 2017, Vol. 37
2017, Vol. 37文章信息
- 刘玮, 徐梦珍, 王兆印, 于丹丹, 周雄冬
- LIU Wei, XU Mengzhen, WANG Zhaoyin, YU Dandan, ZHOU Xiongdong.
- 沼蛤 (Limnoperna fortunei) 幼虫的附着行为特性
- Experimental study of attachment characteristics of golden mussel larvae
- 生态学报. 2017, 37(8): 2779-2787
- Acta Ecologica Sinica. 2017, 37(8): 2779-2787
- http://dx.doi.org/10.5846/stxb201601030010
-
文章历史
- 收稿日期: 2016-01-03
- 网络出版日期: 2016-10-29

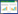




2. 河海大学水文水资源与水利工程科学国家重点试验室, 南京 210098
2. State Key Laboratory of Hydrology-Water Resources and Hydraulic Engineering, Nanjing 210098, China
沼蛤 (Limnoperna fortunei Dunker, 1857), 俗称淡水壳菜、水瓜子, 原产于我国南方地区, 是一种对环境适应能力强的入侵性底栖动物, 现已经广泛入侵到我国及东南亚国家的淡水河湖、水库中[1], 在南美的阿根廷、巴西等国家也发生了比较严重的沼蛤入侵[2]。沼蛤成贝营固着性生活, 移动范围有限, 其种群的扩散主要是依靠幼虫随水流输移完成的[3-4]。其幼虫浮游在水中, 先后经历D型幼虫、前期壳顶幼虫、后期壳顶幼虫、踯行期4个时期, 踯行期幼虫逐渐进入底栖固着阶段, 并逐渐发育成成贝[4]。对自然环境, 沼蛤的侵入会迅速改变当地原有生物群落结构, 减少物种多样性, 导致物种的退化, 甚至灭绝[5]。对输水工程, 沼蛤的侵入会引起输水系统中的沼蛤生物污损, 降低输水效率, 增加能耗;造成混凝土保护层的脱落, 腐蚀管壁[6];对于冷却管等管径较小的管路还会造成堵塞, 引发生产停止和安全事故[7];此外对沼蛤的清除也会损害混凝土保护层[8]。沼蛤入侵的防治已经成为一个世界性难题, 一旦沼蛤成功侵入一个水体, 则很难有效清除它们, 因此阻止其入侵至关重要[9], 研究表明对沼蛤幼虫的控制是阻止沼蛤入侵的最有效手段之一[4]。
本研究基于安徽省滁州市的琅琊山抽水蓄能电站中沼蛤生物污损的工程背景。该电站以日调节方式承担安徽电网调峰填谷和事故备用任务。电站主要设施包括上库、下库和抽水发电机组以及相关的水道工程, 下库是滁州市的饮用水源[10]。由于沼蛤幼虫个体微小, 随水流自由迁移, 进入该电站的直径只有4 cm的冷却水管系统造成堵塞, 影响电站的安全运行。本文着眼于研究沼蛤幼虫的附着行为特性, 并利用该特性对原水中的幼虫进行吸引附着处理, 以减少沼蛤幼虫进入工程造成污损附着。前期研究表明, 沼蛤对不同材质的附着基具有不同的选择性, 本研究中选用对幼虫吸附性良好的黄麻材料来进行附着试验[11-12], 研究幼虫的附着行为表现与附着材料的长度、水流流量等的关系, 为设计和优化吸引沼蛤幼虫附着、减少其进入冷却水系统的装置提供参考。
