 2017, Vol. 37
2017, Vol. 37文章信息
- 阿加尔·恰肯, 吾玛尔·阿布力孜, 排孜力耶·合力力.
- Ajar QAKEN, Omar ABLIZ, Fazliya HELIL.
- 新疆玛纳斯河流域土壤螨类群落多样性
- Community diversity of soil mites in the Manas River Basin, Xinjiang
- 生态学报. 2017, 37(24): 8385-8396
- Acta Ecologica Sinica. 2017, 37(24): 8385-8396
- http://dx.doi.org/10.5846/stxb201610112057
-
文章历史
- 收稿日期: 2016-10-11
- 网络出版日期: 2017-08-15

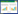




土壤螨类是陆地生态系统的重要功能组分, 它们在分解残体、改变土壤理化性质、土壤形成与发育、土壤物质迁移与能量转化等方面有着重要的作用[1-2]。由于螨类在各类土壤中普遍存在, 类群和数量丰富, 群落生物多样性高, 土壤螨类被看作是最重要的评价土壤质量变化的敏感指示生物之一[3-4]。目前大量的研究表明, 土壤螨类在植物根源的腐烂、陆地腐烂、稳定土壤和生物氮固定中起着很重要的作用[4-6]。土壤螨类对任何生物的干扰能做出迅速敏感的改变和响应, 能对环境的影响程度、干扰、栖息地的破坏、化学污染等给予早期的预警[3]。在国外螨类研究已有170多年的历史, 而我国从20世纪80年代开始土壤螨类生态学方面的研究, 虽然经过30多年的发展, 在土壤螨类研究方面地区发展仍很不均衡, 沿海地区研究较早并广泛, 而西北干旱区, 特别是新疆天山南北广大区域土壤螨类区系分类及其多样性的研究仍仅有零星报道[6-11]。
玛纳斯河流域地处内陆干旱区典型的山地-绿洲-荒漠生态系统Mountain-Oasis-Desert System (MODS), 随海拔不同, 自然景观分异明显[12]。该区域以其深居内陆的地理区位、干旱的大陆性气候、山盆相间的地貌格局、广泛发育的内陆流域、荒漠植被的特点与我国东部季风区和青藏高原区形成鲜明的对比, 垂直分异特征明显, 形成世界上特有的山地-绿洲-荒漠生态系统, 在全球干旱区类型中独具特色。随着全球气候变化, 植被、人类活动、土地利用等的影响, 呈明显的垂直地带分布, 生物群落均存在明显的区域性和差异性[12]。目前, 对该区域的研究主要涉及气候、水文、土壤、植物群落及其多样性等, 而对土壤螨类群落多样性方面的研究极少。本研究旨在查明该区域不同生境土壤螨类群落结构及其季节动态及其与环境的关系, 为干旱区土壤动物的系统研究以及土壤环境的生物学评价提供科学资料。
1 研究方法 1.1 研究区自然概况玛纳斯河流域位于天山北麓中段, 准噶尔盆地西南部, 地理位置为43°05′—46°04′N, 84°56′—86°42′E, 总面积3.09万km2。随海拔高度变化呈现高山冰雪带、高山及亚高山草甸带、中山森林带、低山草原带、荒漠带以及沙漠等自然景观带。新中国成立以后, 玛纳斯河流域成为新疆开垦面积最大的人工绿洲, 同时也是我国第四大灌溉农业区, 是自治区最重要的粮、棉、糖生产基地之一[12]。由于人口增加和大规模经济开发活动, 导致该流域的生态环境发生了巨大变化, 土壤严重的盐渍化和不同程度的污染现象, 土壤退化严重[12]。由于地貌、气候、水文条件、成土母质、植被具有明显的垂直地带分布, 土地利用类型复杂多样, 生物群落均存在明显的区域性和差异性, 使得本研究具有典型性。
1.2 样地设置结合研究区域景观、地形条件、土壤类型和土地利用状况等因素, 在研究区域设长达150 km的两条样带, 并在各条样带上选择了针叶林(Ⅰ)、山地草原(Ⅱ)、山地阔叶林(Ⅲ)、山地灌木林(Ⅳ)、山地耕地(Ⅴ)、防护林(Ⅵ)、平原耕地(Ⅶ)、草地(Ⅷ)、葡萄园(Ⅸ)、菜地(Ⅹ)、灌木林(Ⅺ)和荒漠(Ⅻ)等十二种不同生境进行采样, 探讨天山北麓典型山地、绿洲、荒漠生态系统不同垂直带生境土壤螨类群落多样性及其时空动态变化。
| 区域 Area |
海拔 Elevation/m |
地理位置 Geographic location |
年均气温 Annual temperature/ ℃ |
年均降水量 Annual precipitation/ mm |
年均蒸发量 Annual evaporation/ mm |
土壤类型 Soil types |
土地类型 Land type |
主要植物种类主要植物种类 The main plant species |
| 山地 Mountains |
700—3600 | 43°45′02″—43°59′08″N 85°46′24″—86°15′12″E |
2—5 | 500—700 | 850—1000 | 棕钙土 | Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ | 天山云杉(Picea schrenkiana)、疆方枝柏(Sabina pseudosabina(Fisch))、早熟禾属(Poa annua L) |
| 绿洲 Oasis |
400—600 | 44°21′47″—44°26′02″N 86°16′23″—86°20′20″E |
5—7 | 120—180 | 1500—1800 | 灰漠土 | Ⅵ、Ⅶ、Ⅷ、Ⅸ、Ⅹ | 沙枣(Elaeagnus angustifolia L)、棉花属(Gossypium spp)、玉米(Zea mays)、芦苇(Phragmites Adans)、黑葡萄(Vitis vinifera) |
| 荒漠 Desert |
250—400 | 44°27′06″—44°30′02″N 44°27′06″— 44°30′02″N |
6—8 | 100 —120 | 2000—2200 | 风沙土 | Ⅺ、Ⅻ | 柽柳(Tamarix ramosissima)、白刺(Nitraria sibirica)、骆驼刺(Alhagi sparsifolia) |
| 针叶林, coniferous forest(Ⅰ);山地草原, mountain steppe(Ⅱ);阔叶林, deciduous forest(Ⅲ);山地灌木林, mountain shrubbery(Ⅳ);山地耕地, mountain farmland(Ⅴ);防护林, shelter forest(Ⅵ);平原耕地, plain farmland(Ⅶ);草地, grassland(Ⅷ);葡萄园, vineyard(Ⅸ);菜地, vegetable field(Ⅹ);灌木林, shrubbery(Ⅺ)和荒漠, desert(Ⅻ) | ||||||||
2014年4、6、9和11月份中旬对上述的12种不同生境定点采样, 每个生境分别选取3个样点, 并采用容积为100 cm3土壤环刀, 按土壤深度0—5、5—10、10—15、15—20 cm的4层取样, 共取576份土样, 装入纸袋带回实验室, 采用改进的Tullgren法连续光照48 h分离土壤螨类, 在Leica体视显微镜下观察及制片。完成制片后, 参照尹文英的《中国土壤动物检索图鉴》、江原昭三《日本蜱螨类检索图鉴》、青木淳一《日本土壤动物检索图鉴》及J.Balogh和P.Balogh的《The Oribatid Mites Genera of the World》和Krantz(2009)《蜱螨学手册》(第三册)等进行分类鉴定, 一般鉴定到属的水平[13-17]。
1.4 主要环境因子的测定在调查土壤螨类的同时, 测定各采样点的主要环境因子(土壤容重、温度、湿度、pH、有机质、全氮、全磷、全钾和总盐含量)。土壤容重采用土壤环刀法;土壤温度和土壤酸碱分别使用地温计和土壤酸度计测定;土壤水分含量采用烘干法测定(GB 7172—1987), 按照以下公式计算含水率:含水率(%)=(G0-G2)(湿土重-干土重)/G2×100%。土壤有机质含量采用重铬酸钾一硫酸溶液氧化法(GB 9834—1988)测定。土壤全钾含量参照GB 9836—1988法测定;土壤全磷含量参照GB 9837—1988法测定;土壤全氮含量参照GB 7173—1987法测定;总盐量采用水溶性盐总量(质量法)测定(NY/T 1121.16—2006)[18]。
1.5 数据处理在不同生境土壤螨类群落多样性分析中, 进行了以下生态指标的比较分析[8]。
(1) Shannon-Wiener多样性指数(H):

|
(2) Margalef丰富度指数(M):

|
(3) Pielou均匀度指数(E):

|
(4) Simpson优势度指数(C):

|
(5) Jacccard群落相似性系数(q):

|
(6) S∅rensen相似性指数:

|
式中, N表示群落中所有类群的个体总数, S表示类群数, Pi等于ni/N, ni表示第i类群的个体数, a为A群落类群数, b为B群落类群数, c为两群落共有类群数。所有的数据处理运用SPSS 19.0进行处理, 并在Excel中作图, 用单因素方查分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
