的周转速率就越大[43]。靠近水体高地下水位梯度带土壤微生物商和微生物量氮分配比例最高, 说明土壤有机碳氮周转最快, 这也解释了土壤有机碳在此梯度带含量低的原因。微生物商与土壤含水量之间的相关性远远弱于pH、容重和粘粒含量。研究表明, 短时期的淹水对土壤有机碳的周转速率影响不明显[43], 说明地下水位梯度对微生物商的影响主要是通过土壤pH和土壤质地的作用。
相关分析和RDA分析都表明, 在地下水位梯度空间分异过程中, pH、土壤含水量和土壤机械组成是影响土壤微生物群落结构的主要环境因素。因为土壤pH会对微生物吸附、胞外酶的分泌以及养分利用产生不同的作用, 故而其对微生物群落结构的影响机制比较复杂。一般来说, 细菌和放线菌适合生长在微碱性或弱酸性环境下, 而真菌则适合生长在酸性条件下[44]。因此细菌和放线菌的PLFA含量与土壤pH呈极显著正相关, 而随着pH的升高, 真菌所占的比重逐渐降低。土壤机械组成对微生物群落结构的直接影响主要是因为土壤微生物主要存在于土壤颗粒的空隙或表面。研究发现, 细菌主要存在于0.5—5μm的空隙中, 而放线菌也适合生存在相对较大的空隙中[45], 土壤容重随着土壤孔隙度的增大而减小, 而本研究中细菌和放线菌的含量与土壤容重具有显著的负相关性, 可能是因为在湿地环境中土壤颗粒空隙的增大有利于细菌和放线菌的存活。Cook等发现, 土壤微生物群落中细菌在高水位条件下活性高而真菌则在低水位条件下活性较高[46]。无论是在陆地生态系统还是在湿地生态系统土壤水分都是土壤微生物群落结构的重要控制因素。湿地土壤地下水位的抬升, 最直接的影响就是使土壤含水量升高。土壤碳氮比可以反映微生物生长是碳限制还是氮限制, 同时也是影响土壤微生物群落结构的重要因子。碳氮比越小说明碳的可利用程度越低, 由于真菌需要有机残体作为生命活动的载体, 与外源有机碳输入和土壤有机质积累关系密切[47], 所以本研究中细菌和放线菌群落对碳限制所带来的环境压力耐受性显著高于真菌群落, 有机碳可利用程度的降低所带来的负面效应并没有影响pH升高所带来的正面效应。在湿地土壤环境下, 地下水位的空间变化不仅关系到土壤水分条件和氧气浓度的变化, 而且还反映了土壤结构和养分的可利用性, 因此对土壤微生物群落结构影响复杂。土壤微生物群落的多样性指数和均匀度指数均随着地下水位梯度升高而降低, 说明地下水位梯度所综合表征的土壤环境梯度的变化对土壤微生物具有明显的筛选作用, 不同的湿地微生态环境塑造了不同的微生物群落, 这种微生物群落结构的变化可能是湿地枯水季节地下水位短期分异的结果[21]。
真菌/细菌(F/B)常被用来衡量微生物群落对环境变化的响应程度, 进而能够反映土壤微生物的生理状态, 比如有机质的分解[47]。与其他地下水位梯度带相比, GT-D具有最低的F/B, 说明高地下水位梯度碳的积累效率较慢[48], 与微生物商所表现的结果一致。Gordon等[49]研究发现在干湿交替环境中F/B并未发生明显变化, 间接指示pH可能是影响F/B变化的主要环境因子。单一不饱和脂肪酸/支链脂肪酸(M/B)可以用来指示厌氧与好氧微生物的比例[50]。本研究中GT-C和GT-D的M/B显著低于GT-A和GT-B, 说明随着地下水位的升高, 厌氧微生物的比重在逐渐增多, 而好氧微生物比重的增长速度低于厌氧微生物, 这与土壤含水量的变化是一致的。支链反异构饱和脂肪酸/支链异构饱和脂肪酸(SA/SI)能够反映微生物群落环境压力的变化, 一般随着温度的降低和有机碳可利用程度的增加而升高[51-52]。由于pH的升高和土壤颗粒的变化促进了土壤有机质的分解和微生物的代谢强度, 并且随着地下水位的升高, 土壤温度降低, 所以本研究中SA/SI随地下水位呈显著增大趋势。
鄱阳湖湿地不同地下水位环境梯度间土壤微生物群落结构存在较大差异, 由于本研究实验样地区域为单一灰化苔草群落, 所以土壤微生物群落结构的分异主要是由于地下水位变化及其所驱动的土壤理化性质的改变所引起的。本研究则发现, 在枯水期湿地洲滩出露阶段, 鄱阳湖湿地短时期内所表现出的不同的地下水位环境梯度空间异质性既改变了微生物活性又同时显著改变了土壤微生物群落结构, 进而有可能对有机质分解、土壤呼吸等湿地生态过程产生不同影响, 同时靠近水体、地下水位相对较高的环境梯度带土壤碳氮周转速率快, 是湿地生物地球化学循环的热点区域。
4 结论(1) 地下水位是湿地洲滩微地形、高程和水文情势综合作用的结果。地下水位的升高显著改变了土壤化学性质和物理结构。随着地下水位梯度带从最低到最高, 土壤pH从4.62升至5.05, 沙粒含量增加了4.28倍, 而有机碳、粘粒和粉粒含量分别降低了53.5%、34.6%和7.0%。
(2) 湿地地下水位改变驱动下土壤环境因子的变化进一步改变了湿地土壤微生物量碳氮和土壤碳氮周转速率。与地下水位最低的梯度带(GT-A)相比, 当平均地下水位在地表上下波动时(GT-D), 微生物量碳氮分别增长了2.82和4.30倍, 微生物商提高了5.77倍, 微生物量氮分配比例提高了7.15倍。
(3) 湿地地下水位梯度驱动下湿地微环境的变化短期内改变了土壤微生物群落结构, 与地下水位最低的梯度(GT-A)相比, 干湿交替环境(GT-D)下, 微生物总PLFAs, 细菌PLFA和放线菌PLFA的含量分别增长了106.79%、117.24%、74.89%, 随着地下水位升高, 真菌PLFA比重逐渐降低而厌氧微生物比重增多。
(4) 湿地地下水位梯度的升高增加了土壤微生物群落的环境压力, 进而降低了土壤微生物群落多样性。高地下水位梯度(GT-D)具有最低的F/B(0.11)和最高的SA/SI(0.79)。不同地下水位环境梯度土壤微生物的多样性指数和均匀度指数表现为GT(A)> GT(B)> GT(C)> GT(D), 而优势度指数则与之相反。
(5) 在湿地地下水位变化过程中, 影响土壤有机碳周转的主要土壤环境因子为pH和土壤质地, 而影响土壤微生物群落结构的关键限制性因子为pH, 土壤含水量, 沙粒含量和C: N。
| [1] | 陈宜瑜, 吕宪国. 湿地功能与湿地科学的研究方向. 湿地科学, 2003, 1(1): 7–11. |
| [2] | Mitsch W J, Gosselink J G. Wetlands. New Jesey:John Wiley and Sons, 2007. |
