 2017, Vol. 37
2017, Vol. 37文章信息
- 吴龙飞, 姜文虎, 刘洋, 李忠玲, 刘军侠
- WU Longfei, JIANG Wenhu, LIU Yang, LI Zhongling, LIU Junxia.
- 不同林分类型对昆虫群落多样性的影响
- Effects of different forest stand types on diversity of insect community
- 生态学报. 2017, 37(7): 2217-2224
- Acta Ecologica Sinica. 2017, 37(7): 2217-2224
- http://dx.doi.org/10.5846/stxb201512172518
-
文章历史
- 收稿日期: 2015-12-17
- 网络出版日期: 2016-08-30





2. 河北省林木种质资源与森林保护重点实验室, 保定 071000
2. Hebei Key Lab of Forest Germplasm Resources and Protection, Baoding 071000, China
一直以来植物与昆虫的相互作用都是生态学者们研究的热点, 同时与植物生长有关的各项条件 (如种类、混交形式、郁闭度、林龄、森林经营措施等) 的变化对昆虫群落多样性的影响亦受到广泛关注[1], 如研究不同树种混交及混交比例对昆虫多样性的影响, 表明植被多样性高的林分内昆虫群落多样性亦较高, 且不同混交比例对昆虫种群的分布有明显影响[2-3]。其中, 许多学者针对不同地域、不同类型的天然林进行了昆虫多样性研究[4-5], 但对于近自然森林经营下的人工林内的昆虫群落结构研究涉及很少。近自然森林经营是一种顺应自然的计划和管理森林的模式, 是充分利用森林生态系统内部的自然生长发育规律, 在森林自然更新的基础上计划和设计各项森林经营活动, 优化森林的结构和功能, 从而使生态与经济的需求能最佳结合的一种接近自然地森林经营模式[6-7]。
我国是世界上人工林面积最大的国家, 不少学者对我国不同类型的人工林现状及经营模式进行了分析探讨, 普遍认为我国人工林缺乏科学和有效的经营管理模式, 人工林生物样多样性下降, 生态效益和经济效益低。塞罕坝自然保护区内是以华北落叶松和樟子松为主要树种的人工林, 在保护区采取近自然森林经营措施, 严格控制外界人为因素的干扰, 适时进行林木抚育, 改善林木生长环境, 达到了一种近自然森林的状态。与保护区外的人工林相比保护区内松毛虫等重大森林害虫很少发生, 即使发生也不成灾, 具有良好的生态效益和经济效益。因此本文从昆虫生态学角度出发, 为探究该保护区内这种近自然森林经营下的森林生态系统本身的自然控害作用, 以华北落叶松纯林和华北落叶松不同类型混交林为研究对象, 探讨不同林分类型中昆虫群落结构及其变化规律[3], 明确保护区不同林分类型对昆虫群落的影响以及对主要害虫的控害机制, 为华北落叶松人工林的重大害虫防控和经营管理提供参考。
1 研究地区和研究方法 1.1 研究区域概况塞罕坝自然保护区是2002年经河北省人民政府批准建立的森林生态系统保护区, 该保护区 (42°22′—42°31′N, 116°53′—117°31′E) 位于河北省承德市围场满族蒙古族自治县 (围场镇) 境内, 与内蒙古高原相邻, 总面积20029.8 hm2, 森林覆盖率80.74%, 以华北落叶松和樟子松为主要人工林, 两者均占保护区总面积的8%[8]。保护区属温带半湿润季风气候区, 全年气候特征为:冬季漫长, 低温寒冷;春季干燥多风;夏季不明显, 光照强;降水量偏少。保护区内植物资源丰富, 有森林、灌丛、灌草丛、草原、草甸及沼泽植被等多种植被类型[9]。乔木以华北落叶松、樟子松、白桦为主要树种, 其中华北落叶松混交林以落叶松-樟子松、落叶松-白桦、落叶松-山刺玫等混交类型为主;灌丛主要有山刺玫灌丛、金露梅灌丛、山荆子灌丛、虎榛子灌丛、绣线菊灌丛等。