2 结果与分析 2.1 不同生境土壤螨类群落组成与数量分布调查期间共获螨类标本32308头, 分隶于86科4亚目140属, 其中甲螨亚目为20877只(64.62%), 分隶于80属51科, 中气门亚目为10167只(31.47%), 34属20科, 前气门亚目为1031只(3.19%), 21属12科, 无气门亚目为233只(0.72%), 5属3科。对整个研究区域而言, Ceratozetes和Oribatula为优势类群, 占总螨类捕获量的21.32%;Epilohmannia、Trhypochthonius、Camisia、Oppiella、Tectocepheus等18类为常见类群, 占螨类总捕获量的55.89%, 优势类群和常见类群占总捕获量的77.21%。Palaeacarus、Belba、Epidamaeus等120类均为稀有类群(表 2)。
| 螨类Mites taxa | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | Ⅺ | Ⅻ | 总数 Total |
密度 Density |
百分比 Percentage/ % |
优势度 Abundace degree |
| 古甲螨属Palaeacarus | 61 | 12 | 1 | 31 | 16 | 15 | 23 | 15 | 39 | 39 | 15 | 3 | 270 | 1220.8 | 0.84 | + |
| 拟古甲螨属Palaeacaroides | 26 | 0 | 1 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 38 | 171.8 | 0.12 | + |
| 广缝甲螨属Cosmochthonius | 1 | 0 | 0 | 0 | 0 | 47 | 3 | 4 | 26 | 22 | 14 | 19 | 136 | 614.9 | 0.42 | + |
| 小缝甲螨属Hypochthoniella | 77 | 6 | 31 | 33 | 63 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 210 | 949.5 | 0.65 | + |
| 滑缝甲螨属Liochthonius | 122 | 53 | 32 | 42 | 39 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 288 | 1302.2 | 0.89 | + |
| 短甲螨属Brachychthonius | 14 | 0 | 14 | 2 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 47 | 212.5 | 0.15 | + |
| 塞缝甲螨Sellnickochthonius | 0 | 12 | 45 | 4 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 78 | 352.7 | 0.24 | + |
| 缝甲螨属Hypochthonius | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 40.7 | 0.03 | + |
| 派缝甲螨属Parhypochthonius | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 盲甲螨属Malaconothrus | 0 | 0 | 0 | 0 | 0 | 11 | 7 | 0 | 0 | 0 | 0 | 0 | 18 | 81.4 | 0.06 | + |
| 罗甲螨属Lohmannia | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 7 | 2 | 0 | 14 | 63.3 | 0.04 | + |
| 混居甲螨属Mixacarus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 | 11 | 0 | 20 | 90.4 | 0.06 | + |
| 上罗甲螨属Epilohmannia | 64 | 31 | 35 | 86 | 89 | 252 | 53 | 111 | 89 | 18 | 89 | 43 | 960 | 4340.7 | 2.97 | ++ |
| 拟上罗甲螨Epilohmannoides | 4 | 21 | 16 | 0 | 9 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 50 | 226.1 | 0.15 | + |
| 全罗甲螨属Perlohmannia | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 | 85.9 | 0.06 | + |
| 矮汉甲螨属Nanhermannia | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 31.7 | 0.02 | + |
| 懒甲螨属Nothrus | 69 | 26 | 4 | 26 | 21 | 88 | 5 | 21 | 31 | 3 | 23 | 0 | 317 | 1433.3 | 0.98 | + |
| 礼服甲螨属Trhypochthonius | 5 | 209 | 30 | 112 | 138 | 154 | 70 | 74 | 475 | 13 | 70 | 69 | 1419 | 6416.2 | 4.39 | ++ |
| 异懒甲螨属Allonothrus | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 原甲螨属Archegozetes | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 洼甲螨属Camisia | 15 | 8 | 0 | 24 | 4 | 346 | 8 | 50 | 65 | 29 | 50 | 0 | 599 | 2708.4 | 1.85 | ++ |
| 半懒甲螨属Heminothrus | 10 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 90.4 | 0.06 | + |
| 平懒甲螨属Platynothrus | 4 | 10 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 90.4 | 0.06 | + |
| 叶赫甲螨属Phyllhermannia | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 直卷甲螨属Archoplophora | 0 | 0 | 0 | 0 | 0 | 32 | 0 | 16 | 8 | 4 | 16 | 0 | 76 | 343.6 | 0.24 | + |
| 卷甲螨属Phthiracarus | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 三皱甲螨属Rhysotritia | 43 | 1 | 26 | 11 | 15 | 261 | 3 | 37 | 54 | 38 | 70 | 0 | 559 | 2527.6 | 1.73 | ++ |
| 龙足甲螨属Eremaeus | 2 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 22.6 | 0.02 | + |
| 沙甲螨属Eremulus | 21 | 0 | 0 | 0 | 0 | 4 | 11 | 0 | 0 | 0 | 8 | 0 | 44 | 199.0 | 0.14 | + |
| 沙足甲螨属Eremobellba | 4 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 3 | 0 | 11 | 49.7 | 0.03 | + |
| 珠足甲螨属Belba | 135 | 14 | 0 | 42 | 18 | 10 | 1 | 3 | 2 | 1 | 1 | 0 | 227 | 1026.4 | 0.70 | + |
| 表珠甲螨属Epidamaeus | 120 | 6 | 0 | 5 | 29 | 9 | 0 | 2 | 0 | 1 | 1 | 3 | 176 | 795.8 | 0.54 | + |
| 扇珠足甲螨属Licnodamaeus | 0 | 0 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 1 | 1 | 3 | 14 | 63.3 | 0.04 | + |
| 小奥甲螨属Oppiella | 343 | 59 | 50 | 142 | 154 | 799 | 8 | 29 | 70 | 226 | 23 | 0 | 1903 | 8604.6 | 5.89 | ++ |