1 研究方法 1.1 实验布置试验池建立在琅琊山抽水蓄能电站下库库区, 通过两台潜水泵为整个试验池供水, 试验池结构与采样点设置如图 1a所示。为了控制进口流量和水流掺混, 两台潜水泵分别在两个进水口处供水, 在前池稳定水流后, 再经过进口堰流遍整个附着池, 最终从排水口直接排出。进口堰、排水堰与排水口结构如图 1b和图 1c。在附着池内安放附着排, 在不同位置设置采样点, 图 1a中1—6即设置的采样点, 在这些样点处采集水样, 检测其中的幼虫密度, 代表此时该断面处的幼虫密度。其中样点1的幼虫密度指示原水中的幼虫密度, 样点2的幼虫密度指示原水经过1 m附着材料吸附后的幼虫密度, 其它采样点的意义可依此类推。试验采用黄麻材质制作附着排, 尺寸为65 cm×100 cm (图 1d)。将附着排按照图 1a中所示的方式布置在附着池内, 每一排材料称为一段, 因此在试验中, 材料长度 (m)=材料段数×1。布置材料时, 原则上是在不堵水的前提下, 每段布置附着排越多越好, 在本试验中, 多次试水调整后每段安放15块附着排。

|
| 图 1 附着池结构及采样点设置示意图 Fig. 1 Structure sketch of experimental flume and sampling sites |
(1) 采样与数据处理 采用孔径48 μm的300目浮游生物网在图 1a中各采样点分别采集水样0.12 m3, 收集浮游生物网过滤的滤液约15 mL, 滤液装入50 mL样品管中。将样品管放在试管架上静置, 待沼蛤幼虫充分沉降后, 用吸管缓慢吸出上清液直至剩余2.5 mL底液。将全部底液分批加在1 mL浮游生物计数框中, 在光学显微镜 (上海光学仪器一厂XSP-44X.9) 下检查[13], 通过SmartV Camera图像采集系统对幼虫进行显微成像, 同时分别统计幼虫的死亡个体、D型幼虫、前期壳顶幼虫、后期壳顶幼虫、踯行期个体的数量, 后期可利用MIVNT图像分析软件获取幼虫尺寸[4]。在水体中的所有沼蛤幼虫的密度, 包括所有死亡个体和活体, 称为幼虫的总体密度。其中, 活体幼虫包括全部活体的D型幼虫、前期壳顶幼虫、后期壳顶幼虫、踯行期个体。理论上, 从采样开始时到检查结束时, 采集的样本是不应该发生任何变化, 但在实际试验过程中, 随着试验的进行, 幼虫的活体密度略有降低。因治理主要针对的是活体幼虫, 为了提高分析结果的可靠性, 分析时综合考虑幼虫的总体密度和活体密度这两个指标。
(2) 独立附着试验 控制单一变量, 即控制附着材料长度不变, 以试验池进水流量3.1, 5.7, 6.7, 9.6, 12.1 L/s分别来进行试验;或控制流量不变, 以附着材料的长度1, 3, 5, 6, 8 m分别来进行试验, 研究附着试验在不同流量, 不同附着材料长度下的不同表现。如在进行附着材料长度为3 m的独立试验时, 即在如图 1中样点1和样点3处采样, 暂时去除样点3下游的附着排, 如此即可排除下游附着排的影响。在这些不同的条件下分别进行独立附着试验并记录试验结果, 每个工况下的独立附着试验至少重复3次以排除随机误差的影响, 根据附着材料上下游幼虫密度的变化来分析吸附效果。附着材料的吸附率定义为水体经过试验段后, 幼虫密度降低的百分比:
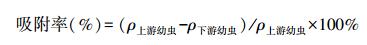
|
(1) |
(3) 沿程分布试验 为了分析吸附排对幼虫吸附的沿程效果, 将附着材料长度设置为最长 (8 m), 然后逐段进行取样, 即依次图 1a中所指示的1, 2, 3, 4, 6这5个样点处的水样, 据此分析幼虫密度在附着材料中的沿程变化。沿程分布试验的工况安排是在流量为5.7 L/s时进行3次试验, 然后再在流量为3.1、6.7 L/s分别进行2次、1次试验以作验证。
(4) 附着材料布置 在材料数量相同的前提下, 简单测试附着排布置密集程度对吸附效果的影响。将8段的附着排的最后4段均匀打散排列为8段, 以将长度为8 m (密集布置) 的附着材料区域延伸为12 m (稀疏布置)。采集原水中及经过所有附着材料后的水流中的水样, 分析比较相同数量的材料密集布置和疏松布置对吸附效果的影响。在进水流量3.1, 6.7, 9.6, 12.1 L/s这4个流量工况下各进行6组重复试验。
(5) 附着的不稳定性 为了验证幼虫, 尤其是早期阶段的幼虫在材料上的附着具有不稳定的特性, 特根据现场条件采用幼虫密度几乎为0的原水流过已吸附了幼虫的附着材料, 来进行其他条件与独立附着试验相同的“冲刷试验”, 观察“冲刷”后下游幼虫的分布情况。对原水及附着材料上、下游的水流进行采样, 分析幼虫密度极低的水流经过附着材料后水体中幼虫密度的变化。本试验进行3次重复试验。
