 2017, Vol. 37
2017, Vol. 37文章信息
- 鲁蕾, 吴亦潇, 张维昊
- LU Lei, WU Yixiao, ZHANG Weihao.
- 城市小型湖泊浮游植物群落结构特征及演替规律——以武汉沙湖为例
- Distribution and succession pattern of phytoplankton communities in a small urban lake, Sand Lake in Wuhan City, China
- 生态学报. 2017, 37(18): 5993-6004
- Acta Ecologica Sinica. 2017, 37(18): 5993-6004
- http://dx.doi.org/10.5846/stxb201606211215
-
文章历史
- 收稿日期: 2016-06-21
- 网络出版日期: 2017-04-25

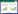




2. 水资源安全保障湖北省协同创新中心, 武汉 430072
2. Hubei Collaborative Innovation Center for Water Resource Security, Wuhan 430072, China
近年来, 城市湖泊成为各界关注的热点之一。其中, 城市小型湖泊由于其具有易受城市生态系统干扰的特性, 受到了广泛关注。研究者发现人类通过农业和气候对湖泊造成持续干扰, 会使浮游植物群落受到明显影响[1];同时, 研究表明浮游植物群落的初级生产力变化与水体生态系统的稳定性有相互影响的关系[2, 3]。因此, 城市小型湖泊浮游植物群落结构特征和演替规律的研究对于其生态保护有重要的意义。
在小型湖泊中易发生多种污染, 小型湖泊浮游动植物群落对有机污染物的抗性与群落结构有明显的相关性, 均呈现出季节性变化的特点[4], 群落中的物种变化可能比表观水体性质参数如叶绿素浓度、总磷要更敏感[5]。目前对于氮磷营养盐与浮游植物群落相关性的研究较多[6], 但是湖泊水体成分较为复杂, 除氮磷外, 浮游植物群落结构变化与水体中的其他组分存在一定关系, Tilman [7]等认为常量和微量金属元素在一些情况下会成为浮游植物生长的限制因素;Holopainen[8]等研究发现矿地废水流入造成阳离子浓度显著升高的浅水湖泊中, 浮游植物群落生物量达到了对照无污染湖泊的两倍。因此对于水体中的常量离子对浮游植物群落的影响应该给予关注。
目前, 富营养化是城市小型湖泊面临的主要环境问题。在富营养化湖泊中, 营养盐不再是限制因素, 浮游动植物群落结构和群落内种间关系成为群落变化的原因[9]。优势种和优势类群很大程度上可以成为湖泊水质的评判标准[10], 并具有一定的时序变化性[11]。目前研究者普遍进行的是群落物种表观数量关系研究[12-13], 优势种种间关联研究缺乏。而优势种作为群落结构的指示, 其种间关联对群落演替具有潜在的驱动作用, 并有较高的研究价值。
综上所述, 本研究以沙湖为例, 研究了典型富营养化城市小型湖泊浮游植物群落的时空异质性, 并从环境因素和优势种更替两个方面探究浮游植物群落的演替规律。其中, 环境因子通过参与复杂的生化反应影响群落的演替过程(即群落演替外驱动力), 而优势种更替则直接影响群落的演替方向(即群落演替内驱动力)。
1 实验方法 1.1 采样点设置采样区域为生态功能处于退化状态的城市小型湖泊——沙湖, 位于武汉市武昌东北部, 水域面积3.197km2, 于2013年完成一期公园建设, 通过人工挖掘的楚河与东湖连通。设置的采样点见图 1。

|
| 图 1 研究区采样点设置 Fig. 1 Sample location of area under study |
采样点坐标见表 1。采样点分布的湖区存在明显差异, 5号采样点为码头;3、4号采样点位于挺水植物生长区边缘;1、2、6号采样点邻近公园, 其中1号点邻近公园道路, 2号点邻近公园树林, 6号点邻近公园草坡。
| 采样点Sample site | 1 | 2 | 3 | 4 | 5 | 6 |
| 经度Longitude/(°E) | 114.320 | 114.329 | 114.339 | 114.335 | 114.327 | 114.319 |