| 多奥甲螨属Multioppia | 76 | 0 | 4 | 15 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 95 | 429.6 | 0.29 | + |
| 纹奥甲螨属Striatoppia | 2 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 奥甲螨Oppia | 68 | 7 | 0 | 4 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 98 | 443.1 | 0.30 | + |
| 四奥甲螨属Quadroppia | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 大奥甲螨属Lasiobelba | 18 | 21 | 2 | 4 | 12 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 57 | 257.7 | 0.18 | + |
| 丽甲螨属Liacarus | 60 | 30 | 13 | 11 | 0 | 326 | 124 | 183 | 157 | 127 | 203 | 0 | 1234 | 5579.7 | 3.82 | ++ |
| 弯步甲螨属Gibbicepheus | 0 | 0 | 0 | 0 | 0 | 12 | 1 | 1 | 0 | 1 | 1 | 0 | 16 | 72.3 | 0.05 | + |
| 步甲螨属Carabides | 0 | 0 | 0 | 0 | 0 | 13 | 1 | 2 | 0 | 1 | 1 | 0 | 18 | 81.4 | 0.06 | + |
| 原步甲螨属Archegocepheus | 0 | 0 | 0 | 0 | 0 | 12 | 1 | 1 | 0 | 1 | 0 | 0 | 15 | 67.8 | 0.05 | + |
| 龙骨足甲螨属Eremaeus | 2 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 22.6 | 0.02 | + |
| 隐助甲螨属Costerems | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 大头甲螨属Megalotocepheus | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 裂头甲螨属Fissicepheus | 71 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 82 | 370.8 | 0.25 | + |
| 阿斯甲螨属Astegistes | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 2 | 5 | 0 | 0 | 10 | 45.2 | 0.03 | + |
| 叉肋甲螨属Furcoribula | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 9.0 | 0.01 | + |
| 刀肋甲螨属Cultroribula | 197 | 5 | 21 | 27 | 7 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 259 | 1171.1 | 0.80 | + |
| 小梳甲螨属Xenillus | 42 | 0 | 12 | 0 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 73 | 330.1 | 0.23 | + |
| 垂盾甲螨属Scutovertex | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 39 | 53 | 3 | 27 | 0 | 128 | 578.8 | 0.40 | + |
| 下盾甲螨属Hypovertex | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 跳甲螨属Zetorchestes | 71 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 73 | 330.1 | 0.23 | + |
| 盖头甲螨属Tectocepheus | 150 | 9 | 128 | 100 | 147 | 115 | 16 | 14 | 19 | 40 | 74 | 0 | 812 | 3671.5 | 2.51 | ++ |
| 大翼甲螨属Galumna | 15 | 0 | 0 | 0 | 0 | 80 | 2 | 15 | 13 | 4 | 25 | 13 | 167 | 755.1 | 0.52 | + |
| 全大翼甲螨属Pergalumna | 90 | 7 | 3 | 34 | 12 | 21 | 50 | 0 | 0 | 0 | 27 | 0 | 244 | 1103.3 | 0.76 | + |
| 尖棱甲螨属Ceratozetes | 107 | 20 | 91 | 128 | 81 | 843 | 138 | 821 | 333 | 157 | 558 | 0 | 3277 | 14817.3 | 10.14 | +++ |
| 单奥甲螨属Phauloppia | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 若甲螨属Oribatula | 212 | 126 | 91 | 129 | 8 | 1209 | 241 | 88 | 544 | 346 | 617 | 0 | 3611 | 16327.5 | 11.18 | +++ |
| 合若甲螨属Zygoribatula | 10 | 0 | 0 | 10 | 8 | 73 | 211 | 93 | 60 | 43 | 117 | 0 | 625 | 2826.0 | 1.93 | ++ |
| 缰板鳃甲螨属Chamobates | 1 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 菌甲螨属Scheloribates | 60 | 30 | 13 | 11 | 36 | 326 | 124 | 183 | 157 | 127 | 203 | 18 | 1288 | 5823.8 | 3.99 | ++ |
| 足肋甲螨属Podoribates | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 角翼甲螨属Achipteria | 22 | 134 | 2 | 30 | 48 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 236 | 1067.1 | 0.73 | + |
| 蛇轮甲螨属Ophidiotrichus | 0 | 0 | 0 | 0 | 19 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 | 85.9 | 0.06 | + |
| 小甲螨属Oribatella | 3 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 8 | 36.2 | 0.02 | + |
| 新助甲螨属Neoribates | 20 | 0 | 2 | 16 | 18 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 56 | 253.2 | 0.17 | + |
| 副大翼甲螨属Parakalumna | 0 | 3 | 0 | 9 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 15 | 67.8 | 0.05 | + |
| 原大翼甲螨属Protokalumna | 7 | 9 | 0 | 65 | 17 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 100 | 452.2 | 0.31 | + |
| 顶鳞甲螨属Lepidozetes | 2 | 11 | 2 | 48 | 11 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 74 | 334.6 | 0.23 | + |
| 长单翼甲螨属Protoribates | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 16 | 72.3 | 0.05 | + |
| 细若甲螨属Incabates | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 圆单翼甲螨属Peloribates | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 27.1 | 0.02 | + |