| [3] | Kayranli B, Scholz M, Mustafa A, Hedmark Å. Carbon storage and fluxes within freshwater wetlands:a critical review. Wetlands, 2010, 30(1): 111–124. DOI:10.1007/s13157-009-0003-4 |
| [4] | 林先贵. 土壤微生物研究原理与方法. 北京: 高等教育出版社, 2010: 1–2. |
| [5] | Joergensen R G, Scheu S. Response of soil microorganisms to the addition of carbon, nitrogen and phosphorus in a forest Rendzina. Soil Biology and Biochemistry, 1999, 31: 859–866. |
| [6] | Harris J A. Measurements of the soil microbial community for estimating the success of restoration. European Journal of Soil Science, 2003, 54(4): 801–808. DOI:10.1046/j.1351-0754.2003.0559.x |
| [7] | 肖烨, 黄志刚, 武海涛, 吕宪国. 三江平原典型湿地类型土壤微生物特征与土壤养分的研究. 环境科学, 2015, 36(5): 1842–1848. |
| [8] | Poret-Peterson A T, Ji B M, Engelhaupt E, Gulledge J. Soil microbial biomass along a hydrologic gradient in a subsiding coastal bottomland forest:implications for future subsidence and sea-level rise. Soil Biology and Biochemistry, 2007, 39(2): 641–645. DOI:10.1016/j.soilbio.2006.09.016 |
| [9] | 杨桂生, 宋长春, 王丽, 王丽丽, 李英臣. 水位梯度对小叶章湿地土壤微生物活性的影响. 环境科学, 2010, 31(2): 444–449. |
| [10] | Gerull L, Frossard A, Gessner M O, Mutz M. Variability of heterotrophic metabolism in small stream corridors of an early successional watershed. Journal of Geophysical Research, 2011, 116(G2). |
| [11] | Schulze E D, Freibauer A. Carbon unlocked from soils. Nature, 2005, 437(7056): 205–206. DOI:10.1038/437205a |
| [12] | 李金前, 王吉, 刘亚军, 邹锋, 马燕天, 吴兰. 水位高程变化对湿地土壤微生物代谢功能的影响研究——以蚌湖为例. 长江流域资源与环境, 2017, 26(5): 730–737. |
| [13] | van Dijk J, Didden W A. M, Kuenen F, van Bodegom P M, Verhoef H A, Aerts R. Can differences in soil community composition after peat meadow restoration lead to different decomposition and mineralization rates? Soil Biology and Biochemistry, 2009, 41(8): 1717–1725. |
| [14] | 赵其国, 黄国勤, 钱海燕. 鄱阳湖生态环境与可持续发展. 土壤学报, 2007, 44(2): 318–326. DOI:10.11766/trxb200512310219 |
| [15] | Dai X, Wan R R, Yang G S, Wang X L, Xu L G. Responses of wetland vegetation in Poyang Lake, China to water-level fluctuations. Hydrobiologia, 2016, 773(1): 35–47. DOI:10.1007/s10750-016-2677-y |
| [16] | 冯 文娟, 徐 力刚, 王 晓龙, 李 海英, 姜 加虎. 鄱阳湖洲滩湿地地下水位对灰化薹草种群的影响. 生态学报, 2016, 36(16): 5109–5115. |
| [17] | 查东平, 冯明雷, 陈宏文, 刘足根, 廖兵, 申展. 鄱阳湖典型湿地植被景观格局的时空变化分析. 水生态学杂志, 2015, 36(5): 1–7. |
| [18] | 方朝阳, 邬浩, 陶长华, 高丹, 周华. 鄱阳湖南矶湿地景观信息高分辨率遥感提取. 地球信息科学, 2016, 18(6): 847–856. |
| [19] | 王晓龙, 徐立刚, 姚鑫, 白丽, 张奇. 鄱阳湖典型湿地植物群落土壤微生物量特征. 生态学报, 2010, 30(18): 5033–5042. |
| [20] | Wang X L, Xu L G, Wan R R, Chen Y W. Seasonal variations of soil microbial biomass within two typical wetland areas along the vegetation gradient of Poyang Lake, China. Catena, 2016, 137: 483–493. DOI:10.1016/j.catena.2015.10.020 |