| 纬度Altitude/(°N) | 30.570 | 30.573 | 30.574 | 30.567 | 30.565 | 30.562 |
由于武汉1、2月多雨雪天气, 水温偏低, 表层浮游植物群落采集困难, 因此设计的浮游植物群落采集时间为2015年3—12月, 样品采集使用1L有机玻璃采水器, 采集表层0.5m处水样, 使用鲁哥氏液进行固定浓缩后, 于高倍显微镜下使用0.1mL计数框分类计数。
常量阳离子监测时间段为2015年5—12月。阳离子浓度测定使用EP-600便携式离子色谱仪, 阳离子保护柱:Thermo scientific Dionex IonPacTM CG12A RFICTM Guard 4×50 mm, 阳离子交换柱:Thermo scientific Dionex IonPacTM CG12A RFICTM Analytical 4×250 mm, 流动相:18mmol/L甲烷磺酸。
1.3 多样性指数计算4个多样性指数, 分别为Shannon-Wiener指数、Simpson指数、Pielou均匀度指数[14]和Whittaker指数[15]。计算公式如下:
Shannon-Wiener指数:

|
(1) |
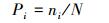
|
(2) |
Simpson指数:

|
(3) |
Pielou均匀度指数:
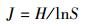
|
(4) |
Whittaker指数:
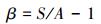
|
(5) |
式中, S为每月采样出现的物种总数, A为每月各采样点的平均物种数, Pi为第i种的相对重要值, ni为采样点第i种物种的个体数, N为采样点各物种的总个体数。H值范围大于等于3为清洁;2—3为轻度污染;1—2为中度污染;小于1为重污染[16]。J值范围0—1时, J值大体现种间个体分布均匀, 污染程度增加J值减小[17]。
1.4 优势种与种间联结性计算优势种使用优势度进行判断[17], 种间联结性选取了3个联结性指数, Ochiai指数、Dice指数和Jaccard指数, 以及校正χ2检验系数[19]进行描述。
优势度:
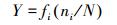
|
(6) |
Ochiai指数:
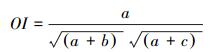
|
(7) |
Dice指数:

|
(8) |
Jaccard指数:
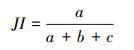
|
(9) |
校正χ2检验系数:
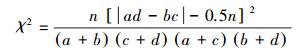
|
(10) |
式中, fi为物种i的出现频率;a为两种物种均出现的采样月数, b、c为只有其中一种物种出现的采样月数, d为两种物种均未出现的采样月数, n为采样总月数。取Y≥0.02为优势种[17]。当ad>bc时为正联结, 当ad < bc时为负联结;当χ2>3.841时为显著联结, 当χ2>6.635时为极显著联结[18]。
1.5 数据分析数据分析使用软件Matlab2014a, 聚类分析使用K-均值法[19]进行计算。数据绘图使用Originlab 8.5。
2 结果 2.1 浮游植物群落时空异质性 2.1.1 群落结构变化规律沙湖浮游植物群落的物种组成和相对丰度均表现出明显的季节性。
从群落物种组成来看(图 2), 2015年3—12月, 绿藻门(Chlorophyta)和硅藻门(Bacillariopgyta)物种数超过沙湖的浮游植物群落总体的70%;其中, 春季硅藻门物种数超过总体的30%, 夏秋季绿藻门物种数超过总体的50%;蓝藻门(Cyanophyta)物种数占比范围为7%—14%, 5—7月以及11—12月蓝藻门物种数占比超过10%;裸藻门(Euglenophyta)物种数占比(3%—9%)呈现波形变化, 5月达最小值, 8月、12月达到最大值;黄藻门(Xanthophyta)物种数占比(3%—6%)4月达最小值, 8月达最大值, 其他各月占比稳定。