| 角单翼甲螨属Rostrozetes | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 木单翼甲螨属Xylobates | 30 | 0 | 0 | 0 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 40 | 180.9 | 0.12 | + |
| 梁甲螨属Lamellobates | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 南角翼甲螨Austrachipteria | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 钉棱甲螨属Passalozetes | 0 | 0 | 0 | 0 | 0 | 23 | 0 | 0 | 11 | 4 | 0 | 103 | 141 | 637.5 | 0.44 | + |
| 真前翼甲螨属Eupelops | 32 | 168 | 21 | 85 | 76 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 382 | 1727.3 | 1.18 | ++ |
| 足助甲螨属Podoribates | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 派盾螨属Parholaspis | 12 | 9 | 0 | 5 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 27 | 122.1 | 0.08 | + |
| 革板螨属Gamasholaspis | 5 | 0 | 0 | 3 | 17 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 113.0 | 0.08 | + |
| 新派盾螨属Neparholaspis | 19 | 11 | 0 | 4 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 39 | 176.3 | 0.12 | + |
| 前小派伦螨Proparholaspulus | 3 | 1 | 0 | 21 | 20 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 45 | 203.5 | 0.14 | + |
| 派伦螨属Parholaspulus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 浩伦螨属Holaspulus | 0 | 0 | 0 | 0 | 0 | 291 | 221 | 301 | 70 | 147 | 101 | 23 | 1154 | 5217.9 | 3.57 | ++ |
| 厉螨属Laelaps | 73 | 39 | 29 | 52 | 57 | 37 | 4 | 9 | 0 | 0 | 13 | 25 | 338 | 1528.3 | 1.05 | ++ |
| 上厉螨属Ololaelaps | 5 | 33 | 13 | 7 | 3 | 216 | 64 | 82 | 61 | 40 | 90 | 27 | 641 | 2898.3 | 1.98 | ++ |
| 维螨属Veigaia | 1 | 0 | 4 | 0 | 6 | 16 | 2 | 82 | 0 | 9 | 8 | 0 | 128 | 578.8 | 0.40 | + |
| 革厉螨属Gamasolaelaps | 0 | 0 | 0 | 0 | 0 | 4 | 2 | 3 | 0 | 1 | 4 | 0 | 14 | 63.3 | 0.04 | + |
| 巨螯螨属Macrocheles | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 13 | 0 | 17 | 76.9 | 0.05 | + |
| 全盾螨属Holostaspella | 0 | 0 | 0 | 0 | 0 | 226 | 99 | 39 | 83 | 40 | 99 | 21 | 607 | 2744.6 | 1.88 | ++ |
| 犹伊螨属Eviphis | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | 1 | 2 | 0 | 0 | 7 | 31.7 | 0.02 | + |
| 厚厉螨属Pachylaelaps | 0 | 1 | 0 | 0 | 0 | 486 | 42 | 246 | 129 | 72 | 333 | 0 | 1309 | 5918.8 | 4.05 | ++ |
| 虫穴螨属Zercon | 183 | 60 | 17 | 77 | 58 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 395 | 1786.0 | 1.22 | ++ |
| 尾足螨属Uroplitella | 1 | 97 | 5 | 7 | 22 | 345 | 10 | 46 | 11 | 62 | 32 | 4 | 642 | 2902.9 | 1.99 | ++ |
| 巨刺螨属Macronyssus | 0 | 0 | 0 | 0 | 0 | 5 | 0 | 2 | 0 | 1 | 2 | 0 | 10 | 45.2 | 0.03 | + |
| 尾卵螨属Uroobovella | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 海厉螨属Halolaelaps | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 线叶爪螨属Linopenthaleus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| Chaussieria Oudemans, 1902 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 长须螨属Stigmaeus | 0 | 0 | 0 | 0 | 0 | 24 | 7 | 0 | 6 | 26 | 2 | 7 | 72 | 325.6 | 0.22 | + |
| 小革螨属Gamasellus | 1 | 0 | 7 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 13 | 58.8 | 0.04 | + |
| 巨须螨属Cunaxa | 0 | 0 | 0 | 0 | 0 | 108 | 2 | 0 | 4 | 14 | 7 | 0 | 135 | 610.4 | 0.42 | + |
| 真长须螨属Eustigmaeus | 0 | 0 | 0 | 38 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 38 | 171.8 | 0.12 | + |
| 毛绥螨属Lasioseius | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 盲蛛螨属Caeculus | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 9.0 | 0.01 | + |
| 土皮须螨属Ledermuelleria | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 手绥螨属Cheiroseius | 0 | 0 | 0 | 0 | 0 | 37 | 0 | 0 | 10 | 5 | 0 | 0 | 52 | 235.1 | 0.16 | + |
| 表刻螨属Epicrius | 4 | 9 | 3 | 40 | 17 | 18 | 0 | 3 | 2 | 5 | 12 | 0 | 113 | 510.9 | 0.35 | + |
| 美绥螨属Ameroseius | 0 | 0 | 0 | 0 | 0 | 257 | 47 | 110 | 171 | 121 | 114 | 21 | 841 | 3802.7 | 2.60 | ++ |
| 足角螨属Podocinum | 0 | 0 | 0 | 0 | 0 | 14 | 12 | 17 | 13 | 1 | 36 | 0 | 93 | 420.5 | 0.29 | + |
| 麦矮蒲螨属Mahunkania | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 0 | 4 | 18.1 | 0.01 | + |
| 赤螨属Erythraeus | 0 | 0 | 1 | 0 | 0 | 70 | 0 | 1 | 0 | 7 | 0 | 0 | 79 | 357.2 | 0.24 | + |