| [21] | Coulis M, Fromin N, David J F, Gavinet J, Clet A, Devidal S, Roy J, Hättenschwiler S. Functional dissimilarity across trophic levels as a driver of soil processes in a Mediterranean decomposer system exposed to two moisture levels. Oikos, 2015, 124(10): 1304–1316. DOI:10.1111/oik.2015.v124.i10 |
| [22] | 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000: 12–235. |
| [23] | Frostegård A, Bååth E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biology and Fertility of Soils, 1996, 22(1/2): 59–65. |
| [24] | 颜慧, 蔡祖聪, 钟文辉. 磷脂脂肪酸分析方法及其在土壤微生物多样性研究中的应用. 土壤学报, 2006, 43(5): 851–859. |
| [25] | Sun Y X, Wu J P, Shao Y H, Zhou L X, Mai B X, Lin Y B, Fu S. Responses of soil microbial communities to prescribed burning in two paired vegetation sites in southern China. Ecological Research, 2011, 26(3): 669–677. DOI:10.1007/s11284-011-0827-2 |
| [26] | Wang X L, Wang X L, Zhang W X, Shao Y H, Zou X M, Liu T, Zhou L X, Wan S Z, Rao X Q, Li Z A, Fu S L. Invariant community structure of soil bacteria in subtropical coniferous and broadleaved forests. Scientific Reports, 2016, 6: 19071. DOI:10.1038/srep19071 |
| [27] | Bossio D A, Scow K M. Impacts of carbon and flooding on soil microbial communities:phospholipid fatty acid profiles and substrate utilization patterns. Microbial Ecology, 1998, 35(3/4): 265–278. |
| [28] | Magurran A E. Ecological diversity and Its Measurement. Princeton, NJ:Princeton University Press, 1988: 141–162. |
| [29] | Pielou E C. Species-diversity and pattern-diversity in the study of ecological succession. Journal of Theoretical Biology, 1966, 10(2): 370–383. DOI:10.1016/0022-5193(66)90133-0 |
| [30] | Hunter P R, Gaston M A. Numerical index of the discriminatory ability of typing systems:an application of Simpson's index of diversity. Journal of Clinical Microbiology, 1988, 26(11): 2465–2466. |
| [31] | Sahrawat K L. Organic matter accumulation in submerged soils. Advances in Agronomy, 2003, 81: 169–201. DOI:10.1016/S0065-2113(03)81004-0 |
| [32] | 孟 伟庆, 莫 训强, 胡 蓓蓓, 贺 梦璇, 李 洪远. 模拟干湿交替对湿地土壤呼吸及有机碳含量的影响. 土壤通报, 2015, 46(4): 910–915. |
| [33] | Lyu X F, Yu J B, Zhou M, Ma B, Wang G M, Zhan C, Han G X, Guan B, Wu H F, Li Y Z, Wang D. Changes of soil particle size distribution in tidal flats in the Yellow river delta. PLoS One, 2015, 10(3). |
| [34] | 黄昌勇. 土壤学. 北京: 中国农业出版社, 2000. |
| [35] | 张全军, 于秀波, 钱建鑫, 熊挺. 鄱阳湖南矶湿地优势植物群落及土壤有机质和营养元素分布特征. 生态学报, 2012, 32(12): 3656–3669. |
| [36] | 白军红, 丁秋祎, 高海峰, 王庆改, 肖蓉. 向海湿地不同植被群落下土壤氮素的分布特征. 地理科学, 2009, 29(3): 381–384. |
| [37] | Callaway J C, Borgnis E L, Turner R E, Milan C S. Carbon sequestration and sediment accretion in San Francisco Bay tidal wetlands. Estuaries and Coasts, 2012, 35(5): 1163–1181. DOI:10.1007/s12237-012-9508-9 |
| [38] | Mavi M S, Marschner P, Chittleborough D J, Cox J W, Sanderman J. Salinity and sodicity affect soil respiration and dissolved organic matter dynamics differentially in soils varying in texture. Soil Biology and Biochemistry, 2012, 45: 8–13. DOI:10.1016/j.soilbio.2011.10.003 |