|
| 图 2 不同采样月沙湖浮游植物群落物种组成 Fig. 2 Taxonomic composition of phytoplankton in different sampling months in Sand Lake |
由群落相对丰度变化可知(图 3), 2015年3—12月绿藻门占比均超过总丰度的50%;蓝藻门(1%—20%)和硅藻门(9%—30%)生物量比例呈波形变化, 最大值分别出现在8月和4月, 最小值分别出现在3月和6月;裸藻门占比大幅增加出现在4月和9—10月, 黄藻门占比大幅增加出现在5—8月;甲藻门(Dinophyta)在3月、金藻门(Crysophyceae)在12月比重明显增大。隐藻门(Cryptophyta)、定鞭藻门(Haptophyta)和红藻门(Rhodophyta)物种数和相对丰度占群落比例均非常小, 为稀有种或偶见种。

|
| 图 3 不同采样月沙湖浮游植物群落物种相对丰度 Fig. 3 Relative abundance of phytoplankton in different sampling months in Sand Lake |
(1)α和β多样性时间异质性
β多样性指数为Whittaker指数;其值降低说明生境异质性增加[17]。由图 4(a)可知, β多样性分布有明显季节性。沙湖各采样点浮游植物群落的异质性在7—9月最高, 此时间段, 采样点内的物种数较多而采样点间的物种差异最大, 群落处于物种更新缓慢的稳定状态;3—7月群落异质性逐渐增强, 表明此时段沙湖整体浮游植物群落在向稳定状态演替;9—12月群落异质性逐渐下降, 表明此时段浮游植物群落开始向不稳定状态演替;12月浮游植物群落分布为监测时段最均匀的月份(Whittaker指数达最大值), 说明此时沙湖各采样点的共有物种明显减少, 整体物种更新速率加快。

|
| 图 4 监测时段沙湖浮游植物群落α、β多样性指数 Fig. 4 α and β diversity indexes of phytoplankton in Sand Lake from March to December |
α多样性指数分别为Shannon-Wiener指数H、Simpson指数D和Pielou均匀度指数J;其中, H值和J值反映水体受污染的状态, H值与丰富度的关系最密切, D值和J值对均匀度的敏感性较高[14]。由图 4(b)可知, α多样性呈现明显波形变化规律。总体来说, 监测时段内H值均集中在1—2之间, J值大都集中在0.3—0.4之间, 说明水体保持在中度污染状态;3、6、12月J值低于0.3说明此时水体处于重污染状态。α多样性分布规律β多样性基本一致, 7—9月沙湖的平均α多样性指数最高, 说明此时各采样点群落物种丰富度和均匀度均达到稳定状态;3—7月为α多样性增加段, 9—12月为α多样性下降段, 总的来说, 3—12月沙湖浮游植物群落的演替规律为由不稳定向稳定再向不稳定群落发展的过程。
(2) 浮游植物群落空间异质性
分别以Shannon-Wiener指数、Simpson指数和Pielou均匀度指数为每月每采样点的x, y, z坐标进行聚类分析(图 5), 最优得到4类, 各类中心信息见表 2。

|
| 图 5 监测时段沙湖浮游植物群落三种α多样性聚类图 Fig. 5 Cluster result of three kinds of α diversity indexes of phytoplankton in Sand Lake in monitoring period |
| 中心点类别 Center points of clusters |
坐标Coordinates | 中心点类别 Center points of clusters |
坐标Coordinates | ||||
| x | y | z | x | y | z | ||
| 类别1 cluster1 | 1.353 | 0.915 | 0.327 | 类别3 cluster3 | 1.479 | 0.941 | 0.344 |