| 开依丽螨属Caeculisoma | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 纤赤螨属Leptus | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 2 | 0 | 5 | 22.6 | 0.02 | + |
| 吸螨属Bdella | 0 | 0 | 0 | 0 | 0 | 11 | 4 | 13 | 16 | 8 | 9 | 9 | 70 | 316.5 | 0.22 | + |
| 盾螨属Scutaracus | 0 | 0 | 0 | 0 | 0 | 88 | 2 | 6 | 13 | 5 | 106 | 0 | 220 | 994.8 | 0.68 | + |
| 莓螨属Rhagidius | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 异绒螨属Allothrombium | 1 | 4 | 0 | 3 | 0 | 50 | 1 | 5 | 1 | 3 | 3 | 0 | 71 | 321.0 | 0.22 | + |
| 小爪螨属Oligonychus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 岩螨属Petrobia | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 4 | 0 | 0 | 7 | 31.7 | 0.02 | + |
| 裂爪螨属Schizotetranychus | 0 | 0 | 17 | 121 | 86 | 10 | 0 | 45 | 0 | 2 | 4 | 0 | 285 | 1288.7 | 0.88 | + |
| Yezonychus Ehara | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 单头螨属Aplonibia | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 4 | 5 | 22.6 | 0.02 | + |
| 肛厉螨属Proctolaelaps | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 植绥螨属Phytoseius | 0 | 0 | 0 | 0 | 0 | 156 | 27 | 121 | 141 | 54 | 88 | 58 | 645 | 2916.4 | 2.00 | ++ |
| 尾绥螨属Uroseius | 0 | 0 | 0 | 0 | 0 | 51 | 0 | 32 | 0 | 0 | 36 | 0 | 119 | 538.1 | 0.37 | + |
| 皮刺螨属Dermanyssus | 1 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 | 22.6 | 0.02 | + |
| 裂胸螨属Aceosejus | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.5 | 0.00 | + |
| 皮膜螨属Laminosioptes | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 13.6 | 0.01 | + |
| 隐爪螨属Nanorchestes | 0 | 0 | 0 | 0 | 0 | 6 | 4 | 1 | 0 | 1 | 0 | 0 | 12 | 54.3 | 0.04 | + |
| 吻体螨属Smaridius | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 3 | 0 | 0 | 5 | 22.6 | 0.02 | + |
| 脂螨属Lardoglyphus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 226 | 0 | 0 | 1 | 0 | 227 | 1026.4 | 0.70 | + |
| 短足螨属Pygmyphorus | 0 | 0 | 0 | 0 | 0 | 6 | 1 | 0 | 0 | 0 | 1 | 7 | 15 | 67.8 | 0.05 | + |
| 嗜粉螨属Aleuroglyphus | 0 | 0 | 0 | 0 | 0 | 115 | 166 | 653 | 177 | 105 | 1644 | 0 | 2860 | 12931.8 | 8.85 | ++ |
| 食酪螨属Tyrophagus | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 1 | 0 | 3 | 13.6 | 0.01 | + |
| 粉螨属Acarus | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 4 | 18.1 | 0.01 | + |
| 根螨属Clapared | 2 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 6 | 27.1 | 0.02 | + |
| 个体数(N)Individuals | 2868 | 1327 | 814 | 1680 | 1488 | 7787 | 1823 | 3878 | 3130 | 2014 | 5019 | 480 | 32308 | 146083.9 | 100.00 | |
| 类群数(S)Group number | 72 | 41 | 49 | 44 | 45 | 67 | 44 | 54 | 45 | 55 | 57 | 20 | 1043.5±428 | |||
| +++优势类群, 个体数占总捕获量的10%以上Dominant group, individual number is more than 10% of total individuals;++常见类群, 个体数占总捕获量的1%—10% Frequent group, individual number is between 10% and 1%;+稀有类群, 个体数占总捕获量的1%以下Rare group, individual number is less than 1%;针叶林, coniferous forest(Ⅰ);山地草原, mountain steppe(Ⅱ);阔叶林, deciduous forest(Ⅲ);山地灌木林, mountain shrubbery(Ⅳ);山地耕地, mountain farmland(Ⅴ);防护林, shelter forest(Ⅵ);平原耕地, plain farmland(Ⅶ);草地, grassland(Ⅷ);葡萄园, vineyard(Ⅸ);菜地, vegetable field(Ⅹ);灌木林, shrubbery(Ⅺ)和荒漠, desert(Ⅻ) | ||||||||||||||||
相关分析表明, 对整个研究区域来讲, 在12种不同生境土壤螨类个体数量和类群数量之间差异显著(P < 0.05), 其中个体数依次为Ⅵ(7787只, 24.10%)>Ⅺ(5019只, 15.53%)>Ⅷ(3878只, 12.00%)>Ⅸ(3130只, 9.69%)>Ⅰ(2868只, 8.88%)>Ⅹ(2014只, 6.23%)>Ⅶ(1823只, 5.64%)>Ⅳ(1680只, 5.20%)>Ⅴ(1488只, 4.61%)>Ⅱ(1327只, 4.11%)>Ⅲ(814只, 2.52%)>Ⅻ(480只, 1.49%)。在生境Ⅵ、Ⅺ及Ⅷ内个体数较高与其植被分布多样有关, 随着海拔高度增加, 生境条件不同, 造成了螨类个体数在各生境分布差异。
类群数依次为Ⅰ(72属, 51.43%)>Ⅵ(67属, 47.86%)>Ⅺ(57属, 40.71%)>Ⅹ(55属, 39.29%)>Ⅷ(54属, 38.57%)>Ⅲ(49属, 35%)>Ⅴ(45属, 32.14%)>Ⅸ(45属, 32.14%)>Ⅺ(44属, 31.43%)>Ⅶ(44属, 31.43%)>Ⅱ(41属, 29.29%)>Ⅻ(20属, 14.29%)。由于微环境生态条件各异, 导致类群数分布不均匀, 见表 2。
2.2 不同生境土壤螨类群落的垂直分布从垂直分布来看, 土壤螨类主要集中分布于表层土壤, 并呈现出明显的表聚性, 即0—5 cm最多(19588只, 60.63%), 其次为5—10 cm(7290只, 22.56%), 在10—15 cm中较少(3724只, 11.53%), 而在15—20 cm中最少(1706只, 5.28%)。不同生境土壤螨类个体数之间单因素分析表明, 在各生境不同土层之间差异显著(P < 0.05), 这与土壤有机质、温湿度垂直分布差异有关(图 1)。

|