1.2 研究方法 1.2.1 样地设置根据保护区内常见的植被类型选择了6种不同林分类型的华北落叶松林, 林龄为35—40 a, 林分密度约为600—750株/hm2, 且华北落叶松与其他树种的混交比例为1:1。样地大小设置为20 m×20 m, 每种林分类型设3个重复, 样地概况见表 1。
| 样地 Plot |
林分类型 Forest stand type |
重复 Repetition |
海拔 Altitude/m |
坡度 Slope/(°) |
坡向 Aspect |
坡位 Slope location |
林下植被盖度 Forest vegetation coverage |
| Ⅰ | 华北落叶松 | 1 | 1448 | 2 | 阳 | 中 | 0.6 |
| Prince Rupprecht′s larch | 2 | 1477 | 7 | 阳 | 下 | 0.5 | |
| 3 | 1401 | 2 | 阳 | 下 | 0.6 | ||
| Ⅱ | 华北落叶松-樟子松 | 1 | 1428 | 1 | 阳 | 中 | 0.7 |
| Prince Rupprecht′s larch-Mongolian scots pine | 2 | 1414 | 3 | 阳 | 下 | 0.6 | |
| 3 | 1466 | 10 | 阳 | 上 | 0.6 | ||
| Ⅲ | 华北落叶松-白桦 | 1 | 1450 | 2 | 阳 | 下 | 0.4 |
| Prince Rupprecht′s larch-White Birch | 2 | 1561 | 8 | 阳 | 上 | 0.5 | |
| 3 | 1503 | 6 | 阳 | 上 | 0.4 | ||
| Ⅳ | 华北落叶松-山刺玫 | 1 | 1501 | 2 | 阳 | 上 | 0.4 |
| Prince Rupprecht′s larch-Rosa davurica Pall | 2 | 1470 | 6 | 阳 | 中 | 0.3 | |
| 3 | 1518 | 1 | 阳 | 下 | 0.5 | ||
| Ⅴ | 华北落叶松-樟子松-山刺玫 | 1 | 1477 | 7 | 阳 | 中 | 0.5 |
| Prince Rupprecht′s larch-Mongolian scots | 2 | 1423 | 5 | 阳 | 下 | 0.6 | |
| pine-Rosa davurica Pall | 3 | 1430 | 1 | 阳 | 下 | 0.4 | |
| Ⅵ | 华北落叶松-樟子松-白桦-山刺玫-山荆子 | 1 | 1481 | 11 | 阳 | 中 | 0.5 |
| Prince Rupprecht′s larch-Mongolian scots pine-White | 2 | 1474 | 13 | 阳 | 上 | 0.3 | |
| Birch-Rosa davurica Pall-Malus baccata | 3 | 1477 | 12 | 阳 | 中 | 0.6 |
调查采集于2014.6—2014.9进行。使用扫网法在样地内采样, 用捕虫网 (网口直径30 cm, 深50 cm, 锥形, 以白色尼纶纱制成) 在样方的各个方位随机扫取100网 (共计200下), 将采集的标本装入毒瓶中杀死, 然后放入三角纸袋内, 记录好时间、地点带回实验室鉴定。昆虫标本至少鉴定到科, 未能鉴定到种的昆虫标本将其编号作为单独的种进行处理[10]。
1.2.3 数据分析方法本文使用Excel、Forstat和SPSS对数据进行统计和分析。
(1) 群落特征指数测定
丰富度指数 (Richness Index)
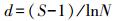
|
式中, S为物种数;N为群落中总的个数。
Shannon-Wiener多样性指数 (Diversity Index)
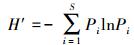
|
式中, S为物种数;Pi=Ni/N;Ni为第i种的个体数;N为群落中总的个数。
Pielou均匀度指数 (Evenness Index)