| 类别2 cluster2 | 1.215 | 0.876 | 0.301 | 类别4 cluster4 | 0.927 | 0.748 | 0.236 |
由表 2可知, 4类的综合α多样性排序为:3>1>2>4。对监测时段各采样点α多样性根据聚类进行分类可知(表 3), 沙湖群落存在明显的时空异质性。从空间分布来看, 2号点α多样性最高, 群落状态最稳定;1、3号点以及4、6号点差异不大, 其群落演替状态较接近;5号点α多样性最低, 群落受到扰动最大;总体来看, 沙湖的浮游植物群落空间异质性表现为湖泊中段群落较两侧稳定, 同一侧湖岸的两端群落演替状态相似, 湖泊中人类频繁活动区域群落多样性最低。
| 月份Month | 采样点Sample site | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 3 | 2 | 3 | 2 | 2 | 4 | 4 |
| 4 | 2 | 3 | 2 | 2 | 2 | 2 |
| 5 | 1 | 3 | 2 | 2 | 1 | 2 |
| 6 | 4 | 2 | 2 | 2 | 1 | 2 |
| 7 | 1 | 1 | 1 | 1 | 1 | 3 |
| 8 | 3 | 3 | 3 | 1 | 1 | 3 |
| 9 | 1 | 3 | 1 | 1 | 1 | 1 |
| 10 | 1 | 1 | 1 | 1 | 1 | 1 |
| 11 | 1 | 2 | 1 | 2 | 2 | 1 |
| 12 | 1 | 1 | 2 | 2 | 2 | 4 |
本研究考虑的环境因子包括温度、钠、钾、镁、钙的阳离子浓度, 除温度外环境因子测定时间为5—12月。由图 6可知, 水质参数结果分布呈明显季节性, 总体上为3—7月逐渐增大, 9—12月逐渐减小, 8月出现夏季最低值。钙离子浓度明显高于其他3种离子, 9月各离子浓度达到最大值, 7月温度达到最大值, 4—10月表层水温度高于20℃, 12月水温低于10℃。

|
| 图 6 监测时段沙湖水质因子变化图 Fig. 6 The results of water quality factors of Sand Lake from March to December |
(1) 多样性与环境因子相关性分析
由表 4可知, Whittaker指数与钾、钙离子浓度和温度有极强负相关关系, 说明当其浓度增大或温度增加, 各采样点间的异质性增强。Shannon-Wiener指数与镁离子浓度和温度有强相关关系, 与钠离子浓度有极强相关关系;Simpson指数与钠离子浓度有强相关关系, 与镁离子浓度有极强相关关系;Pielou均匀度指数与钠、镁离子浓度有强相关关系。相关性分析结果说明, 环境因子对浮游植物群落状态有明显的影响, 其中常量离子浓度对群落演替有比温度更强的影响力, 钠、镁离子浓度是α多样性的主要影响因素, 温度主要影响群落物种丰富度, 钾、钙离子浓度主要影响沙湖浮游植物群落β多样性。
| 相关系数 R |
Pielou均匀 度指数 |
Simpson 指数 |
Shannon-Wiener 指数 |
Whittaker 指数 |
钠离子 Na |
钾离子 K |
镁离子 Mg |
钙离子 Ca |
温度 Temperature |
| Pielou均匀度指数 | 1 | - | |||||||
| Simpson指数 | 0.882 | 1 | - | ||||||
| Shannon-Wiener指数 | 0.796 | 0.913 | 1 | - | |||||
| Whittaker指数 | -0.319 | -0.475 | -0.729 | 1 | - | ||||
| 钠离子Na | 0.663 | 0.771 | 0.811 | -0.785 | 1 | - | |||
| 钾离子K | 0.176 | 0.357 | 0.585 | -0.932 | 0.718 | 1 | - | ||
| 镁离子Mg | 0.781 | 0.819 | 0.744 | -0.555 | 0.943 | 0.471 | 1 | - | |
| 钙离子Ca | 0.376 | 0.470 | 0.599 | -0.885 | 0.878 | 0.882 | 0.744 | 1 | - |