| 图 1 玛纳斯河流域不同生境土壤螨类群落的垂直分布 Fig. 1 The vertical distribution of soil mite communities at different habitats in Manas River Basin 针叶林, coniferous forest(Ⅰ);山地草原, mountain steppe(Ⅱ);阔叶林, deciduous forest(Ⅲ);山地灌木林, mountain shrubbery(Ⅳ);山地耕地, mountain farmland(Ⅴ);防护林, shelter forest(Ⅵ);平原耕地, plain farmland(Ⅶ);草地, grassland(Ⅷ);葡萄园, vineyard(Ⅸ);菜地, vegetable field(Ⅹ);灌木林, shrubbery(Ⅺ)和荒漠, desert(Ⅻ) |
从春夏秋冬4个季节调查结果来看, 不同季节各种生境土壤螨类的个体数和类群数量除了人为干扰程度较大的Ⅸ、Ⅹ和Ⅵ生境以外, 其他生境均有一定的变化规律, 即秋季最多、其次为夏季。类群数和个体数的季节动态变化依次为秋季(15146只, 46.88%)>春季(6682只, 20.68%)>冬季(6041只, 18.7%)>夏季(4439只, 13.74%), 这可能与秋冬季节枯枝落叶普遍增厚, 螨类的食物丰富有关, 详情见图 2。

|
| 图 2 玛纳斯河流域不同生境土壤螨类群落的季节动态 Fig. 2 Seasonal dynamics of soil mite communities at different habitats of Manas River Basin |
由表 3可知, Shannon-Wiener多样性指数(H)在Ⅰ生境最高、而在ⅩⅡ生境最低;Margalef丰富度指数(M)在Ⅰ生境最高、而在ⅩⅡ生境最低;Simpson优势度指数(C)在ⅩⅠ生境最高, 而在Ⅳ生境最低;Pielou均匀度指数(E)在Ⅳ生境最高, 在ⅩⅠ生境最低。在12种不同生境之间土壤螨类群落多样性均有显著差异(P < 0.05), 其中Shannon-Wiener多样性指数(H)为:Ⅰ>Ⅴ>Ⅳ>Ⅵ>Ⅲ>Ⅹ>Ⅱ>Ⅸ>Ⅷ>Ⅶ>Ⅺ>Ⅻ。Margalef(M)丰富度指数依次为Ⅰ>Ⅵ>Ⅲ>Ⅹ>Ⅺ>Ⅷ>Ⅴ>Ⅳ>Ⅶ>Ⅱ>Ⅸ>Ⅻ。Pielou均匀度指数(E):Ⅳ>Ⅴ>Ⅻ>Ⅱ>Ⅰ>Ⅲ>Ⅸ>Ⅹ>Ⅴ>Ⅵ>Ⅷ>Ⅺ。Simpson优势度指数(C)为:Ⅺ>Ⅻ>Ⅷ>Ⅸ>Ⅶ>Ⅱ>Ⅹ>Ⅲ>Ⅵ>Ⅴ>Ⅳ>Ⅰ。多样性指标在山地生境均高, 因为植被分布多样与人为干扰较小有关。
| 多样性指标 Diversity indices |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | Ⅺ | Ⅻ | 全区 Total |
| 个体数(N) Individuals |
2868 | 1327 | 814 | 1680 | 1488 | 7787 | 1823 | 3878 | 3130 | 2014 | 5019 | 480 | 32308 |
| 类群数(S) Groups | 72 | 41 | 49 | 44 | 45 | 67 | 44 | 54 | 45 | 55 | 57 | 20 | 140 |
| 多样性指数(H) Diversity index |
3.409 | 2.980 | 3.082 | 3.289 | 3.294 | 3.140 | 2.836 | 2.844 | 2.871 | 3.005 | 2.605 | 2.537 | 3.545 |
| 优势度指数(C) Dominance index |
0.047 | 0.076 | 0.069 | 0.048 | 0.052 | 0.066 | 0.078 | 0.097 | 0.084 | 0.074 | 0.147 | 0.106 | 0.048 |
| 丰富度指数(M) Abundance index |
8.918 | 5.563 | 7.162 | 5.790 | 6.023 | 7.366 | 5.727 | 6.414 | 5.467 | 7.098 | 6.572 | 3.078 | 7.609 |
| 均匀度指数(E) Evenness index |
0.797 | 0.802 | 0.792 | 0.869 | 0.865 | 0.747 | 0.749 | 0.713 | 0.754 | 0.750 | 0.644 | 0.847 | 0.809 |
不同生境土壤螨类群落间的Jaccard相似性指数和S∅rensen相似性指数分析结果如表 4和表 5。
| 生境 Habitats |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | Ⅺ | Ⅻ |
| Ⅰ | 0.000 | 0.242 | 0.239 | 0.244 | 0.256 | 0.199 | 0.131 | 0.165 | 0.149 | 0.158 | 0.178 | 0.089 |
| Ⅱ | 0.00 | 0.224 | 0.292 | 0.286 | 0.211 | 0.150 | 0.190 | 0.167 | 0.158 | 0.190 | 0.090 | |
| Ⅲ | 0.000 | 0.250 | 0.236 | 0.134 | 0.147 | 0.149 | 0.130 | 0.140 | 0.165 | 0.092 | ||
| Ⅳ | 0.000 | 0.294 | 0.165 | 0.162 | 0.169 | 0.168 | 0.189 | 0.179 | 0.099 | |||
| Ⅴ | 0.000 | 0.164 | 0.160 | 0.168 | 0.159 | 0.153 | 0.177 | 0.097 | ||||
| Ⅵ | 0.000 | 0.270 | 0.248 | 0.228 | 0.295 | 0.262 | 0.187 | |||||
| Ⅶ | 0.000 | 0.263 | 0.252 | 0.267 | 0.284 | 0.200 | ||||||
| Ⅷ | 0.000 | 0.051 | 0.288 | 0.284 | 0.169 | |||||||
| Ⅸ | 0.000 | 0.265 | 0.250 | 0.188 | ||||||||
| Ⅹ | 0.000 | 0.282 | 0.176 | |||||||||
| Ⅺ | 0.000 | 0.189 | ||||||||||
| Ⅻ | 0.000 | |||||||||||
| 极相似Much similar(0.75—1.00), 中等相似Moderately similar(0.50—0.75), 中等不相似Moderately dissimilar(0.25—0.50), 极不相似Much dissimilar(0.00—0.25) | ||||||||||||
| 生境 Habitats |
Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | Ⅺ | Ⅻ |
| Ⅰ | 0.000 | 0.637 | 0.628 | 0.672 | 0.684 | 0.417 | 0.362 | 0.333 | 0.376 | 0.346 | 0.434 | 0.196 |
| Ⅱ | 0.000 | 0.578 | 0.824 | 0.791 | 0.426 | 0.447 | 0.421 | 0.442 | 0.375 | 0.469 | 0.164 | |
| Ⅲ | 0.000 | 0.667 | 0.617 | 0.310 | 0.344 | 0.350 | 0.298 | 0.327 | 0.396 | 0.203 | ||
| Ⅳ | 0.000 | 0.831 | 0.396 | 0.432 | 0.408 | 0.404 | 0.364 | 0.436 | 0.219 | |||
| Ⅴ | 0.000 | 0.393 | 0.382 | 0.404 | 0.378 | 0.360 | 0.431 | 0.215 | ||||
| Ⅵ | 0.000 | 0.739 | 0.661 | 0.643 | 0.836 | 0.710 | 0.460 | |||||
| Ⅶ | 0.000 | 0.714 | 0.674 | 0.727 | 0.792 | 0.500 | ||||||
| Ⅷ | 0.000 | 0.586 | 0.807 | 0.793 | 0.405 | |||||||
| Ⅸ | 0.000 | 0.720 | 0.667 | 0.462 | ||||||||
| Ⅹ | 0.000 | 0.786 | 0.427 | |||||||||
| Ⅺ | 0.000 | 0.468 | ||||||||||
| Ⅻ | 0.000 |
由表 4可见, 根据Jaccard相似性系数分析, 螨类群落间的相似性系数具有较大的差异, 最高值发生在生境Ⅴ(0.294), 最低点发生在生境ⅩⅡ(0.089)。所有系数中有49个系数在0和0.25之间表现为极不相似, 而17个系数在0.25和0.5之间表现为中等不相似。
由表 5可见, S∅rensen相似性系数在不同生境各异, 最高值发生在生境Ⅰ和生境Ⅹ之间(0.836), 最低点发生在生境Ⅰ和生境ⅩⅡ之间(0.164)。36个系数在0.25和0.50之间表现为中等不相似, 25个系数在0.50和0.75之间群落之间中等相似, 5个系数在0和0.25之间表现为极不相似。说明海拔高度、植被组成及人为干扰对螨类群落结构差异的影响很大。