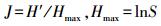
|
式中, H′为Shannon-Wiener多样性指数;Hmax为给定物种完全均匀时的群落多样性指数;S为物种数。
Berger-Parker优势度指数 (Dominance Index)
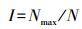
|
式中, Nmax为优势类群的数量;N为群落中总的个数[11]。
(2) 群落相似性测定
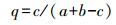
|
式中, c为两样地相同的物种数;a、b分别为两样地各自物种数[12]。
该公式规定相似性等级是:当q=0.00—0.25时, 极不相似;当q=0.25—0.50时, 中等不相似;当q=0.50—0.75时, 中等相似;当q=0.75—1.00时, 极相似。
(3) 群落的主成分分析
以昆虫不同类群的物种数和个体数为因子[13], 应用SPSS软件进行主成分分析。M1为捕食性类群的物种数, M2为捕食性类群的个体数, M3为植食性类群的物种数, M4为植食性类群的个体数, M5为寄生性类群的物种数, M6为寄生性类群的个体数, M7为中性类群的物种数, M8为中性类群的个体数。
(4) 群落相对稳定性测定
采用群落物种数与个体数之比 (St/Si) 和天敌类群物种数与植食性类群物种数之比(Sn/Sp) 表示; St/Si反映种间数量上的制约作用, Sn/Sp反映食物网关系和天敌—害虫相互制约的复杂程度[14]。
2 结果与分析 2.1 群落组成结构变化本次共获得标本9542头, 隶属于7目 (分别为双翅目、半翅目、鞘翅目、膜翅目、直翅目、鳞翅目、脉翅目), 68科, 187种。
如表 2所示, 从目数来看, 样地Ⅴ、Ⅵ内昆虫目数7个最多, 样地Ⅱ、Ⅳ次之, 样地Ⅲ昆虫目数最少。从科数和物种数上看, 样地Ⅱ、Ⅰ和Ⅵ的科数和物种数均较多, 主要优势类群包括双翅目的蝇科、寄蝇科、舞虻科、花蝇科, 半翅目的叶蝉科、沫蝉科、姬猎蝽科、盲蝽科, 鞘翅目的叶甲科、肖叶甲科、瓢甲科等;主要常见的物种有家蝇 (Musca domestica)、麻蝇 (Sarcophaga naemorrhoidalis Fallen)、大青叶蝉 (Cicadella viridis)、松沫蝉 (Aphrophora flavipes Uhler)、窄姬猎蝽 (Nabis stenoferus Hsiao)、四点苜蓿盲蝽 (Adelphocoris suturalis)、斑额隐头叶甲 (Cryptocephalus kulibini Gebler)、蒿金叶甲 (Chrysolina aurichalcea)、七星瓢虫 (Coccinella septempunctata) 等。样地Ⅲ的物种数最少, 主要为叶蝉科、花蝇科、大蚊科、寄蝇科物种。从个体数上看, 样地Ⅱ内昆虫个体数最多, 样地Ⅲ和样地Ⅴ内昆虫个体数最少。
| 样地 Plot |
目数 Order |
科数 Families |
所占比例 Percent/% |
物种数 Species |
所占比例 Percent/% |
个体数 Individual number |
所占比例 Percent/% |
| Ⅰ | 5 | 43 | 63.24 | 91 | 48.66 | 2134 | 22.36 |
| Ⅱ | 6 | 45 | 66.18 | 90 | 48.13 | 3210 | 33.64 |
| Ⅲ | 4 | 27 | 39.71 | 52 | 27.81 | 603 | 6.32 |
| Ⅳ | 6 | 36 | 52.94 | 65 | 34.76 | 1014 | 10.63 |
| Ⅴ | 7 | 36 | 52.94 | 60 | 32.09 | 644 | 6.75 |
| Ⅵ | 7 | 41 | 60.29 | 86 | 45.99 | 1937 | 20.30 |
