 2018, Vol. 38
2018, Vol. 38文章信息
- 白耀宇, 庞帅, 韦珊, 肖瑶, 丁伟.
- BAI Yaoyu, PANG Shuai, WEI Shan, XIAO Yao, DING Wei.
- 烟草青枯病危害对烟田中小型土壤动物群落的影响
- Effects of tobacco bacterial wilt on meso-micro soil fauna in tobacco fields of Chongqing
- 生态学报. 2018, 38(11): 3792-3805
- Acta Ecologica Sinica. 2018, 38(11): 3792-3805
- http://dx.doi.org/10.5846/stxb201705090853
-
文章历史
- 收稿日期: 2017-05-09
- 网络出版日期: 2018-03-02

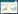




烟草青枯病是由茄科雷尔氏菌(Ralstonia solanacearum)引起的一种毁灭性土传细菌性病害, 严重制约世界上主要产烟国家的烟草生产[1]。该病在我国南方烟区普遍发生, 特别在湖南、贵州、福建、广东、四川、重庆等省发生较为严重[2]。由于该病原菌主要在土壤中完成越冬、侵染和传播, 导致其很难从根本上有效控制[3]。
据报道, 该病害的病原菌在土壤中传播和侵染寄主植物根系的途径也与土壤动物的发生及危害密切相关[1]。而作为土壤生态系中重要组成部分的体长小于2 mm的小型(Micro-)和中型(Meso-)土壤动物, 如线虫(Nematoda)及节肢动物中的蜱螨(Acari)、弹尾虫(Collembola)和昆虫(Insecta)等[4], 具数量大、种类多和对环境变化敏感等特点, 在可持续农业生产中能有效指示土壤的健康程度[5-6]。但目前在烟草青枯病发生机理和控制技术研究中, 尤其突出该病与土壤微生物的互作规律研究[7-8];却忽视了与土壤动物的相关性研究。前人研究认为, 几乎所有的土壤过程和土壤功能都与土壤动物有关[9];只有土壤动物的积极参与, 植物根系、根系分泌物和土壤微生物间的相互作用才能顺利进行[9-10];土壤动物在调节根际微生物过程中担负着重要的生态功能, 从而显著影响植物的生长[11-12]。其中, 中小型土壤动物的生态功能主要通过直接取食真菌、细菌和植物凋落物碎屑和间接作用土壤理化性质及土壤微生物群落结构组成和功能特征来实现[13];也能在抑制植物土传病原菌致病危害方面发挥重要作用[14-16]。总之, 研究青枯病与中小型土壤动物的生态关系是揭示其爆发危害和寻找有效防治手段的重要理论基础。因此有必要开展烟草青枯病危害与该类土壤动物群落生态关系的研究。
迄今, 研究烟草青枯病对土壤动物影响的报道很少。我们前期在青枯病危害烟田的研究指出, 螨类数量在该病害发病的中后期显著下降, 而弹尾虫的数量却显著上升;螨类与弹尾虫的数量比值(A/C值)也随着青枯病危害程度的加剧而显著降低[17]。尽管如此, 在田间群落水平上, 青枯病危害对烟田中小型土壤动物的影响如何, 不得而知。基于此, 本研究以重庆市彭水县植烟区青枯病典型发病田及其对照无青枯病田为代表样点, 比较分析该病害发生对两类烟田中小型土壤动物群落结构及多样性功能特征的影响及规律, 并解析中小型土壤动物群落对青枯病危害响应的主要特征指标;旨在丰富和完善调控烟草青枯病作用的土壤生物基础理论, 促进形成可持续和有效的烟草青枯病生物防治理论和技术体系。
1 研究区概况研究区位于重庆市彭水县润溪乡(29°2′—29°13′ N, 107°52′—108°1′ E)。该乡地处渝贵交界处, 在彭水县城西南方向约53 km处, 属中亚热带温润季风气候, 雨量充沛, 四季分明, 山地立体气候特征十分明显, 海拔落差大, 最高处1498 m, 平均海拔841 m;以烟叶种植为特色;烟草业产值约占全乡农业总收入的三分之一。该县年均温17.6 ℃, 高山与低谷气温相差10—14 ℃, 年均降雨量1224 mm。
烟草青枯病典型发病区位于该乡白果坪村。该片植烟区近10年均连年受到烟草青枯病的重大危害, 部分田块在烟草成熟期青枯病发病株率甚至达到了100%[2, 17]。因此, 彭水县烟草分公司专门开辟了13.3 hm2青枯病危害严重的烟田作为该病害的研究与综合治理示范区[2]。本实验选取的烟草青枯病实验田就位于该示范区中。同时在该发病田块附近的润溪乡樱桃井村植烟区选取无青枯病危害史的烟田作为对照田(青枯病发病株率为0)。
2 材料与方法 2.1 样地设置及土壤理化特性测定在青枯病发病区和对照样区各选取3个面积为30 m×20 m的烟田样地为实验地, 每块实验地之间通过1.0 m的垄作非耕作区进行间隔。烟田育秧育苗、移栽和肥水管理等均按照当地农民种植习惯进行。试验田均不进行病虫害的化学药剂防治。
于2015年3月样品采集后, 将土壤动物分离后的0—15 cm样品混合, 经风干、除杂、研碎和混匀并过2 mm尼龙筛后, 用于测定土壤的理化性质。测定分析按鲍士旦主编的《土壤农化分析》[18]中的方法进行。经分析供试土壤为黄壤粘土。其他测定结果见表 1。
| 指标 Indexes |
青枯病发病田 Hazard fields of tobacco wilt |
无青枯病田(CK) Control fields without hazard |
| pH值pH value | 4.97 | 4.93 |
| 有机质Organic matter content/(g/kg) | 17.97 | 35.21 |
| 全氮Total nitrogen/(g/kg) | 0.31 | 1.64 |
| 全磷Total phosphorus/(g/kg) | 0.52 | 1.36 |
| 全钾Total potassium/(g/kg) | 13.61 | 15.30 |
| 容重Soil bulk density/(g/cm3) | 1.13 | 1.128 |
| 持水量Water holding capacity/% | 42.36 | 44.07 |
| 阳离子交换量Cation exchange capacity/(cmol/kg) | 7.25 | 13.68 |
分别于2015年3月烟田休耕期、6月烟苗旺长期(青枯病发病初期)和9月烟叶成熟期(青枯病发病后期)对青枯病发病样点和不发病对照样点各3个实验样地(田块)进行土壤取样。取样时, 3月, 3个实验样地均设5个重复采样点(均匀布置), 每个采集点间距约为10 m;6月和9月, 3个实验样地均设6个重复采样点(均匀布置), 每个采集点间距约为10 m。3月烟田休耕期的样点均选择垄作烟田中较为平坦、人为活动少的垄台采样, 每个点并排同时采样2次, 分别用于调整的贝尔曼(Bearmann)湿漏斗和Tullgren干漏斗分离湿生动物和其他土壤动物;而6月和9月烟草生育期的采样点均选择垄台靠近烟苗主茎两侧取样根际土壤。每个采样点由上到下分为0—5、5—10 cm和10—15 cm共3个土层取样。