2.6 不同生境土壤螨类群落指标与环境因子的关系玛纳斯河流域不同生境土壤螨类群落指标与环境因子之间的相关分析结果如表 6。
| 多样性指标Diversity indices | 海拔/m Elevation |
土温/℃ Temperature |
含水量/% Water content |
容重/ (g/cm3) Bulk density |
pH | 有机质/ (g/kg) Organic mater |
全氮/ (g/kg) Total nitrogen |
全磷/ (g/kg) Total phosphor |
全钾/ (g/kg) Total potassium |
总盐/ (g/kg) Total salt |
| 个体数(N) Individual number | 0.738** | 0.162* | 0.215* | -0.214** | -0.255* | 0.818** | 0.382** | 0.525* | 2.303* | 0.214* |
| 类群数(S) Group number | 0.706** | 0.215* | 0.154* | -0.186** | -0.212 | 0.712** | 0.356* | 0.384 | 2.436 | -0.192 |
| 多样性指数(H) Diversity index | -0.618** | 0.232** | -0.205** | -0.512** | -0.285* | 0.648* | 0.352** | 0.438* | 3.215 | -0.215* |
| 优势度指数(C) Dominance index | 0.626* | -0.152 | 0.212* | 0.482* | 0.305* | 0.496* | 0.272* | -0.652 | 2.306* | 0.262* |
| 丰富度指数(M) Abundance index | 0.734** | 0.184** | -0.254** | 0.484* | -0.255 | 0.732** | 0.392** | -0.476* | 4.214* | 0.236* |
| 均匀度指数(E) Evenness index | 0.626 | -0.152* | 0.312* | 0.312** | -0.412* | 0.626* | 0.256** | 0.515 | 3.564* | 0.184 |
| *在0.05水平上显著相关;**在0.01水平上显著相关 | ||||||||||
在不同生境土壤螨类群落的多样性指标中个体密度、类群密度、多样性指数(H)、丰富度指数(M)与海拔、温度、有机质含量和全氮含量呈极显著正相关(P < 0. 01), 与土壤容重、pH、全磷、全钾及总盐量呈显著负相关(P < 0. 05)。相关分析结果表明, 海拔、温度和有机质含量对土壤螨类群落结构和多样性的影响显著。
3 讨论 3.1 不同生境土壤螨类群落结构的差异在不同生境土壤螨类个体数及类群数之间以及不同季节之间均有显著差异(P < 0.05), 与在我国其他区域研究结果基本一致, 其个体数为甲螨类>革螨类>辐螨类>粉螨类[18-26], 但也存在一定差异, 不同管理模式对土壤螨群落结构的影响均较小[24], 而耕作方式对土壤螨类数量和类群数存在显著影响[25]。对山地-绿洲\慌莫三大生态系统来讲, 个体密度及类群数在植被多样且有机质丰富的山地最高, 其次绿洲, 而荒漠最低。由于玛纳斯河流域海拔差异高、生境异质性强, 在山地生境植被丰富多样、腐殖层厚而人类干扰较小, 在绿洲生境虽植被多样、但腐殖层较薄、人类活动影响极为深刻, 相对而言荒漠生境干旱、植被稀少。土壤螨类的生态分布主要受生境类型、土壤理化性质和营养状况的制约[25-28]。
本研究表明不同生境土壤螨类群落的垂直分布呈明显的表聚性, 这与国内外同类研究结果一致[26-27]。土壤螨类的垂直分布差异与微环境土壤的理化性质(土壤含水量、pH、容重、温度及有机质)、植被以及营养状况的垂直分布差异密切相关[27-28]。土壤养分、湿度和容重是影响土壤螨类垂直分布的主要因素之一, 容重小, 土壤疏松, 土壤螨类类群数和个体数多, 在同一样地的土壤中, 随着土壤深度增加容重增大, 螨类类群数和个体数都减少。有机质和全氮对螨类的影响最大, 也有学者指出螨类的数量与土壤养分呈正相关[29-32]。温度对土壤螨类群落组成与个体数量也具有一定的影响, 土壤温度较低(≤ 10.5℃)时, 土壤螨类群落组成和个体数量减少, 而土壤温度在15—23℃之间时, 螨类群落组成和个体数量增多, 特别是捕食性螨类的增加明显[8, 18, 22]。
从4个季节调查结果来看, 在9月份的类群和个体数最高, 而7月份最低, 此结果与其他相关研究结果一致[23, 28]。也有研究表明, 土壤螨类在不同季节以9月最多, 而在11月最少[27-28], 这主要与区域温度及湿度的季节性变化密切相关。
3.2 不同生境土壤螨类群落多样性的比较生物多样性是群落生物组成结构的重要指标, 反映群落内物种的多少和生态系统食物网的复杂程度, 从而反映各生境间的相似性和差异性。采用多样性指数分析土壤螨类群落表明, 不同生境土壤螨类群落多样性指标之间具有显著性差异, 在Ⅰ生境最高、在Ⅻ生境最低, 与其他学者调查结果基本一致[24-26]。组成简单、类群数较低的群落可能比组成丰富数量较高的多样性指数更高, 这由研究地区地理环境以及气候条件不同所造成的[29-30]。生物群落生物多样性越大, 代表采样区域的生态系统的结构越稳定, 反之亦然。研究表明, pH值和含水量是影响土壤螨类群落的重要因素, 其中含水量是影响土壤螨类种类组成与多样性的关键因子[32-33], 含水量较高的土壤环境中螨类的群落组成较多, 个体数大, 生物多样性也高, 否则不利于土壤螨类的生长繁殖。含盐量和pH值是土壤螨类分布的限制因素, 大多数土壤螨类适应于微酸性或中性条件下生存[34]。本研究表明, pH值为6.8—7.8之间防护林生境的土壤螨类群落组成较为丰富, 而在pH值≤8.5的草地及荒漠生境土壤螨类群落组成和数量都比较少。本文相似性分析结果显示不同生境间的土壤螨类群落属于中等不相似或者极不相似, 表现出该区域所选生境类型之间差别很大、代表性强, 因而具有研究意义。总之, 土壤螨类群落结构及多样性变化在很大程度上受区域诸多环境因素(生物及非生物之间扩散限制和环境筛选的调控作用、气候和水温等多种因素)的综合作用的影响[35-36]。
4 结论研究区域土壤螨类资源丰富, 土壤螨类种类和多样性指标在山盆复合生态系统不同垂直带生境间均存在显著差异, 尤其是优势种类均不相同。在绿洲生态系统中有67科88属的代表, Ceratozetes、Oribatula、Holaspulus及Aleuroglyphus 4属为优势类群。山地生态系统中有57科87属, Oppiella和Eupelops为优势类群, 而荒漠生态系统有45科57属, Passalozetes和Liacarus为优势类群, 其中Passalozetes为沙漠环境特有分布的种类, 具有重要的环境指示作用。
| [1] | 王宗英, 路有成, 王慧芙. 九华山土壤螨类的生态分布. 生态学报, 1996, 16(1): 58–64. |
| [2] | 吴东辉, 张柏, 卜照义, 陈鹏. 长春市不同土地利用生境土壤螨类群落结构特征. 生态学报, 2006, 26(1): 16–25. |
| [3] | Fischer B M, Schatz H. Biodiversity of oribatid mites (Acari:Oribatida) along an altitudinal gradient in the Central Alps. Zootaxa, 2013, 3626: 429–454. DOI:10.11646/zootaxa.3626.4 |
| [4] | 吴东辉, 尹文英, 杨振明. 松嫩草原中度退化草地不同植被恢复方式下土壤螨类群落特征的差异. 动物学报, 2007, 53(4): 607–615. |
| [5] | Gulvik M E. Mites (Acari) as indicators of soil biodiversity and land use monitoring:a review. Polish Journal of Ecology, 2007, 55(3): 415–440. |
| [6] | 殷秀琴, 宋博, 董炜华, 辛未冬. 我国土壤动物生态地理研究进展. 地理学报, 2010, 65(1): 91–102. DOI:10.11821/xb201001010 |
| [7] | 郑经鸿, 王丽真, 张兴亚, 王新华, 薄新文. 新疆草地甲螨分布规律研究. 生态学报, 1993, 13(2): 171–176. |
| [8] | 吾玛尔·阿布力孜, 阿布都如苏力·吐逊, 木卡热木·阿吉木, 张卫红. 乌鲁木齐地区不同生境土壤捕食性螨类群落结构及其多样性研究. 新疆农业科学, 2012, 49(9): 1748–1758. DOI:10.6048/j.issn.1001-4330.2012.09.028 |
| [9] | 吾玛尔·阿布力孜, 阿布都如苏力·吐孙, 木开热木·阿吉木, 吴松林. 准噶尔盆地东南缘绿洲-荒漠交错带土壤螨类的群落多样性与相似性. 干旱区资源与环境, 2013, 27(6): 75–79. |