类群的划分可以直观的了解昆虫群落结构的组成情况, 由表 3可知, 植食性类群在6种林分内的物种数和个体数均最多, 捕食性类群和寄生性类群两者的物种数与植食性类群物种数相当, 中性类群的物种数和个体数均最少。总体可以看出, 6种林分类型内的天敌类群具有一定优势地位, 在某种程度上可以控制植食性类群部分害虫的大发生, 使昆虫群落结构具有一定的稳定性。
| 样地 Plot |
捕食性类群Predatory group | 植食性类群Herbivorous group | 寄生性类群Parasitic group | 中性类群Neutral group | |||||||
| S | I | S | I | S | I | S | I | ||||
| Ⅰ | 20 | 513 | 47 | 1147 | 20 | 227 | 4 | 247 | |||
| Ⅱ | 22 | 487 | 43 | 2341 | 18 | 267 | 7 | 115 | |||
| Ⅲ | 7 | 76 | 21 | 332 | 15 | 114 | 9 | 81 | |||
| Ⅳ | 14 | 220 | 33 | 315 | 12 | 432 | 6 | 47 | |||
| Ⅴ | 13 | 219 | 39 | 371 | 5 | 31 | 3 | 23 | |||
| Ⅵ | 28 | 446 | 40 | 1286 | 13 | 118 | 5 | 87 | |||
| S:物种数Number of species;I:个体数Number of individual | |||||||||||
由图 1可知, 0.05显著水平下, 从丰富度指数上看, 样地Ⅱ的值最高, 样地Ⅲ的值最低, 且样地Ⅲ与样地Ⅰ、样地Ⅱ存在显著差异, 与其它样地之间差异不明显。推断原因可能是落叶松纯林和落叶松-樟子松混交林下植被盖度较高, 物种多样性、丰富度亦较高 (以细叶苔草、地榆、朝天萎陵菜、歪头菜、唐松草、羊茅等为优势物种), 增加了昆虫取食和生存的机会, 使昆虫群落的丰富度和优势度指数较高, 均匀度指数相应较低, 而落叶松-白桦混交林下植被盖度虽高, 但物种比较单一 (主要为草地早熟禾、蚊子草、草原老鹳草), 昆虫群落的丰富度指数低。

|
| 图 1 不同林分类型昆虫群落特征指数 Fig. 1 Characteristic index of insect communities for different forest stand types 图中字母表示在0.05水平下的显著性; Ⅰ:华北落叶松,Prince Rupprecht′s larch;Ⅱ:华北落叶松—樟子松,Prince Rupprecht′s larch-Mongolian scots pine;Ⅲ:华北落叶松—白桦,Prince Rupprecht′s larch-White Birch;Ⅳ:华北落叶松—山刺玫,Prince Rupprecht′s larch-Rosa davurica Pall;Ⅴ:华北落叶松—樟子松—山刺玫,Prince Rupprecht′s larch-Mongolian scots pine-Rosa davurica Pall;Ⅵ:华北落叶松—樟子松—白桦—山刺玫—山荆子,Prince Rupprecht′s larch-Mongolian scots pine-White Birch-Rosa davurica Pall-Malus baccata |
从多样性指数上看, 6种林分类型之间不存在显著差异, 即使是纯林和林分结构简单的针叶混交林也表现为较高的多样性指数, 可能由于在近自然森林经营模式下, 保护区内的森林达到了一种近自然林的状态, 逐渐形成了较为复杂且稳定的森林生态系统, 加上落叶松林下植被物种丰富, 增加了昆虫群落多样性。从均匀度指数上看, 样地Ⅲ的值最高, 且样地Ⅱ与样地Ⅲ、样地Ⅴ之间存在明显差异, 其它林分间差异不明显, 表明样地Ⅲ内昆虫物种分布相对均衡, 没有突发性, 种间具有一定的制约能力。从优势度指数上看, 样地Ⅱ的优势度指数明显高于其它, 且样地Ⅱ与样地Ⅰ之间存在显著差异, 其它林分间不存在差异, 表明样地Ⅱ中优势类群较多 (主要有叶蝉科、盲蝽科、花蝇科、叶甲科等), 且优势类群的地位比较突出。