| [39] | Mentzer J L, Goodman R M, Balser T C. Microbial response over time to hydrologic and fertilization treatments in a simulated wet prairie. Plant and Soil, 2006, 284(1/2): 85–100. |
| [40] | Davidsson T E, Stepanauskas R, Leonardson L. Vertical patterns of nitrogen transformations during infiltration in two wetland soils. Applied and Environmental Microbiology, 1997, 63(9): 3648–3656. |
| [41] | Foulquier A, Volat B, Neyra, M, Bornette G, Montuelle B. Long-term impact of hydrological regime on structure and functions of microbial communities in riverine wetland sediments. FEMS Microbiology Ecology, 2013, 85(2): 211–226. DOI:10.1111/femsec.2013.85.issue-2 |
| [42] | Rousk J, Brookes P C, Bååth E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Applied and Environmental Microbiology, 2009, 75(6): 1589–1596. DOI:10.1128/AEM.02775-08 |
| [43] | 柴雪思, 雷利国, 江长胜, 黄哲, 范志伟, 郝庆菊. 三峡库区典型消落带土壤微生物生物量碳、氮的变化特征及其影响因素探讨. 环境科学, 2016, 37(8): 2979–2988. |
| [44] | 胡亚林, 汪思龙, 颜绍馗. 影响土壤微生物活性与群落结构因素研究进展. 土壤通报, 2006, 37(1): 170–176. |
| [45] | Foster R C. Microenvironments of soil microorganisms. Biology and Fertility of Soils, 1988, 6(3): 189–203. |
| [46] | Cook R J, Papendick R I. Soil water potential as a factor in the ecology of Fusarium roseum f. sp. cerealis 'Culmorum'. Plant and Soil, 1970, 32(1/3): 131–145. |
| [47] | Huang Y M, Liu D, An S S. Effects of slope aspect on soil nitrogen and microbial properties in the Chinese Loess region. CATENA, 2015, 125: 135–145. DOI:10.1016/j.catena.2014.09.010 |
| [48] | Joergensen R G, Wichern F. Quantitative assessment of the fungal contribution to microbial tissue in soil. Soil Biology and Biochemistry, 2008, 40(12): 2977–2991. DOI:10.1016/j.soilbio.2008.08.017 |
| [49] | Gordon H, Haygarth P M, Bardgett R D. Drying and rewetting effects on soil microbial community composition and nutrient leaching. Soil Biology and Biochemistry, 2008, 40(2): 302–311. DOI:10.1016/j.soilbio.2007.08.008 |
| [50] | Cao Y S, Fu S L, Zou X M, Cao H L, Shao Y H, Zhou L X. Soil microbial community composition under eucalyptus plantations of different age in subtropical China. European Journal of Soil Biology, 2010, 46(2): 128–135. DOI:10.1016/j.ejsobi.2009.12.006 |
| [51] | Bach E M, Baer S G, Meyer C K, Six J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biology and Biochemistry, 2010, 42(12): 2182–2191. DOI:10.1016/j.soilbio.2010.08.014 |
| [52] | Petersen S O, Klug M J. Effects of sieving, storage, and incubation temperature on the phospholipid fatty acid profile of a soil microbial community. Applied and Environmental Microbiology, 1994, 60(7): 2421–2430. |