| [10] | 阿布都如苏力·吐孙, 吾玛尔·阿布力孜, 木开热木·阿吉木. 准噶尔盆地东南缘绿洲-荒漠交错带土壤螨类群落特征初步研究. 干旱区资源与环境, 2013, 27(3): 160–166. |
| [11] | 吾玛尔·阿布力孜, 阿提坎木·吾布力喀斯木, 吴松林. 新疆喀纳斯国家级自然保护区土壤螨类群落特征. 干旱区资源与环境, 2015, 29(2): 99–104. |
| [12] | 刘金巍, 靳甜甜, 刘国华, 李宗善, 杨荣金. 新疆玛纳斯河流域2000-2010年土地利用/覆盖变化及影响因素. 生态学报, 2014, 34(12): 3211–3223. |
| [13] | 尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998: 163–242. |
| [14] | 江原昭三. 日本蜱螨类检索图鉴. 东京: 全国农村教育协会出版社, 1980: 1–562. |
| [15] | 青木淳一. 日本土壤动物检索图鉴. 东京: 东海大学出版社, 1999: 173–436. |
| [16] | Balogh J, Balogh P. The Oribatid Mites Genera of the world (Vol.1 & 2). Budapest: The Hungarian National Museum Press, 1992: 1–263. |
| [17] | Krantz G W, Walter D E. A Manual of Acarology.3rd ed. Texas: Texas Tech University Press, 2009: 1–806. |
| [18] | Omar A, Ajar Q, Mukaram H, Wu S L. Community diversity and its seasonal dynamics of soil mites in Oasis of the Sangong river watershed of Xinjiang, China. Journal of Xinjiang University:Natural Science Edition, 2016, 33(1): 6–19. |
| [19] | 陈鹏, 文在根, 青木淳一, 芝, 石川和男. 长春净月潭地区土壤螨类的调查研究. 动物学报, 1988, 34(3): 282–293. |
| [20] | 吴东辉, 张柏, 陈鹏. 吉林省中西部平原区土壤螨类群落结构特征. 动物学报, 2005, 51(3): 401–412. |
| [21] | 吴东辉, 张柏, 安艳芬. 吉林省中部黑土区农业土地利用方式对土壤螨类群落特征的影响. 土壤通报, 2006, 37(1): 121–125. |
| [22] | 吴东辉, 尹文英, 陈鹏. 刈割条件下松嫩平原碱化羊草草地土壤螨类群落变化特征的研究. 土壤学报, 2008, 45(5): 1007–1014. |
| [23] | 丁程成, 戴征凯, 薛晓峰, 李辉信, 刘满强, 陈小云, 周静, 张斌, 胡锋. 退化红壤植被恢复对土壤螨类群落结构的影响. 生态学报, 2008, 28(10): 4771–4781. DOI:10.3321/j.issn:1000-0933.2008.10.019 |
| [24] | 李钰飞, 郑春燕, 李吉进, 孙钦平, 邹国元, 刘本生, 许俊香, 高利娟, 刘东生, 李季. 不同管理模式对温室土壤螨群落结构的影响. 生态学报, 2016, 36(12): 3802–3811. |
| [25] | 战丽莉, 许艳丽, 张兴义, 裴希超, 赵刚, 王建华. 耕作方式对土壤螨类群落结构的影响. 中国生态农业学报, 2013, 21(5): 598–605. |
| [26] | 阿提坎·吾布力喀斯木, 吾玛尔·阿布力孜, 努尔合尼穆·艾麦提. 玛纳斯河流域平原区土壤螨类群落特征研究. 安徽农业科学, 2014, 42(23): 7829-7834, 7843-7843. |
| [27] | Lin L, Gao M X, Liu D, Zhang X P, Wu H T, Wu D H. Co-occurence patterns of above-ground and below-ground mite communities in farmland of Sanjiang plain, northeast China. Chinese Geographical Science, 2014, 24(3): 339–347. DOI:10.1007/s11769-014-0683-4 |
| [28] | 高梅香, 刘冬, 张雪萍, 吴东辉. 三江平原农田地表和地下土壤螨类丰富度与环境因子的空间关联性. 生态学报, 2016, 36(6): 1782–1792. |
| [29] | Shakir M M, Ahmed S. Seasonal abundance of soil arthropods in relation to meteorological and edaphic factors in the agroecosystems of Faisalabad, Punjab, Pakistan. International Journal of Biometeorology, 2015, 59(5): 605–616. DOI:10.1007/s00484-014-0874-9 |
| [30] | Bolger T, Arroyo J J, Kenny J, Caplice M. Hierarchical analysis of mite community structures in Irish forests-A study of the relative contribution of location, forest type and microhabitat. Applied Soil Ecology, 2014, 83: 39–43. DOI:10.1016/j.apsoil.2013.06.004 |
| [31] | Dirilgen T, Arroyo J, Dimmers W J, Faber J, Stone D, da Silva P M, Carvalho F, Schmelz R, Griffiths B S, Francisco R, Creamer R E, Sousa J P, Bolger T. Mite community composition across a European transect and its relationships to variation in other components of soil biodiversity. Applied Soil Ecology, 2016, 97: 86–97. DOI:10.1016/j.apsoil.2015.06.008 |
| [32] | 韩雪梅, 李丹丹, 梁子安, 陈云峰, 胡诚. 北方常见农业土地利用方式对土壤螨群落结构的影响. 生态学报, 2013, 33(16): 5026–5034. |
| [33] | Kardol P, Reynolds W N, Norby R J, Classen A T. Climate change effects on soil microarthropod abundance and community structure. Applied Soil Ecology, 2011, 47(1): 37–44. DOI:10.1016/j.apsoil.2010.11.001 |
| [34] | Klimek A, Rolbiecki S, Rolbiecki R. Impact of ectohumus application in birch and pine nurseries on the presence of soil mites (Acari), Oribatida in particular. Folia Forestalia Polonica, Series A, 2016, 58(1): 20–30. |
| [35] | Gao M X, He P, Zhang X P, Liu D, Wu D H. Relative roles of spatial factors, environmental filtering and biotic interactions in fine-scale structuring of a soil mite community. Soil Biology and Biochemistry, 2014, 79: 68–77. DOI:10.1016/j.soilbio.2014.09.003 |
| [36] | 张丽梅, 高梅香, 刘冬, 张雪萍, 吴东辉. 环境筛选和扩散限制在地表和地下螨群落物种共存中的调控作用. 生态学报, 2016, 36(13): 3951–3959. |