2.3 群落的相似性分析由表 4可知, 样地Ⅲ与样地Ⅴ、样地Ⅵ的相似性系数分别为0.2308、0.2432(q<0.25), 表示样地Ⅲ与样地Ⅴ、样地Ⅵ内昆虫群落结构极不相似。其它林分类型之间的q值均大于0.25小于0.50, 表示其它林分类型之间的昆虫群落均为中等不相似。由此可知, 6种林分类型之间的昆虫群落结构均不相似, 各自具有不同的昆虫群落组成特点, 表明不同林分类型内由于树种和混交形式的不同对昆虫物种组成有明显作用。
| 样地Plot | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ |
| Ⅰ | — | 0.3507 | 0.2768 | 0.2893 | 0.2797 | 0.3615 |
| Ⅱ | — | 0.2566 | 0.3136 | 0.3636 | 0.3750 | |
| Ⅲ | — | 0.3295 | 0.2308 | 0.2432 | ||
| Ⅳ | — | 0.3020 | 0.3246 | |||
| Ⅴ | — | 0.3519 | ||||
| Ⅵ | — |
主成分分析方法明确了不同林分类型内昆虫群落的主要构成因素以及各因素之间的相互关系, 结果见表 5。由表 5可知, 6种林分类型内均提取了2个主成分, 且累计贡献率均高达100%, 足以概括组成昆虫群落的主要因子[15]。
| 样地 Plot |
成分 Component |
因子载荷量Factor Loading | 累计贡献率 Cumulative/% |
|||||||
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | M8 | |||
| Ⅰ | 1 | -0.753 | 0.323 | -0.803 | -0.362 | 0.943 | 0.920 | 0.947 | 0.915 | 61.44 |
| 2 | -0.658 | 0.947 | 0.596 | 0.932 | 0.332 | -0.393 | -0.323 | 0.403 | 100.00 | |
| Ⅱ | 1 | -0.992 | 0.238 | 0.831 | 0.764 | 0.919 | 0.996 | 0.015 | 0.917 | 62.38 |
| 2 | 0.130 | 0.971 | 0.557 | -0.645 | -0.395 | -0.090 | 1.000 | 0.398 | 100.00 | |
| Ⅲ | 1 | -0.722 | 0.018 | 0.985 | 0.988 | -0.093 | 0.725 | 0.971 | 1.000 | 61.81 |
| 2 | 0.692 | -1.000 | 0.175 | 0.154 | 0.996 | 0.689 | -0.238 | 0.018 | 100.00 | |
| Ⅳ | 1 | -0.619 | 0.353 | -0.982 | 0.842 | -0.143 | -0.991 | 0.888 | 0.897 | 59.71 |
| 2 | 0.786 | 0.936 | 0.189 | -0.540 | 0.990 | -0.134 | 0.459 | 0.442 | 100.00 | |
| Ⅴ | 1 | 0.885 | 0.658 | 0.992 | 0.901 | 0.999 | 0.911 | 0.873 | 0.845 | 78.96 |
| 2 | 0.466 | 0.753 | -0.130 | -0.433 | -0.039 | -0.412 | -0.488 | 0.534 | 100.00 | |
| Ⅵ | 1 | -0.206 | -0.259 | -0.998 | -0.940 | 0.969 | 1.000 | 0.990 | 0.874 | 70.90 |
| 2 | 0.978 | 0.966 | 0.060 | 0.340 | 0.246 | 0.030 | 0.140 | 0.487 | 100.00 | |