采用口径为8 cm、高度为5 cm的环刀采样器对各土壤分层取样, 分层采集土样后按个层次分别进行土样铝盒装盒保存。土样拿回室内后, 采用直径为2 mm的Tullgren干漏斗分离土壤动物;土壤动物的分离在各个干漏斗烘虫箱中进行, 控制烘虫箱温度在30—50 ℃, 烘虫时间为48 h;分离出的土壤动物用盛有75%浓度的酒精收集于玻璃瓶中, 待分类鉴定并计数。线虫等湿生动物在贝尔曼湿漏斗中进行分离;经48 h后取样15 mL漏斗收集管中的水溶液镜检和统计。
将分离获得的土壤动物置于双目解剖镜下鉴定。参考鉴定资料《中国亚热带土壤动物》[19]和《中国土壤动物检索图鉴》[20]进行分类鉴定。由于受分类限制, 采用大类进行分类并统计数量。
2.3 数据处理与分析土壤动物群落内多样性(α多样性)指数测度分析主要采用Simpson优势集中性指数(C)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)和Margalef丰富度指数(D)来进行计算。
Simpson优势集中性指数(C):
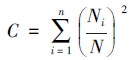
|
Shannon-Wiener多样性指数(H′):
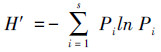
|
Pielou均匀度指数(J):
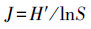
|
Margalef丰富度指数(D):
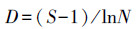
|
式中, Ni为第i类群的个体数;N为群落中所有类群的个体数;Pi=Ni/N, S为类群数。由于线虫在土壤动物群落中的密度很大, 因此, 为了深入分析线虫和干漏斗中土壤动物(以螨类和弹尾虫为主)与青枯病危害的关系, 对上述4个土壤动物群落特征指数的计算分为有或无线虫两种情况并分别统计分析。
土壤动物群落间多样性(β多样性)系数分析主要采用Sørensen相似性系数来进行计算。该系数主要反映两类生境烟田土壤动物类群上的相似程度。
Sørensen相似性系数(Cs):
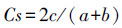
|
式中, a为发病烟田全部类群数, b为不发病对照烟田(CK)全部类群数, c为两类烟田共有的类群数。该公式规定的相似性等级是:计算值在0.75—1.00为极相似, 在0.50—0.74为中等相似, 在0.25—0.49为中等不相似, 在0—0.24为极不相似。
土壤动物群落相对稳定性测定采用群落物种数与个体数之比(St/Si)表示。该值反映种间数量上的制约作用[21]。由于土壤动物群落中螨类和弹尾虫发生量大, 它们之间的数量比(A/C, A代表螨类, C代表弹尾虫)与烟田青枯病危害存在相关性[17], 因此统计分析了两类烟田不同调查期该比值的变化规律。
分离的土壤动物个体数量(多度)占捕获总量的百分比为相对多度。其中, 10.0%以上者为优势类群, 占1.0%—10.0%者为常见类群, 不足1.0%者为稀有类群。土壤动物密度数据均为分离的土壤动物个体数换算成的平均密度(个/m2)。数据分析前, 对土壤动物多度和密度数据进行log(x+1)对数转换;对土壤动物相对多度进行Arcsin(sqrt(x))平方根反正旋转换。对于服从正态分布和方差齐性的土壤动物数据进行单因素方差分析(One-way ANOVA)。对于不服从正态分布和方差齐性的土壤动物数据进行两独立样本的非参数Mann-Whitney U秩和检验。
主成分分析(Principal component analysis, PCA):对两类生境烟田的土壤动物群落主要特征指标进行主成分分析, 并根据各烟田优势和常见土壤动物类群PCA得分对土壤动物采样点进行排序分析。
以上分析和作图采用Microsoft Excel 2007、SPSS 19.0以及CANOCO 5软件完成。
3 结果与分析 3.1 不同烟田土壤动物类群组成和数量特征通过调查共捕获中小型土壤动物50112只, 隶属于3门13纲33类。3月休耕烟田(简称3月), 线虫相对多度为80.91%, 属于优势类群;等节跳虫科、中气门亚目、甲螨亚目、棘跳虫科、前气门亚目和长角跳虫科依次是8.84%, 2.39%, 2.17%, 1.52%, 1.10%和1.02%, 为常见类群;其他2.02%均为稀有类群。6月烟苗旺长期(简称6月), 线虫和中气门亚目相对多度分别为66.38%和10.88%, 属于优势类群;棘跳虫科、甲螨亚目、长角跳虫科、啮虫目、等节跳虫科和前气门亚目依次为5.45%, 4.87%, 4.06%, 2.13%, 2.48%和1.22%, 为常见类群;其他2.53%均为稀有类群。9月烟叶成熟期(简称9月), 线虫相对多度为68.64%, 属于优势类群;中气门亚目、甲螨亚目、等节跳虫科、棘跳虫科、长角跳虫科、鞘翅目和短角跳虫科依次是9.95%, 7.19%, 3.73%, 2.37%, 1.95%, 1.78%和1.01%, 为常见类群;其他3.38%为稀有类群。因此, 优势类群和常见类群构成了烟田中小型土壤动物的主体。
青枯病烟田(简称青枯病田)及其对照无青枯病田(简称对照田)土壤动物优势和常见类群及其相对多度存在一定差异。3月, 青枯病田优势类群是线虫和等节跳虫科, 常见类群是甲螨亚目、中气门亚目、棘跳虫科、长角跳虫科和前气门亚目;对照田优势类群是线虫, 常见类群是等节跳虫科、中气门亚目、甲螨亚目和原跳虫科;在两者共有的优势和常见类群中, 青枯病田线虫的相对多度显著低于对照(F=9.082, P < 0.01), 等节跳虫科和甲螨亚目均显著大于对照(F=10.577, P < 0.01;F=4.384, P < 0.05), 中气门亚目差异不显著(F=0.175, P>0.05)。6月, 青枯病田优势类群是线虫和中气门亚目, 优势类群是棘跳虫科、长角跳虫科、甲螨亚目、啮虫目和前气门亚目;对照田优势类群是线虫, 常见类群是甲螨亚目、中气门亚目、棘跳虫科、长角跳虫科、等节跳虫科、前气门亚目和短角跳虫科;在两者共有的优势和常见类群中, 青枯病田线虫、中气门亚目、甲螨亚目、前气门亚目、棘跳虫科和长角跳虫科的相对多度均与对照差异不显著(F=0.013—4.399, P>0.05)。9月, 青枯病田优势类群是线虫, 常见类群是中气门亚目、甲螨亚目、等节跳虫科、棘跳虫科、长角跳虫科、鞘翅目、短角跳虫科和前气门亚目;对照田优势类群是线虫和中气门亚目, 常见类群是甲螨亚目、等节跳虫科、棘跳虫科、膜翅目和长角跳虫科;在两者共有的优势和常见类群中, 青枯病田中气门亚目和等节跳虫科的相对多度均显著低于对照(U=117.0, P < 0.05;F=7.292, P < 0.01), 线虫、甲螨亚目、棘跳虫科和长角跳虫科在两者间差异不显著(F=0.025—3.488, P>0.05)。