2 结果及讨论 2.1 黄麻材料长度和水流流量对沼蛤幼虫附着的影响分别控制流量和吸附材料长度的条件下, 分析经过吸附排吸附作用后上下游总体密度和活体密度的变化 (图 2), 结果一致表明:(1) 相同试验流量下, 不同长度的附着材料对幼虫的吸附效果不同, 材料长度与吸附效果关系的总体趋势是:在同一流量下, 随着吸附材料的延长, 上下游幼虫密度降低也明显, 即吸附效果明显提高。具体表现为:材料长度为1 m时吸附效果较差 (上下游密度平均值相当, 下游密度平均值较上游降低31%), 3—5 m时效果尚可 (下游密度平均值较上游降低55%—56%左右), 6—8 m效果较好 (下游密度平均值较上游降低62%—76%, 最佳时达到93%)。(2) 而在相同的材料长度情况下, 流量对吸附效果也有一定的影响, 当流量过大时 (在本试验中为12.1 L/s), 幼虫密度最高吸附率和平均吸附率都普遍低于其他低流量组。说明当吸附材料长度不足或吸附池流量过高流速过快时, 幼虫还未来得及吸附到材料上就被水流带出附着区域, 或者吸附十分不稳定极易从吸附材料上脱落导致吸附失效。徐梦珍等[11-12]的研究也认为沼蛤在粗布上的早期附着是不稳定的。

|
| 图 2 不同附着材料长度及不同流量下的吸附效果 Fig. 2 Attachment efficiency under the different attaching-plate lengths and discharges 1m-U:1m-上游1m-Upstream;1m-D:1m-下游1m-Downstream, 其它依此类推; Length:附着材料的长度length of attachment material |
沼蛤幼虫的发育经历变态过程, 不同发育阶段的幼虫具有不同的特点, Dos Santos、Darrigran、中国科学院水生生物研究所管道小组等学者对沼蛤幼虫的发育阶段有不一致的划分[8]。于2010—2012年在广东省惠州市西枝江, 2014—2015年在安徽省滁州市琅琊山抽水蓄能电站下库, 长期观测原水中沼蛤幼虫的分布, 观测结果与Dos Santos[14]的研究基本一致。原水中沼蛤幼虫的发育主要分为4个阶段:D型幼虫期、前期壳顶幼虫期、后期壳顶幼虫期、踯行期[12-14]。幼虫发育到后期壳顶幼虫期结束需要30d左右, 其中后期壳顶幼虫期约持续10d[15]。各阶段结构如图 3所示, 其中D型幼虫期、前期壳顶幼虫以面盘为游泳器官, 营游泳生活, 为浮游型幼虫, 附着性较差;后期壳顶幼虫、踯行期幼虫面盘逐渐退化, 开始以足做爬行运动[16], 为底栖型幼虫, 已初步具备主动附着能力, 向固着阶段发育, 因此附着能力要较前期壳顶幼虫阶段更强[17]。本文定义水样中底栖型幼虫在所有活体幼虫中所占的比例为底栖幼虫比 (benthic larvae ratio), 公式为:

|
| 图 3 不同发育阶段的沼蛤幼虫及其体长 Fig. 3 Length of the golden mussel larvae in the different development stages |
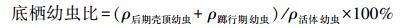
|
(2) |