| M1:捕食性类群的物种数;M2:捕食性类群的个体数;M3:植食性类群的物种数;M4:植食性类群的个体数;M5:寄生性类群的物种数;M6:寄生性类群的个体数;M7:中性类群的物种数;M8为中性类群的个体数 | ||||||||||
样地Ⅰ的第一主成分代表寄生性昆虫物种数、寄生性昆虫个体数和中性昆虫物种数的综合因子, 累计贡献率为61.44%, 第二主成分代表捕食性昆虫个体数和植食性昆虫个体数的综合因子, 且第二主成分的贡献率为38.56%, 表明捕食性昆虫个体数和植食性昆虫个体数及寄生性昆虫物种和个体数对样地Ⅰ的昆虫群落组成有重要的影响作用。
样地Ⅱ的第一主成分代表捕食性昆虫物种数、寄生性昆虫物种数和寄生性昆虫个体数的综合因子, 累计贡献率为62.38%, 第二主成分代表捕食性昆虫个体数和中性昆虫物种数的综合因子, 贡献率为37.62%, 表明捕食性昆虫和寄生性昆虫是样地Ⅱ的主导因子。
样地Ⅲ的第一主成分代表植食性昆虫物种数、植食性昆虫个体数和中性昆虫个体数的综合因子, 贡献率为61.81%, 第二主成分代表捕食性昆虫个体数和寄生性昆虫物种数的综合因子, 第二主成分的贡献率为38.19%, 昆虫群落的不同类群均对样地Ⅲ有大小不同的影响, 表明样地Ⅲ的昆虫物种分布比较均衡, 也解释了样地Ⅲ均匀度最高的原因。
样地Ⅳ的第一主成分代表植食性昆虫物种数、寄生性昆虫个体数和中性昆虫个体数的综合因子, 贡献率为59.71%, 第二主成分代表捕食性昆虫个体数和寄生性昆虫物种数的综合因子, 贡献率为40.29%, 略低于第一主成分, 表明第一、二主成分的综合因子均对样地Ⅳ有主导作用。
样地Ⅴ的第一主成分代表植食性昆虫物种数、寄生性昆虫物种数和寄生性昆虫个体数的综合因子, 贡献率为78.96%, 第二主成分代表捕食性昆虫个体数, 贡献率为21.04%, 明显低于第一主成分, 表明第一主成分的综合因子是影响样地Ⅴ昆虫群落结构的重要因素。
样地Ⅵ的第一主成分代表植食性昆虫物种数、寄生性昆虫个体数和中性昆虫物种数的综合因子, 贡献率为70.90%, 第二主成分代表捕食性昆虫物种数和捕食性昆虫个体数的综合因子, 贡献率为29.10%。
综合以上分析结果可得出, 捕食性昆虫和寄生性昆虫是影响不同林分类型内昆虫群落结构的主要因素, 各林分类型中捕食性昆虫和寄生性昆虫对植食性昆虫的控制作用比较强, 昆虫群落结构比较稳定。
2.5 群落的稳定性分析由表 6可以看出, 不同林分类型St/Si的比值排序为样地Ⅴ>样地Ⅲ>样地Ⅳ>样地Ⅵ>样地Ⅰ>样地Ⅱ, 样地Ⅴ和样地Ⅲ的比值明显高于其它样地, 表明落叶松-白桦混交林和落叶松-樟子松-山刺玫混交林内的昆虫物种数相对于个体数而言较多, 昆虫种类分布相对均衡, 种间制约作用较强;Sn/Sp的比值排序为样地Ⅲ>样地Ⅵ>样地Ⅱ>样地Ⅰ>样地Ⅳ>样地Ⅴ, 样地Ⅵ和样地Ⅲ的比值明显高于其它, 表明落叶松-白桦混交林和落叶松-樟子松-白桦-山荆子-山刺玫混交林内的天敌类群相对于其本身的植食类群来说较多, 昆虫群落内部的相互制约关系比较复杂, 所以昆虫群落的稳定性和对外界的抗干扰能力比较高。
| 比值 Ratio |
样地Plot | |||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | |
| St/Si | 0.0426 | 0.0280 | 0.0862 | 0.0641 | 0.0932 | 0.0444 |
| Sn/Sp | 0.7234 | 0.8837 | 1.0500 | 0.6970 | 0.3500 | 0.9737 |
昆虫群落特征指数是反应群落结构水平的重要依据, 不仅代表了群落的多样性、丰富度、均匀度及变化规律等, 也在一定程度上体现了其所处地理环境、生存环境及林分状况等条件[16], 甚至一些特殊昆虫可以作为评价某一生态系统或环境的指示物种[17]。因此从昆虫群落结构的角度分析不同林分类型的多样性和稳定性, 对于寻求科学的利于林木生长发育的条件及科学的经营管理措施具有很大意义[18], 为计划近自然森林经营活动提供参考依据。本研究结果显示, 不同林分类型内昆虫种类丰富, 昆虫群落多样性指数均高, 不同林分类型间无明显差异, 个别林分在丰富度、均匀度和优势度指数上存在显著差异。在不同林分类型中捕食性和寄生性天敌是影响昆虫群落结构的主要因子, 对植食性昆虫的控制作用较强, 群落相似性分析显示不同林分类型内昆虫群落结构不相似, 表明不同林分类型内的树种匹配和混交类型对昆虫物种变化有明显影响。