3.2 不同烟田土壤动物群落密度及其动态特征 3.2.1 土壤动物群落密度的时间动态青枯病田及其对照田土壤动物总密度(图 1A)和线虫密度(图 1B)均为3月休耕期大于6月和9月生育期;其中, 3月青枯病田和对照田土壤动物密度差异不显著(F=0.796, P>0.05), 而对照田线虫密度显著大于青枯病田(F=6.77, P < 0.05);6月和9月土壤动物和线虫密度均为青枯病田显著大于对照田(土壤动物:F6月=16.08, P < 0.01;F9月=8.67, P < 0.01;线虫:F6月=7.07, P < 0.05;F9月=8.45, P < 0.01)。螨类在两类烟田密度以生育期为高(图 2A);除了6月青枯病田显著大于对照田(F=9.31, P < 0.01)外, 3月和9月均差异不显著(P>0.05)。弹尾虫在两类烟田生境密度为休耕期大于生育期(图 2B), 3个时期的密度均为青枯病田显著大于对照田(F3月=5.54, P < 0.05;F6月=18.59, P < 0.01;F9月=9.29, P < 0.01)。其他稀有类群密度以生育期为高(图 2C);6月和9月均为青枯病田显著大于对照田(F6月=11.22, P < 0.01;F9月=11.29, P < 0.01), 3月差异不显著(P>0.05)。

|
| 图 1 不同调查期两类烟田土壤动物群落密度(平均值±标准误) Fig. 1 Densities of soil animal community in tobacco fields during different period of investigation (Mean±SE) *表示青枯病田与对照田差异显著(P < 0.05) |

|
| 图 2 不同调查期两类烟田土壤动物群落密度(平均值±标准误) Fig. 2 Densities of soil faunal community in tobacco fields during different period of investigation (Mean±SE) |
在3个调查期, 青枯病田及其对照田土壤动物总密度及其各类群密度在0—5、5—10 cm和10—15 cm 3个土层均存在明显变化。3月, 土壤动物和线虫密度(图 1)在5—10 cm和10—15 cm均为对照田显著高于青枯病田(土壤动物:F5—10=10.17, P < 0.01;F10—15=6.15, P < 0.05;线虫:F5—10=11.26, P < 0.01;F10—15=18.21, P < 0.01);6月和9月均为青枯病田高于对照, 但土壤动物只有6月0—5 cm以及9月0—5 cm和10—15 cm存在显著差异(6月:F0—5=16.63, P < 0.01;9月:U0—5=96.0, P < 0.01;U10—15=137.0, P < 0.05), 而线虫6月0—5 cm及9月3个土层均存在显著差异(6月:F0—5=6.34, P < 0.05;9月:U0—5=138.5, P < 0.05;U5—10=112.0, P < 0.05;U10—15=125.5, P < 0.05)。3月10—15 cm和6月0—5 cm的螨类密度为青枯病田显著高于对照田(3月:F10—15=6.89, P < 0.05;6月:F0—5=6.59, P < 0.05), 其他时间各土层均差异不显著(P>0.05)。3个时间3个土层弹尾虫密度均为青枯病田高于对照田, 但3月、6月和9月的0—5 cm(F3月=7.95, P < 0.01;F6月=14.85, P < 0.01;F9月=6.07, P < 0.05)以及6月的5—10 cm和10—15 cm存在显著差异(F5—10=9.79, P < 0.01;F10—15=6.49, P < 0.05)。其他稀有类群密度, 6月10—15 cm和9月0—5 cm为青枯病田显著高于对照(U10—15=2.0, P < 0.01;F0—5=99.0, P < 0.05), 其他各时间的土层差异均不显著(P>0.05)(图 2)。
3.3 不同烟田土壤动物群落多样性指数及其动态由图 3可知, 在统计分析时线虫的有或无对两类生境烟田的计算结果有较大影响。3月的Shannon-Wiener多样性指数在有或无线虫2种条件下均为青枯病田显著高于对照田(F有=83.21, P < 0.01;F无=92.03, P < 0.01);6月无线虫条件下对照田显著高于青枯病田(F无=17.21, P < 0.01), 而9月为青枯病田显著高于对照田(F无=47.42, P < 0.01)。Pielou均匀度指数只在6月的无线虫条件下对照田显著高于青枯病田(F无=35.65, P < 0.01), 其他均差异不显著(P>0.05)。3月的Simpson优势集中性指数在线虫条件下为对照田显著高于青枯病田(F有=6.69, P < 0.05), 而无线虫时青枯病田显著高于对照(F无=4.72, P < 0.05);6月和9月均为无线虫时对照田显著高于青枯病田(F无=11.71, P < 0.01;U无=60.0, P < 0.01)。3月和6月的Margalef丰富度指数均为对照田大于青枯病田, 其中3月无线虫和6月有或无线虫条件下均差异显著(3月:F无=7.81, P < 0.01;6月:F有=5.64, P < 0.05;F无=6.98, P < 0.05);但9月在这2种条件下均为青枯病田显著高于对照田(F有=49.41, P < 0.01;F无=95.85, P < 0.01)。

|
| 图 3 不同调查时间烟田土壤动物群落多样性(平均值±标准误) Fig. 3 Diversities of soil animal community in tobacco fields during different period of investigation (Mean ± SE) |
通过计算, 青枯病烟田及其对照田不同调查期土壤动物群落Sørensen相似性系数均大于0.75, 说明2类烟田土壤动物群落处于极相似程度;同时发现Cs值3月最高, 9月最低, 表明群落空间相似性程度也受季节或烟草成熟期青枯病危害加剧的影响(表 2)。两类烟田不同季节土壤动物群落稳定性St/Si比值存在明显不同, 3月的比值尽管青枯病田大于对照田, 但两者比值相差较小;而两类烟田6月和9月的比值差异较大, 均为对照田大于青枯病田(表 2)。
| 烟田Tobacco fields | 群落相似性Community similarity | 群落稳定性Community stability | |||||
| 3月March | 6月June | 9月September | 3月March | 6月June | 9月September | ||
| 青枯病田Hazard fields of tobacco wilt | 0.87 | 0.85 | 0.80 | 0.0019 | 0.0032 | 0.0022 | |