| 温度Temp. | 0.085 | 0.379 | 0.640 | -0.893 | 0.579 | 0.907 | 0.302 | 0.681 | 1 |
(2) 主成分分析
主成分分析考虑的因素包括α多样性指数与环境因子。得到的主成分中, 主成分1、主成分2和主成分3分别能够解释60.01%、24.44%和11.28%(累积95.73%)的数据量(表 5)。根据监测数据对应3个主成分空间的变换值进行聚类, 最优可得3类结果, 对3类结果进行整理见表 6, 主成分分析聚类结果见图 7。
| 主成分 Principal component |
Shannon- Wiener指数 |
Simpson 指数 |
Pielou均匀 度指数 |
钠离子 Na |
钾离子 K |
镁离子 Mg |
钙离子 Ca |
温度 Temperature |
| 主成分1 PC1 | 0.364 | 0.329 | 0.298 | 0.416 | 0.347 | 0.375 | 0.375 | 0.309 |
| 主成分2 PC2 | 0.349 | 0.450 | 0.511 | -0.138 | -0.399 | 0.00806 | -0.340 | -0.347 |
| 主成分3 PC3 | -0.328 | -0.171 | 0.00974 | 0.344 | -0.273 | 0.577 | 0.202 | -0.545 |
| 月份Month | 采样点Sample site | |||||
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 5 | 1 | 1 | 1 | 2 | 1 | 1 |
| 6 | 2 | 2 | 2 | 2 | 2 | 2 |
| 7 | 3 | 3 | 3 | 3 | 3 | 3 |
| 8 | 3 | 3 | 3 | 3 | 3 | 3 |
| 9 | 3 | 3 | 3 | 3 | 3 | 3 |
| 10 | 3 | 3 | 3 | 3 | 3 | 3 |
| 11 | 1 | 1 | 1 | 1 | 1 | 1 |
| 12 | 1 | 1 | 1 | 1 | 1 | 2 |
| 每一类中心为:类别1(-1.642, 1.154, 0.227), 类别2(-2.384, -1.847, -0.678), 类别3(1.889, -0.154, 0.0745) | ||||||

|
| 图 7 监测时段沙湖生物多样性与水质因子主成分分析聚类图 Fig. 7 Cluster results of principal component analysis of biodiversity indexes and water quality factors from March to December 主成分3对应的数值为原数值减去最小值, 以使点均落在z轴正向; 蓝色点为类别1, 绿色点为类别2, 红色点为类别3 |
主成分分析是从原本存在相关性的各成分中提取出同样个数的完全线性无关的主成分进行分析的过程。由表 5可知, 主成分1综合性较强;主成分2多样性指数为主要因子;主成分3镁离子浓度、温度为主要因子。
由表 6类别中心可知, 类别3主成分1为主要因素, 该类点α多样性和环境因子数值均较高;类别2处于3个成分的负象限, 说明该类点α多样性和环境因子数值均较低;类别1主成分1和2为主要因素, 主成分1为负象限, 说明该类点的α多样性较高而离子浓度等环境因子数值较低。由表 6分可知, 对各采样点位来说, 6月均为2类, 7—10月均为3类, 此时段内群落状态明显受到环境因子的驱动, 表现为两者分布趋势一致;5、11、12月大多数采样点为1类, 说明此时段内, 除了环境因子的影响外, 可能存在其他作用较强的影响因素。结合表 6和表 3的聚类结果可知, 夏秋季节群落状态较稳定, 环境因子有明显的驱动性, 冬春季节群落处于演替状态, 环境因子的驱动力减弱, 内驱动力可能为主要作用因素。