由于在近自然森林经营模式下的塞罕坝自然保护区内植物资源丰富, 混交类型复杂、多变, 且林下植被丰富、盖度较大, 为昆虫群落的生存提供了适合的生活环境, 提高了昆虫群落的丰富度和多样性, 不仅植食性昆虫种类丰富, 同时植食性昆虫又为天敌昆虫的生长提供了营养供给, 使捕食性和寄生性昆虫对植食性昆虫具有一定的控制作用, 有效地避免了森林害虫的大发生, 同时也提高了自身的稳定性和抵御外界干扰的能力。
大量的研究结果[2, 19-21]表明, 林分类型越复杂, 昆虫群落多样性越高, 森林抵抗外界干扰的能力越强。本研究发现保护区内华北落叶松纯林和林分结构简单的落叶松-樟子松混交林亦表现为较高的昆虫群落多样性和稳定性, 主要是由于对保护区采用自然更新、封山育林等近自然森林经营措施, 使保护区很少受到外界因素的干扰, 自身形成了封闭且复杂的生态系统, 昆虫群落内部的控制作用较强, 森林对害虫具有一定的自然调控能力, 很少有害虫发生, 表现较为稳定。加上落叶松林下植被物种丰富度和盖度较高, 为昆虫提供了充足的取食和生存条件, 昆虫群落多样性相对较高, 群落结构复杂, 控害能力较强。不同林分类型昆虫群落结构不相似, 表明不同林分类型虽然具有丰富的昆虫物种, 但群落间共有种较少, 不同林分群落结构对昆虫有明显影响。昆虫群落在一定程度上受植物群落的影响而变化[22-23], 林下植被多样性高的林分内, 昆虫群落的丰富度、多样性高, 因此考虑林分类型对昆虫群落多样性的影响时, 不仅要考虑树种, 而且更不能忽略林下植被对昆虫群落的作用。
本文进一步说明在近自然森林经营下的人工林表现了较高的生物多样性, 具有良好的生态效益, 能够较好地发挥森林生态系统自身的稳定性, 并有效抵御外界干扰。因此, 我国当前应转变传统的森林经营管理模式, 充分利用森林生态系统生物间关系, 科学营林, 合理配置林木的空间结构, 选择适当的造林树种和密度, 建立近自然森林经营的管理理念[24], 对人工林进行科学的经营管理, 才能更好地发挥森林生态系统本身的稳定性和调控作用, 实现森林资源的生态效益和经济效益, 达到可持续利用和健康发展的状态。
| [1] | 刘兴平, 刘向辉, 王国红, 韩瑞东, 戈峰. 多样化松林中昆虫群落多样性特征. 生态学报, 2005, 25(11): 2976–2982. DOI:10.3321/j.issn:1000-0933.2005.11.026 |
| [2] | 马国强, 刘贤谦. 山西同朔地区不同混交林中昆虫群落结构初步研究. 山西农业大学学报:自然科学版, 2007, 27(4): 360–362. |
| [3] | 李菁, 骆有庆, 石娟. 基于生物多样性保护的兴安落叶松与白桦最佳混交比例——以阿尔山林区为例. 生态学报, 2012, 32(16): 4943–4949. |
| [4] | 周丕宁, 陈尚文, 李殷奎. 广西大明山保护区天然林昆虫多样性研究. 广西科学, 2009, 16(1): 92–95. |
| [5] | 曹金亮. 福建紫薇天然林昆虫群落结构特征及多样性研究. 安徽农业科学, 2014, 42(24): 8183–8184. DOI:10.3969/j.issn.0517-6611.2014.24.038 |
| [6] | 吴迪. 红松林的近自然经营技术研究[D]. 长沙: 中南林业科技大学, 2014. |
| [7] | 王慧, 郭晋平. 中国人工林现状及其近自然经营. 现代农业科学, 2008, 15(10): 124–125. |
| [8] | 张云玲. 塞罕坝自然保护区森林生态系统服务功能价值研究[D]. 石家庄: 河北师范大学, 2012. |