| 对照田Control fields without hazard | 0.0015 | 0.0111 | 0.0044 | ||||
在3个调查期, 螨类与弹尾虫个体数的A/C比值均为对照烟田大于青枯病田;且该比值在两类烟田均为3月>6月>9月;其中, 3月和6月对照田均显著性高于青枯病田(F3月=5.65, P < 0.05;F6月=9.96, P < 0.01);说明A/C比值的变化与青枯病的发生存在密切关系(图 4)。

|
| 图 4 不同调查期两类烟田螨类与弹尾虫A/C比值的动态变化(平均值±标准误) Fig. 4 The values of A/C in individual numbers between Acari and Collembola in tobacco fields during the investigation period (Mean±SE) |
本研究对土壤动物个体数、类群、多样性指数(H′)、均匀性指数(J)、丰富度指数(D)、优势度指数(C)、类群密度、A/C比值以及不同调查期的优势类群和常见类群等土壤动物特征指标采用主成分法进行了分析, 确定对青枯病危害敏感的指示土壤动物特征指标。3个调查期共有的优势类群和常见类群包括线虫、甲螨亚目、中气门亚目、前气门亚目、棘跳虫科、等节跳虫科和长角跳虫科等7类;此外, 3月还包括原跳虫科;6月有短角跳虫科和啮虫目;9月有短角跳虫科、鞘翅目和膜翅目。本研究选取前4个主成分, 如表 3所示, 其中3个调查时间的累积方差贡献率在74%—85%, 可以包含上述土壤动物特征指标的绝大部分信息。
| 时间 Time |
主成分 Principal component |
特征根 Eigenvalues |
方差贡献率/% Variance contribution |
累积方差贡献率/% Cumulative variance contribution |
| 3月March | 1 | 4.16 | 25.97 | 25.97 |
| 2 | 3.49 | 21.81 | 47.78 | |
| 3 | 2.37 | 14.79 | 62.57 | |
| 4 | 1.88 | 11.72 | 74.29 | |
| 6月June | 1 | 5.96 | 35.06 | 35.06 |
| 2 | 4.79 | 28.16 | 63.22 | |
| 3 | 2.07 | 12.16 | 75.38 | |
| 4 | 1.58 | 9.27 | 84.64 | |
| 9月September | 1 | 6.80 | 37.79 | 37.79 |
| 2 | 3.86 | 21.42 | 59.21 | |
| 3 | 2.07 | 11.51 | 70.72 | |
| 4 | 1.39 | 7.75 | 78.46 |
以每个主成分的对应特征值占所选取的4个主成分特征值之和的比值作为权重, 分别与对应的主成分值相乘后求和得到每个土壤动物特征指标的综合分值, 然后进行排序[22], 结果如表 4所示。由表 4可知, 不同季节烟田土壤动物各特征指标对青枯病危害的指示作用明显不同。其中, 3月综合分值前5名依次为中气门亚目、土壤动物个体数、土壤动物密度、线虫和A/C值, 表明烟田休耕期这5个土壤动物指标对青枯病的发生危害最为敏感;6月综合分值前5名依次为土壤动物密度、土壤动物个体数、中气门亚目、啮虫目和线虫, 表明烟株旺长期(青枯病发病初期)这5个中小型土壤动物指标对青枯病的发生危害最为敏感;9月综合分值前5名依次为土壤动物个体数、线虫、中气门亚目、甲螨亚目和土壤动物密度, 表明烟叶成熟期(青枯病发病盛期)这5个中小型土壤动物指标对青枯病的发生危害最为敏感;上述3个时期的土壤动物指标均可作为分析青枯病危害作用的重要特征指标。另外, 6月棘跳虫科、等节跳虫科、长角跳虫科和短角跳虫科及9月A/C值、前气门亚目和棘跳虫科排名靠前, 也可分别作为各时期分析青枯病危害作用的参考指标。
| 时间 Time |
指标 Index |
主成分1 Principal component 1 |
主成分2 Principal component 2 |
主成分3 Principal component 3 |
主成分4 Principal component 4 |
综合分值 Comprenhensive values |
排序 Rank |
| 3月 March |
个体数Individual number | 0.579 | 0.739 | 0.108 | -0.142 | 0.213 | 2 |
| 类群数Group number | 0.150 | 0.393 | -0.478 | 0.458 | 0.078 | 11 | |
| 密度Density | 0.871 | 0.398 | -0.025 | -0.216 | 0.184 | 3 | |
| D | 0.051 | 0.329 | -0.702 | 0.551 | 0.033 | 14 | |
| H′ | -0.858 | 0.299 | 0.101 | 0.148 | -0.070 | 15 | |
| J | 0.536 | 0.051 | -0.100 | 0.393 | 0.132 | 7 | |
| C | 0.706 | -0.292 | -0.008 | 0.144 | 0.091 | 9 | |
| A/C值A/C value | 0.263 | -0.081 | 0.799 | 0.106 | 0.148 | 5 | |
| 线虫Nematode | 0.776 | 0.461 | 0.007 | -0.359 | 0.165 | 4 | |
| 甲螨亚目Oribatida | -0.166 | 0.794 | 0.354 | -0.082 | 0.133 | 6 | |
| 中气门亚目Mesostigmata | 0.157 | 0.370 | 0.716 | 0.390 | 0.223 | 1 | |
| 前气门亚目Prostigmata | 0.007 | -0.266 | 0.280 | 0.718 | 0.078 | 12 | |
| 原跳虫科Poduridae | 0.716 | -0.241 | -0.222 | 0.279 | 0.088 | 10 | |
| 棘跳虫科Onychiuridae | -0.172 | 0.844 | 0.007 | 0.174 | 0.124 | 8 | |
| 等节跳虫科Isotomidae | -0.410 | 0.588 | 0.064 | 0.256 | 0.060 | 13 | |
| 长角跳虫科Entomobryidae | -0.284 | 0.411 | -0.453 | -0.334 | -0.081 | 16 | |
| 6月 June |