以附着材料上下游水体中底栖幼虫比为指标, 统计不同条件下经过附着试验后, 上下游水体中幼虫发育阶段组成的变化, 据此分析不同幼虫发育阶段的附着表现, 结果如图 4。图 4中每个虚线隔开的区域为一组相同流量不同材料长度情况下的试验结果:每个流量下5个材料长度梯度, 图中显示的数据是至少3组重复试验的底栖幼虫比平均值。各种试验条件下, 附着试验的整体趋势是:经过附着材料后, 水体中底栖幼虫比明显降低, 即水体中后期壳顶幼虫及踯行期幼虫的比例降低, 说明后期壳顶幼虫及踯行期幼虫在材料上发生了附着, 且相较于浮游型幼虫底栖型幼虫更容易发生附着。随着附着排长度延长, 底栖幼虫比降低, 甚至趋近于0, 如试验组“流量3.1 L/s, 材料长度3 m”、“流量3.1 L/s, 材料长度6 m”、“流量5.7 L/s, 材料长度6 m”、“流量5.7 L/s, 材料长度8 m”等工况下, 下游底栖幼虫比为0, 下游没有底栖型幼虫, 即效果最好时水体中的后期壳顶幼虫及踯行期幼虫基本都附着在吸附材料上。需指出的是流量过大会抑制幼虫的吸附效果, 如流量为12.1 L/s时在各组试验中吸附效果均较差, 而在9.6 L/s时相对较好, 因此在本试验条件下, 流量不宜超过10 L/s。

|
| 图 4 附着材料长度对幼虫发育阶段组成的影响 Fig. 4 Influence of attaching-plate length on the composition of the different larval stages |
如图 5, 幼虫密度整体上随着附着材料的延长而逐渐降低, 即附着材料越长, 附着效果越好, 这点与之前的独立附着试验结果一致。结合图 4、图 5可知, 由于不同发育阶段的幼虫在附着材料上具有不同的吸附表现, 后期壳顶幼虫吸附效果比前期壳顶幼虫的吸附效果要好, 吸附也更稳定。在图 5中的这几种情况下, 通过8 m附着材料后, 下游水中后期壳顶幼虫的密度都趋近于0, 从而进一步证明了附着材料能有效降低水中的底栖幼虫比。从幼虫密度在整个附着材料段的沿程变化可以看出, 在密度整体降低的趋势下, 密度曲线有一定的波动, 尤其是在附着材料长度较小时 (如材料为1—2m时), 这可能与有些幼虫附着不稳定发生脱落有关。在图 5中出现的这种附着不稳定时幼虫密度的波动, 也解释了图 2中出现的下游幼虫密度比上游高的现象。

|
| 图 5 幼虫密度在附着实验中的沿程分布 Fig. 5 The variation of golden mussel larval density along distance Total, 全部幼虫total larvae;Alive, 活体幼虫alive larvae;Pre-stage, 前期壳顶幼虫Umbonal veligers;Post-stage, 后期壳顶幼虫Pediveligers |
调整“8 m密集布置试验组”为“12 m稀疏布置试验组”的结果如图 6。不论是从上下游总体密度还是活体密度变化来分析, 在各种流量下, 当附着排的总数量一定时, 将附着材料“稀疏布置”的试验效果比“密集布置”的试验效果要差很多。材料摆放成12 m时, 材料之间空隙较大, 使得大量水流直接从材料两旁的水道流走, 而没有与材料发生接触, 从而减小了幼虫向材料发生附着的机会[11]。因此, 附着试验的效果并不仅仅取决于附着材料的长度, 断面上幼虫接触附着材料的机会也十分重要。因此在安装附着材料时需要注意:在达到过水能力要求的前提下, 断面上附着材料的安放密度越高越好。

|
| 图 6 材料长度为8 m与12 m时吸附效果的对比 Fig. 6 Comparison of attachment efficiency for the attaching-plate length of 8 m and 12 m |
冲刷试验结果如图 7所示, 当上游的幼虫密度较低时, 通过试验材料后, 水体中幼虫密度不降反增, 甚至提高了一个量级。由于原水中幼虫密度是极低的, 因此下游密度的增高是因为附着材料上吸附的部分幼虫脱落下来, 进入水体。由此可见, 幼虫在附着材料上的附着处于一种“动态过程”中, 即幼虫附着到材料表面后, 不一定一直附着在材料上, 可能会在水流作用等原因下脱落下来[11], 因此吸附率难以达到100%。结合附着试验对幼虫发育阶段组成的影响可知, 通过附着材料后, 相比于浮游型幼虫, 附着的底栖型幼虫脱落下来的较少。

|
| 图 7 冲刷试验上下游的幼虫密度对比 Fig. 7 Comparison of the larvae density of upstream and downstream in the flushing experiment |