2.3 浮游植物群落演替内驱动力 2.3.1 优势种变化规律每月的优势种由优势度确定, 3—12月出现的优势种共40种(表 7)。其中绿藻门21种, 硅藻门10种, 蓝藻门4种, 黄藻门和裸藻门各2种, 金藻门和甲藻门各1种。由表 7可知, 每月的优势种数量呈现先增多再减少的规律, 最大值为9月, 达到17种, 最小为3月, 有5种;其他各月基本上呈现出季节上的稳定性;监测时段主要优势种有5个, 为S9、S10、S17、S23、S33, 主要优势种集中在春季和夏季, 且均为绿藻。
| 编号 Species number |
中文种名 Species name in Chinese |
学名 Species name in Latin |
出现时间 Appearance time |
| S1 | 卷曲鱼腥藻 | Anabaena circinalis Rab. | 6, 7 |
| S2 | 针形纤维藻 | Ankistrodesmus acicularis (A. Braun) Korschikoff | 4 |
| S3 | 卷曲纤维藻 | Ankistrodesmus convolutus Corda | 9 |
| S4 | 四棘鼓藻 | Arthrodesmus convergens Ehrenberg ex Ralfs | 9—11 |
| S5 | 英克斯四棘鼓藻 | Arthrodesmus incus (Breb.) Hassall ex Ralfs | 10, 11 |
| S6 | 查氏四棘藻 | Attheya Zachariasi J.Brun. | 12 |
| S7 | 拟气球藻 | Botrydiopsis arhiza Borzi | 5 |
| S8 | 艾氏衣藻 | Chlamydomonas Ehrenbergii Gorosch. | 12 |
| S9 | 椭圆小球藻* | Chlorella ellipsoidea Gerneck | 3—12 |
| S10 | 蛋白核小球藻* | Chlorella pyrenoidosa Chick | 3—8, 10—12 |
| S11 | 土生绿球藻 | Chlorococcum humicola (Nag.) Rabenhorst | 11 |
| S12 | 华美色金藻 | Chromulina elegans Doflein | 12 |
| S13 | 网状空星藻 | Coelastrum reticulatum (Dang.) | 9 |
| S14 | 囊形柄裸藻 | Colacium vesiculosum Ehrenberg | 4, 10 |
| S15 | 双眼鼓藻 | Cosmarium bioculatum Brebisson | 9, 12 |
| S16 | 模糊鼓藻 | Cosmarium obsoletum (Hantzsch) Reinsch | 11 |
| S17 | 美丽鼓藻* | Cosmocladium pulchellum Breb. | 6—8, 10 |
| S18 | 湖北小环藻 | Cyclotella hubeiana Chen & Zhu | 3—5, 8, 10, 11 |
| S19 | 具盖小环藻 | Cyclotella operculata (A.g.) | 9 |
| S20 | 星芒小环藻 | Cyclotella stelligera Cl. et Grun. | 9 |
| S21 | 贪食拟裸藻 | Euglenopsis vorax Klebs. | 9 |
| S22 | 绿脆杆藻 | Fragilaria virescens Ralfs | 8 |
| S23 | 褶皱双胞藻* | Geminella crenulatocollis Prescott | 3, 6, 10—12 |
| S24 | 月状蹄形藻 | Kirchneriella lunaris (Kirchner) | 9 |
| S25 | 肥壮蹄形藻 | Kirchneriella obesa (W. West) Schmidle | 9 |
| S26 | 模糊直链藻 | Melosira ambigua (Grun.) | 12 |
| S27 | 颗粒直链藻 | Melosira granulata (Ehr.) Ralfs | 4, 9 |
| S28 | 意大利直链藻 | Melosira italica (Ehr.) Kutzing | 4 |