个体数Individual number | 0.855 | 0.432 | 0.254 | 0.037 | 0.240 | 2 |
| 类群数Group number | -0.544 | 0.276 | 0.412 | 0.472 | 0.032 | 10 | |
| 密度Density | 0.855 | 0.432 | 0.254 | 0.038 | 0.240 | 1 | |
| D | -0.876 | -0.012 | 0.187 | 0.304 | -0.105 | 17 | |
| H′ | -0.769 | 0.582 | 0.130 | 0.004 | -0.029 | 15 | |
| J | -0.713 | 0.589 | 0.033 | -0.129 | -0.039 | 16 | |
| C | 0.722 | -0.620 | -0.135 | 0.106 | 0.024 | 11 | |
| A/C值A/C value | -0.024 | -0.399 | 0.830 | -0.295 | -0.006 | 13 | |
| 线虫Nematode | 0.964 | 0.136 | 0.051 | 0.130 | 0.201 | 5 | |
| 甲螨亚目Oribatida | -0.557 | 0.386 | 0.222 | -0.073 | -0.020 | 14 | |
| 中气门亚目Mesostigmata | 0.585 | 0.417 | 0.633 | -0.011 | 0.225 | 3 | |
| 前气门亚目Prostigmata | -0.003 | 0.238 | 0.434 | -0.708 | 0.017 | 12 | |
| 棘跳虫科Onychiuridae | 0.091 | 0.904 | -0.108 | 0.042 | 0.146 | 6 | |
| 等节跳虫科Isotomidae | 0.137 | 0.766 | -0.324 | 0.023 | 0.109 | 8 | |
| 长角跳虫科Entomobryidae | 0.378 | 0.632 | -0.299 | -0.134 | 0.119 | 7 | |
| 短角跳虫科Neelidae | 0.147 | -0.342 | 0.399 | 0.690 | 0.073 | 9 | |
| 啮虫目Corrodentia | 0.192 | 0.888 | 0.038 | 0.350 | 0.202 | 4 | |
| 9月 September |
个体数Individual number | 0.677 | 0.685 | 0.108 | -0.023 | 0.229 | 1 |
| 类群数Group number | 0.925 | 0.100 | -0.262 | 0.133 | 0.169 | 6 | |
| 密度Density | 0.924 | 0.106 | -0.253 | 0.141 | 0.171 | 5 | |
| D | 0.825 | -0.178 | -0.381 | 0.121 | 0.099 | 11 | |
| H′ | 0.557 | -0.805 | 0.076 | -0.045 | -0.005 | 16 | |
| J | 0.046 | -0.937 | 0.246 | -0.150 | -0.109 | 18 | |
| C | -0.372 | 0.811 | -0.259 | 0.079 | 0.024 | 15 | |
| A/C值A/C value | 0.111 | 0.227 | 0.866 | 0.274 | 0.163 | 7 | |
| 线虫Nematode | 0.428 | 0.861 | -0.032 | -0.060 | 0.191 | 2 | |
| 甲螨亚目Oribatida | 0.661 | 0.245 | 0.370 | -0.260 | 0.172 | 4 | |
| 中气门亚目Mesostigmata | 0.617 | -0.039 | 0.653 | 0.144 | 0.187 | 3 | |
| 前气门亚目Prostigmata | 0.717 | 0.087 | -0.130 | 0.273 | 0.154 | 9 | |
| 棘跳虫科Onychiuridae | 0.733 | 0.030 | 0.330 | -0.206 | 0.156 | 8 | |
| 等节跳虫科Isotomidae | 0.358 | -0.327 | -0.391 | -0.255 | -0.041 | 17 | |
| 长角跳虫科Entomobryidae | 0.720 | -0.058 | 0.033 | -0.474 | 0.089 | 12 | |
| 短角跳虫科Neelidae | 0.638 | -0.289 | -0.212 | 0.174 | 0.071 | 13 | |
| 鞘翅目Coleoptera | 0.613 | 0.066 | -0.099 | 0.000 | 0.112 | 10 | |
| 膜翅目Hymenoptera | 0.094 | -0.299 | 0.000 | 0.841 | 0.046 | 14 |
青枯病及其对照田土壤动物优势和常见类群PCA得分排序图见图 5。由图 5可知, 在3个调查期青枯病田及其对照田各土样采集点分布的梯度聚集特点明显, 两者间少数样点有重叠;主要土壤动物类群在2类烟田的多度均较大, 说明两类生境烟田土壤动物群落组成及结构相似度高。其中, 3月各样点聚集分布最为显著, 只有3个分类区, 两类烟田样点均有重叠分布, 各主要类群多度在两类烟田均较大;6月有4个分类区, 两类烟田采样点只在3个分类区有重叠分布, 前气门亚目在对照田部分采样点多度较大;9月有4个分类区, 两类烟田采样点只在一个分类区有重叠分布, 线虫、棘跳虫科和长角跳虫科在青枯病田部分采样点多度较大, 而鞘翅目和膜翅目在对照田部分样采点多度较大;说明两类烟田土壤动物群落的差异随着青枯病危害程度的加剧而加大, 差异程度9月>6月>3月。

|
| 图 5 烟草青枯病及其对照烟田土壤动物主要类群的PCA排序图 Fig. 5 PCA ordination diagramse of soil animals in tobacco fields and its control during different period of investigations ○代表青枯病烟田采样点Hazard fields of tobacco wilt;△代表无青枯病烟田采样点(CK)Control fields without hazard;Ori:甲螨亚目Oribatida;Mes:中气门亚目Mesostigmata;Pros:前气门亚目Prosfigmata;Pod:原跳虫科Poduridae;Ony:棘跳虫科Onychiuridae;Entom:长角跳虫科Entomobryidae;Isot:等节跳虫科Isotomidae;Nee:短角跳虫科Neelidae;Cor:啮虫目Corrodentia;Col:鞘翅目Coleoptera;Hym:膜翅目Hymenoptera;Nem:线虫Nematoda |