此外, 附着材料的吸附效果具有一定的有效期, 随着黄麻布在水中的浸泡, 其吸附效果有下降的趋势。原因可能有以下几点:其一, 黄麻布在浸泡的过程中, 由于水压的关系, 原本蓬松的材质被压紧, 导致相同数量的布排, 在浸泡的过程中, 彼此间隔逐渐扩大, 造成幼虫接触到附着排的机会降低。其二, 黄麻布在浸泡的过程中, 黄麻表面的孔隙被水中的浮泥粘粒填满, 造成麻布表面十分光滑, 不利于沼蛤幼虫的附着[18]。其三, 水压与浮泥粘粒的共同作用也使包裹在铁支架上的两块布片彼此粘连, 造成了与水流的接触面明显减小, 并进一步加大了附着排之间的空隙, 在检查布排时都可以发现这些情况。这点与作者团队在西枝江进行的沼蛤成贝的附着特性研究一致[12]。在本实验的条件下, 由于针对的是幼虫的附着特性, 附着材料的有效期相比控制成贝时, 有效期较长, 约2周。
3 结论通过以上试验对沼蛤幼虫的附着特性进行研究, 作者认为有效提高材料吸附效果、减少水中幼虫密度的关键因素是保证足够的附着材料长度和控制合适的流量。同时对吸附材料的安装和使用提出了一些建议。结果总结如下:结合独立试验及沿程分布试验的结果可知, 同一流量下, 不同长度的附着材料对幼虫的吸附效果不同, 总体上, 随着吸附材料的延长, 吸附效果越好, 上下游幼虫密度和底栖幼虫比降低越明显。附着材料长度达到6—8 m, 吸附效果较好, 下游密度平均值较上游降低62%—76%, 最佳时达到93%。在相同的材料长度情况下, 流量对吸附效果也有一定的影响, 当流量过大时, 吸附效果会降低。
而在实际工程运行情况下, 水流中的幼虫只会随水流在工程设备中停留数小时便随水流流出, 因此其中的浮游型幼虫来不及发育为初步具备附着能力的底栖型幼虫, 因而相比于原水中的底栖型幼虫, 浮游型幼虫对工程造成的威胁较小。本文试验表明合理布置的吸附排能够有效吸附水中的底栖型幼虫, 降低进入工程的底栖型幼虫密度, 从而降低工程设备遭受沼蛤污损附着的威胁。最后, 在设计吸附设备时, 首先应注意, 在达到工程输水量的前提下, 附着排宜密集布置; 其次, 考虑材料的有效期, 附加排在使用一段时间后需及时清理和更换, 也避免已附着的幼虫脱落而造成二次入侵。
| [1] | Xu M Z. Distribution and spread of Limnoperna fortunei in China//Boltovskoy D, ed. Limnoperna Fortunei. Switzerland: Springer International Publishing, 2015: 313-320. |
| [2] | Darrigran G, Damborenea C, Greco N. An evaluation pattern for antimacrofouling procedures:Limnoperna fortunei larvae study in a hydroelectric power plant in South America. Ambio, 2007, 36(7): 1575–579. |
| [3] | Ricciardi A. Global range expansion of the Asian mussel Limnoperna fortunei (Mytilidae): another fouling threat to freshwater systems. Biofouling: The Journal of Bioadhesion and Biofilm Research, 1998, 13(2): 97–106. DOI:10.1080/08927019809378374 |
| [4] | 徐梦珍, 王兆印, 王旭昭, 刘玮. 输水通道中沼蛤入侵及水力学防治. 水利学报, 2013, 44(7): 856–862. |