| S29 | 远距直链藻 | Melosira listans (Ehr.) | 9 |
| S30 | 变异直链藻 | Melosira varians Agardh | 9 |
| S31 | 马氏平裂藻 | Merismopedia marssonii Lemm. | 5—11 |
| S32 | 细小平裂藻 | Merismopedia minima G. Beck | 5, 7—11 |
| S33 | 集球藻* | Palmellococcus miniatus (Kutz.) Chodat | 7, 8 |
| S34 | 二角多甲藻 | Peridinium bipes Stein | 3 |
| S35 | 月牙藻 | Selenastrum bibraianum Reinsch | 6, 10, 11 |
| S36 | 小形月牙藻 | Selenastrum minutum (Nag.) Collinus | 10, 11 |
| S37 | 光角星鼓藻 | Staurastrum muticum (Breb.) Ralfs | 7, 10 |
| S38 | 锥形刺叉星鼓藻 | Staurodesmus subulatus (Kutz.) Croasdale | 5, 7 |
| S39 | 三角四角藻 | Tetraedron trigonum (Nag.) Hansgirg | 9, 10 |
| S40 | 普通黄丝藻 | Tribonema vulgare Pasch. | 9 |
| *:主要优势种 | |||
优势种每月分布的联结性计算结果见表 8。Ochiai、Dice、Jaccard指数确定的完全相关的物种分别为S4和S3、S20和S19、S21和S19、S21和S20、S25和S24、S30和S29;这6对物种同时作为优势种出现的时间为9月, 且其优势度均在0.02-0.04中间, 不是主要优势种。Jaccard指数强相关的物种分别为S9和S10, S32和S31;Dice指数强相关的物种有5对, 分别为S9和S10, S32和S31, 以及S27和S28、S33和S31、S39和S40;Ochiai指数强相关的物种有8对, 分别为S9和S10, S32和S31、S27和S28、S33和S31、S39和S40, 以及S33和S32、S11和S9、S29和S27。
| 联结性指数 Correlation coefficients |
0—0.2 | 0.2—0.4 | 0.4—0.6 | 0.6—0.8 | 0.8—1.0 | 1 |
| 落合系数Ochiai | 28.72% | 46.54% | 19.23% | 3.72% | 1.02% | 0.77% |
| 戴斯系数Dice | 33.72% | 47.82% | 14.10% | 2.95% | 0.64% | 0.77% |
| 杰卡德相似系数Jaccard | 70.00% | 25.26% | 2.82% | 0.90% | 0.26% | 0.77% |
校正χ2检验结果见表 9。χ2检验结果中显著正联结的物种有6对, 分别为S22和S19、S28和S27、S29和S27、S30和S27、S33和S31、S40和S39;极显著正联结的物种有7对, 分别为S4和S3、S20和S19、S21和S19、S21和S20、S25和S24、S30和S29以及S32和S31;不相关的结果中, S9的与编号在其后的优势种不相关, 2对不相关的物种分别为S26和S22、S39和S35。
| 联结性 Coefficient results |
正, 显著 Positive, significant |
正, 极显著 Positive, highly significant |
正, 不显著 Positive, nonsignificant |
负, 不显著 Negative, nonsignificant |
不相关 Uncorrelated |
| χ2 | 0.77% | 0.90% | 70.38% | 23.72% | 4.23% |