| [9] | 王建军. 塞罕坝自然保护区植物多样性及3S辅助分析技术研究[D]. 北京: 北京林业大学, 2004. |
| [10] | 韩争伟, 马玲, 曹传旺, 张静, 王步勇. 太湖湿地昆虫群落结构及多样性. 生态学报, 2013, 33(14): 4387–4397. |
| [11] | 赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990: 134–160. |
| [12] | 丁岩钦. 昆虫数学生态学. 北京: 科学出版社, 1994. |
| [13] | 刘颖华. 森林草原交错带节肢动物群落组成与多样性的研究. 保定: 河北农业大学, 2006. |
| [14] | 高宝嘉, 张执中, 李镇宇. 封山育林对昆虫群落结构及多样性稳定性影响的研究. 生态学报, 1992, 12(1): 1–7. |
| [15] | 卢纹岱. SPSS for Windows统计分析. 北京: 电子工业出版社, 2000: 484–492. |
| [16] | 孙儒泳. 动物生态学原理 (第三版). 北京: 北京师范大学出版社, 2001: 394–403. |
| [17] | Choi S W. Diversity and composition of larger moths in three different forest types of Southern Korea. Ecological Research, 2008, 23(3): 503–509. DOI:10.1007/s11284-007-0406-8 |
| [18] | Alalouni U, Brandl R, Auge H, Schädler M. Does insect herbivory on oak depend on the diversity of tree stands?. Basic and Applied Ecology, 2014, 15(8): 685–692. DOI:10.1016/j.baae.2014.08.013 |
| [19] | 张飞萍, 尤民生. 不同林分类型毛竹林节肢动物群落的多样性与稳定性. 昆虫学报, 2007, 50(1): 31–37. |
| [20] | 申曙光, 高宝嘉, 张玉会, 王正文, 王洪霞, 王秀君. 不同植物类型昆虫群落结构的研究. 河北农业大学学报, 1997, 20(4): 61–65. |
| [21] | 徐郑周, 刘广营, 王广海, 赵国华, 马长明. 燕山山地华北落叶松人工林群落生物多样性及其生物量的研究. 林业资源管理, 2010(2): 43–49. |
| [22] | Jobidon R, Cyr G, Thiffault N. Plant species diversity and composition along an experimental gradient of northern hardwood abundance in Picea mariana plantations. Forest Ecology and Management, 2004, 198(1/3): 209–221. |
| [23] | Moreira X, Abdala-Roberts L, Rasmann S, Castagneyrol B, Mooney K A. Plant diversity effects on insect herbivores and their natural enemies:current thinking, recent findings, and future directions. Current Opinion in Insect Science, 2016, 14: 1–7. DOI:10.1016/j.cois.2015.10.003 |
| [24] | 彭舜磊, 王得祥, 赵辉, 杨涛. 我国人工林现状与近自然经营途径探讨. 西北林学院学报, 2008, 23(2): 184–188. |