青枯病在各种农作物上的发生机理和控制措施一直是国内外农业生产上研究的热点问题。尤其是近年来随着我国各烟区烟田连作年限的不断增加, 导致该病害的流行和扩散速度在不断加快。研究指出, 对根际土传植物病害进行成功的生物防治, 必须建立在对土壤微生物和土壤动物的组成及相互关系了解的基础上[15]。土壤动物与农田土壤系统中的生物和非生物因素关系密切, 可作为土壤健康的有效指示类群[23]。前期的研究表明, 青枯病危害程度与根际土壤螨类和弹尾虫的发生存在显著的相关性[17]。本研究结果进一步表明, 重庆山地烟田中小型土壤动物主要由线虫和节肢动物构成, 相对多度超过了95%;它们中的优势类群和常见类群对烟田青枯病病原菌的繁殖和侵染起重要作用;烟田青枯病的发生危害能显著影响中小型土壤动物群落的组成及结构特征, 特别是土壤线虫、螨类和弹尾虫及其主要类群对烟田青枯病的发生有重要指示作用。目前, 大量的研究表明青枯病病原菌的发生危害与土壤微生物存在密切关系[24-26];但与土壤动物的关系如何, 鲜有报道。此外, 烟田不同时期调查结果表明, 烟田土壤动物群落组成具有明显的季节特征, 休耕期土壤动物总密度和线虫密度高于生育期, 即3月>6月>9月;螨类和其他土壤动物的密度均以生育期为高, 而弹尾虫则以休耕期为高;这种变化规律主要受温度和降雨量[27]、土壤营养[28]以及烟田不同季节农事耕作和管理的人为扰动[29]的综合影响;这些因素显著影响土壤理化特性、土壤动物的摄食活动及其微生境的变化, 最终对土壤动物的发生量起到决定性影响。另外, 植物根系作为根际的主要调控者, 对根际土壤微生物和土壤动物种群分布及其结构也可造成重要影响[30], 从而导致烟田休耕期和生育期土壤动物密度出现差异。
土壤动物群落内的多样性(α多样性)指标反应群落内的稳定性, 主要包括个体数、密度和群落多样性指数等[31]。本研究发现, 烟草青枯病田土壤动物总量、线虫、螨类和弹尾虫发生量均要高于无青枯病田, 特别是烟草生育期比休耕期表现的更为显著。同时, 青枯病烟田及其对照田优势和常见类群及其相对多度明显不同;两类烟田不同烟草生产阶段特征优势类群也存在差异。3月烟田休耕期, 青枯病田特征优势类群是棘跳虫科、长角跳虫科和前气门亚目, 而对照田是原跳虫科;6月烟株旺长期, 青枯病田特征优势类群是啮虫目, 对照田是等节跳虫科和短角跳虫科;9月烟叶成熟期, 青枯病田特征优势类群是鞘翅目、短角跳虫科和前气门亚目, 而对照田是膜翅目。在两者共有的优势和常见类群中, 休耕期, 青枯病田等节跳虫科和甲螨亚目均显著大于对照田;烟株旺长期, 青枯病田中气门亚目和等节跳虫科均显著低于对照田。青枯病的发生也能显著影响各季节烟田不同土层动物及其主要类群的数量分布。螨类与弹尾虫个体数的A/C比值也与青枯病的发生存在密切关系。青枯病危害也对土壤动物多样性指数产生了显著影响, 且这些变化在不同季节呈现明显不同。Margalef丰富度指数、Shannon-Wiener多样性指数和Simpson优势集中性指数在不同季节受青枯病危害的影响较大;而Pielou均匀度指数在青枯病及其对照田变化较小。上述这些变化与青枯病影响线虫、螨类和弹尾虫等优势类群的发生存在密切的相关性。相关的研究指出, 青枯病病原菌在土壤中能显著影响不同季节土壤中的细菌、真菌和放线菌等土壤微生物群落[23];从而影响土壤动物与微生物在土壤生态系统物质循环和能量流动中的互作关系[12]。其中, 土壤线虫主要以微生物为食, 并能调节微生物的群落结构和生物量[32];土壤螨类和弹尾虫以腐食性为主, 也取食微生物, 能影响土壤微生物的群落结构[14]。因此, 可推测青枯病的发生直接或间接地影响了土壤微生物群落的组成和多样性, 最终对土壤动物群落的组成和功能特性产生了作用。
土壤动物群落间的多样性(β多样性)指标主要反应群落间环境异质性的大小或群落间物种组成的差异, 其与α多样性一起构成了群落或生态系统总体多样性或一定地段的生物异质性[33]。本研究群落相似性结果表明, 青枯病田及对照烟田不同季节土壤动物群落物种组成相似度很高, 说明两类烟田的共有种较多, 物种更替速率越低;尽管如此, 群落间异质性程度也受到了青枯病危害的较大影响, 特别是烟叶成熟期群落相似性明显低于休耕期和烟株旺长期。另外, 除了休耕期, 两类烟田土壤动物群落稳定性均为对照田大于青枯病田, 说明对照田土壤动物物种数相对于个体数而言较多, 土壤动物种类分布相对均衡, 种间制约作用较强。两类烟田的这种差异也可参照营养级联反应(Cascading trophic interactions)[34]或上行/下行理论(Bottom-up/top-down theory)[35]来解释。尽管这两种理论的实质都是建立在以捕食者为主要调控因子的基础上, 但它们在陆地生态系统中的研究内容都强调以植物为中心的地上部分(Aboveground-)和地下部分(Belowground-)生物群的生态关联(Ecological linkages)[36-37]。与对照田相比, 烟草生育期土壤青枯病病原菌、土壤生物(包括土壤动物)和植物根系的互作易导致青枯病田烟株地上部分生长势减弱(可看作上行作用), 反之, 青枯病田烟株地上部分变化(如烟叶衰弱和茎秆腐烂及其带来的生物类群的减少)也会影响到根系的生长, 从而对土壤生物(包括青枯病病原菌和土壤动物)的发生产生重大影响(可看作下行作用), 最终造成两类烟田在中小型土壤动物群落物种组成上出现较大差异。
主成分分析结果进一步指出, 不同季节影响青枯病发生的土壤动物主要指标存在明显差异, 可作为植烟区各时期分析青枯病危害作用的参考特征指标, 如土壤动物个体数、密度和A/C值, 以及中气门亚目、线虫、甲螨亚目、棘跳虫科和等节跳虫科等土壤动物类群。另外, 基于两类烟田土壤动物群落优势和常见类群主成分得分的土样采集点排序结果进一步说明, 两类烟田土壤动物群落组成相似度高, 但土壤动物群落间的异质性也明显受到了青枯病危害的影响。因此, 在该病害生物防治的生产实践中, 应充分研究和利用土壤动物对该病原菌危害的这些指示作用。目前这方面工作开展的较少, 应不断加强相关内容的基础研究。报道指出, 根际线虫直接参与根际土壤生态系统的物质循环和能量流动, 对植物的生长尤其是抗病性影响很大[38]。例如用青枯菌单独接种引起个别叶片轻度萎蔫, 用根结线虫Meloidogyne incognita单独接种萎蔫叶片更少, 而将两者同时接种时植株严重萎蔫死亡, 而且病害病状出现的时间提前[39]。土壤食真菌螨通过取食根际真菌, 可刺激某些根系有益细菌的繁殖;而一些食土壤动物螨既能降低一些植食有害动物数量, 也会降低一些有益土壤动物(腐食类线虫、昆虫和螨)的密度[14]。迄今, 有一些弹尾虫抑制土传病原菌方面的报道, 但仅限于对病原真菌的探索。Dromph和Borgen[40]研究表明, 等节跳虫科中的Folsomia fimetaria和Priostoma minuta能降低接种到土壤中土传病原菌小麦腥黑粉菌Tilletia tritici冬孢子的比例;且P. minuta种群密度的明显增加显著减少了小麦黑粉病的发病率。Wolfarth等[16]室内研究发现, 线虫Aphelenchoides saprophilus和等节跳虫科中的白符跳虫F. candida无论是单独或结合使用, 都很好地抑制了土传根腐病真菌Fusarium culmorum的生长, 并且对其分泌的霉菌毒素起到了降解作用。