| [5] | Martín S M, Darrigran G. Limnoperna fortunei (Dunker 1857) en el balneario Bagliardi, Río de la Plata: Alteración en la composición de la malacofauna litoral. Tankay, 1994, 1: 164–166. |
| [6] | 叶宝民, 曹小武, 徐梦珍, 王兆印, 林正傑. 沼蛤对长距离输水工程入侵调查研究. 给水排水, 2011, 37(7): 99–103. |
| [7] | 娄康后, 刘健. 贻贝堵塞管道的防除研究. 海洋与湖沼, 1958, 1(3): 316–324. |
| [8] | 徐梦珍. 底栖动物沼蛤对输水通道的入侵及防治试验研究[D]. 北京: 清华大学, 2012. |
| [9] | Simberloff D. Eradication of island invasives: practical actions and results achieved. Trends in Ecology & Evolution, 2001, 16(6): 273–274. |
| [10] | 唐书童. 琅琊山抽水蓄能电站及上下库的调度运行. 江淮水利科技, 2009(4): 20–23. |
| [11] | 徐梦珍, 曹小武, 王兆印, 王旭昭. 输水工程中沼蛤的附着特性. 清华大学学报, 2012, 52(2): 170–176. |
| [12] | Xu M Z, Darrigran G, Wang Z Y, Zhao N, Lin C C, Pan B Z. Experimental study on control of Limnoperna fortunei biofouling in water transfer tunnels. Journal of Hydro-Environment Research, 2015, 9(2): 248–258. DOI:10.1016/j.jher.2014.06.006 |
| [13] | 李洪武, 宋培学. 海洋浮游生物学. 合肥:中国科学技术大学出版社, 2012: 193–194. |
| [14] | Dos Santos C P, Würdig N L, Mansur M C D. Fases larvais do mexilhão dourado Limnoperna fortunei (Dunker) (Mollusca, Bivalvia, Mytilidae) na bacia do Guaíba, Rio Grande do Sul, Brasil. Revista Brasileira de Zoologia, 2005, 22(3): 702–708. DOI:10.1590/S0101-81752005000300029 |
| [15] | Cataldo D, Boltovskoy D, Hermosa J L, Canzi C. Temperature-dependent rates of larval development in Limnoperna fortune (bivalvia: mytilidae). Journal of Molluscan Studies, 2005, 71(1): 41–46. DOI:10.1093/mollus/eyi005 |
| [16] | 吴宝玲. 贝类繁殖附着变态生物学. 济南: 山东科学技术出版社, 1999: 98–99. |
| [17] | 王如才, 王昭萍. 海水贝类养殖学. 青岛: 中国海洋大学出版社, 2008: 37–37. |
| [18] | 曹小武, 徐梦珍, 王兆印, 王旭昭, 刘明祥. 泥沙落淤对输水工程中淡水壳菜污损的抑制作用. 中国农村水利水电, 2014(8): 45–48. |