总之, 食性多样化的中小型土壤动物与土壤微生物(包括青枯病病原菌)存在着复杂的直接营养关系和非营养关系;烟草青枯病的爆发危害显然会影响到烟田土壤中小型动物的发生, 而土壤中小型动物可对烟草青枯病的发生和危害起到有效的生物指示作用;在农业生产实践上要开展可持续和有效的烟草青枯病生物防治, 应不断加强该病原菌与土壤中小型动物生态关系的基础研究。
致谢: 西南大学植物保护学院喻延和李石力同学协助田间调查,美国密苏里大学Qisheng Song教授帮助写作, 特此致谢。| [1] | Hayward A C. Biology and epidemiology of bacterial wilt caused by Pseudomonas Solanacearum. Annual Review of Phytopathology, 1991, 29(1): 65–87. DOI:10.1146/annurev.py.29.090191.000433 |
| [2] | 喻延, 李石力, 杨亮, 丁伟. 烟草青枯病防治技术的集成应用. 植物医生, 2016, 29(4): 55–56. |
| [3] | 郑世燕, 丁伟, 杜根平, 杨亮, 刘晓姣, 张永强. 增施矿质营养对烟草青枯病的控病效果及其作用机理. 中国农业科学, 2014, 47(6): 1099–1110. |
| [4] | Swift M J, Heal O W, Anderson J M. Decomposition in Terrestrial Ecosystems. Oxford: Blackwell Scientific Publications, 1979. |
| [5] | Büchs W. Biotic indicators for biodiversity and sustainable agriculture-introduction and background. Agriculture, Ecosystems & Environment, 2003, 98(1/3): 1–16. |
| [6] | 宋理洪, 武海涛, 吴东辉. 我国农田生态系统土壤动物生态学研究进展. 生态学杂志, 2011, 30(12): 2898–2906. |
| [7] | 刘训理, 王超, 吴凡, 薛东红, 陈凯. 烟草根际微生物研究. 生态学报, 2006, 26(2): 552–557. |
| [8] | Niu J J, Rang Z W, Zhang C, Chen W, Tian F, Yin H Q, Dai L J. The succession pattern of soil microbial communities and its relationship with tobacco bacterial wilt. BMC Microbiology, 2016, 16: 233. DOI:10.1186/s12866-016-0845-x |
| [9] | 傅声雷. 土壤生物多样性的研究概况与发展趋势. 生物多样性, 2007, 15(2): 109–115. |
| [10] | 柯欣, 梁文举, 宇万太, 谢荣栋, 翁朝联, 杨毅明, 尹文英. 下辽河平原不同土地利用方式下土壤微节肢动物群落结构研究. 应用生态学报, 2004, 15(4): 600–604. |
| [11] | Bonkowski M, Cheng W X, Griffiths B S, Alphei J, Scheu S. Microbial-faunal interactions in the rhizosphere and effects on plant growth. European Journal of Soil Biology, 2000, 36(3/4): 135–147. |
| [12] | Bonkowski M, Villenave C, Griffiths B. Rhizosphere fauna:the functional and structural diversity of intimate interactions of soil fauna with plant roots. Plant and Soil, 2009, 321(1/2): 213–233. |
| [13] | 邵元虎, 张卫信, 刘胜杰, 王晓丽, 傅声雷. 土壤动物多样性及其生态功能. 生态学报, 2015, 35(20): 6614–6625. |
| [14] | Smrž J, Čatská V. The effect of the consumption of some soil fungi on the internal microanatomy of the mite Tyrophagus putrescentiae (Schrank) (Acari:Acaridida). Acta Universitatis Carolinae-Biologica, 1989, 33: 81–93. |
| [15] | Lartey R T. Dynamics of soil flora and fauna in biological control of soil inhabiting plant pathogens. Plant Pathology Journal, 2006, 5(2): 125–142. DOI:10.3923/ppj.2006.125.142 |
| [16] | Wolfarth F, Schrader S, Oldenburg E, Weinert J. Nematode-collembolan-interaction promotes the degradation of Fusarium biomass and deoxynivalenol according to soil texture. Soil Biology and Biochemistry, 2013, 57: 903–910. DOI:10.1016/j.soilbio.2012.11.001 |
| [17] | 白耀宇, 庞帅, 李石力, 丁伟. 烟草青枯病危害对烟株根际土壤螨类和弹尾虫发生的影响. 西南大学学报: 自然科学版, 2015: 1–6. |
| [18] | 鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000. |
| [19] | 尹文英. 中国亚热带土壤动物. 北京: 科学出版社, 1992. |
| [20] | 尹文英. 中国土壤动物. 北京: 科学出版社, 2000. |
| [21] | 高宝嘉, 张执中, 李镇宇. 封山育林对昆虫群落结构及多样性稳定性影响的研究. 生态学报, 1992, 12(1): 1–7. |
| [22] | 杨旭, 高梅香, 张雪萍, 林琳, 沙迪, 张利敏. 秸秆还田对耕作黑土中小型土壤动物群落的影响. 生态学报, 2017, 37(7): 2206–2216. |
| [23] | Pankhurst C E, Hawke B G, McDonald H J, Kirkby C A, Buckerfield J C, Michelsen P, O'Brien K A, Gupta V V S R, Doube B M. Evaluation of soil biological properties as potential bioindicators of soil health. Australian Journal of Experimental Agriculture, 1995, 35(7): 1015–1028. DOI:10.1071/EA9951015 |
| [24] | Shiomi S, Yamamoto M, Nakamura R, Inaba A. Expression of ACC synthase and ACC oxidase genes in melons harvested at different stages of maturity. Journal of the Japanese Society for Horticultural Science, 1999, 68(1): 10–17. DOI:10.2503/jjshs.68.10 |
| [25] | 匡传富, 何志明, 汤若云, 黄声仪, 邓芳席. 烟草青枯病土壤微生物数量及生理群的测定. 中国烟草科学, 2003, 24(1): 43–45. |
| [26] | 郑雪芳, 刘波, 蓝江林, 朱育菁, 车建美, 苏明星. 无致病力青枯雷尔氏菌对烟草根系土壤微生物脂肪酸生态学特性的影响. 生态学报, 2012, 32(14): 4496–4504. |
| [27] | 宋敏. 增加降水及施氮对弃耕草地土壤线虫和小型节肢动物的影响. 生态学杂志, 2017, 36(3): 631–639. |
| [28] | 张雪萍, 李春艳, 殷秀琴, 陈鹏. 不同使用方式林地的土壤动物与土壤营养元素的关系. 应用与环境生物学报, 1999, 5(1): 26–31. |
| [29] | 林英华, 黄庆海, 刘骅, 彭畅, 朱平, 张树清, 张夫道. 长期耕作与长期定位施肥对农田土壤动物群落多样性的影响. 中国农业科学, 2010, 43(11): 2261–2269. DOI:10.3864/j.issn.0578-1752.2010.11.009 |
| [30] | 陈智裕, 马静, 赖华燕, 马祥庆, 吴鹏飞. 植物根系对根际微环境扰动机制研究进展. 生态学杂志, 2017, 36(2): 524–529. |
| [31] | 马克平. 生物群落多样性测度的方法——Ⅰα多样性的测度方法(上). 生物多样性, 1994, 2(3): 162–168. DOI:10.17520/biods.1994027 |
| [32] | Ferris H, Tuomisto H. Unearthing the role of biological diversity in soil health. Soil Biology and Biochemistry, 2015, 85: 101–109. DOI:10.1016/j.soilbio.2015.02.037 |
| [33] | 马克平, 刘灿然, 刘玉明. 生物群落多样性的测度方法——Ⅱβ多样性的测度方法. 生物多样性, 1995, 3(1): 38–43. |
| [34] | Carpenter S R, Kitchell J F, Hodgson J R. Cascading trophic interactions and lake productivity:fish predation and herbivory can regulate lake ecosystems. BioScience, 1985, 35(10): 634–639. DOI:10.2307/1309989 |
| [35] | McQueen D J, Post J R, Mills E L. Trophic relationships in freshwater pelagic ecosystems. Canadian Journal of Fisheries and Aquatic Sciences, 1986, 43(8): 1571–1581. DOI:10.1139/f86-195 |
| [36] | A'Bear A D, Johnson S N, Jones T H. Putting the 'upstairs-downstairs' into ecosystem service:what can aboveground-belowground ecology tell us?. Biological Control, 2014, 75: 97–107. DOI:10.1016/j.biocontrol.2013.10.004 |
| [37] | Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629–1633. DOI:10.1126/science.1094875 |
| [38] | 陈小云, 刘满强, 胡锋, 毛小芳, 李辉信. 根际微型土壤动物-原生动物和线虫的生态功能. 生态学报, 2007, 27(8): 3132–3143. |
| [39] | Lucas G B. Diseases of Tobacco.3rd ed. Raleigh, NC: Biological Consulting Associates, 1975. |
| [40] | Dromph K M, Borgen A. Reduction of viability of soil borne inoculum of common bunt (Tilletia tritici) by collembolans. Soil Biology and Biochemistry, 2001, 33(12/13): 1791–1795. |