种间联结性测定值本身有一定的生态学意义, 在一定程度上衡量了物种的相互关系和物种对环境综合因子反应的差异[18];χ2检验能比较准确度量种对的关联显著程度[20]。3个种间联结性指数结果略有不同, Jaccard、Dice指数得到的信息量小于Ochiai指数。优势种种间联结性表现为同属相关性较强, 包括三个指数均为强相关的2种小球藻和2种平裂藻, 以及Ochiai和Dice指数强相关的直链藻, 不同属也存在强相关的现象, 包括三角四角藻和普通黄丝藻、绿球藻和椭圆小球藻;绿藻门和蓝藻门优势种显著相关, 包括集球藻和2种平裂藻;硅藻门和其他藻类相关性均较低, 表现为同属的高相关性。χ2检验结果和联结性指数计算结果基本一致, 但χ2检验结果出现不相关的物种, 包括模糊直链藻和绿脆杆藻、三角四角藻和月牙藻, 其中, χ2检验中椭圆小球藻与其他物种相关性均较低。
总体来说, 9月群落处于监测时段最稳定状态, 4月和9月优势种相关性较高, 夏季和秋季优势种相关性较高, 该结果与表 6和表 3结果一致, 说明群落物种结构的强关联性对夏秋季节沙湖浮游植物群落演替的稳定状态有一定贡献;5和11月优势种仅2种(S31, S32) 与7、8月优势种S33有强相关性, 其他优势种表现为月内优势种极强相关性(S9和S10) 和不与其他时段优势种相关的情况, 12月优势种和其他优势种相关性也普遍较低, 出现的是耐低温的物种;该结果与表 6结果一致, 说明5、11、12月浮游植物群落演替主要由群落内物种构成变化驱动。
3 讨论和结论 3.1 城市小型湖泊浮游植物群落时空分布异质性沙湖的浮游植物群落结构与分布存在明显的时空变化规律。沙湖是典型的富营养化湖泊, 主要优势类群为绿藻-硅藻-蓝藻(累积物种数占比超过80%), 研究者普遍认为优势类群的占比可以用于指示湖泊的污染状态[10], 硅藻的物种数占比从春季(约30%)到夏季(低于20%)有明显下降, 说明沙湖受污染程度从夏季开始加深;蓝藻、裸藻物种数占比在夏秋季达到最大值, 表明沙湖在夏秋季处于富营养化加剧的状态。整个监测时段中, 7—9月浮游植物群落整体α多样性最高, β多样性最低(图 4), 群落处于演替的稳定阶段, 该时段的群落表现为耐污种占主要(表 7), 各采样点在该时段群落多样性差异不大。α多样性较低的3—7月和9—12月, 多样性分布存在明显空间差异(表 3), 5号点码头的多样性总体来说最小, 表明人为扰动对小型湖泊生物多样性有明显的影响;刘春光[21]等认为水生植物能够通过多种机制影响湖泊生态系统, 3、4号点为人工种植挺水、沉水植物的水域边缘带, 由采样点间对比发现, 挺水、沉水植物对小型湖泊浮游植物群落的影响较小;6月各点生物多样性均明显下降, 而1号点多样性下降最明显, 由1、2、6号采样点对比结果可知, 小型湖泊的浮游植物群落的稳定性与湖滨带的环境有相关关系。
3.2 浮游植物群落演替外驱动力外驱动力主要作用因子为钠、镁离子浓度, 温度为辅助作用因子, 外驱动力主要作用时段为夏秋季节。由主成分分析结果可知, 6—10月α多样性与环境因子变化规律一致, 该时段内浮游植物群落主要驱动力为外驱动力, 可能原因为多样性较高的7—9月浮游植物群落处于较稳定状态, 物种替换减缓, 以物种更新为主的内驱动力减小, 环境因子的驱动作用相对增大;多样性较低的6月浮游植物群落处于较敏感状态, 加之水体整体营养水平在该时段较低[22], 环境因子成为群落更新生长的限制因素。通过浮游植物群落多样性和环境因子的相关性分析可知, 钠、镁离子浓度与群落α多样性有强相关性, 主要原因可能为镁离子是藻类的叶绿素的组成元素之一且与叶绿体基粒堆垛直接相关, 能够对叶绿体荧光产率和光合作用原初过程造成影响[23], 该结果与其他研究结果类似[24]。本研究发现温度与浮游藻类群落的Shannon-Wiener指数有强相关性, 但与Simpson指数相关性较低, 与Pielou均匀度指数基本无相关性, 与大中型湖泊水库研究结果[25, 26]有明显差异, 表明大中型湖泊与小型湖泊浮游植物群落演替外驱动力因素存在差异, 温度影响降低可能由于小型湖泊层间温度和湖区温度差异化不明显。Whittaker指数与Shannon-Wiener指数和其他环境因子均有强负相关关系, 说明较高的生境内多样性与较高的常量离子浓度有利于浮游植物群落维持演替的稳定状态。
3.3 浮游植物群落演替内驱动力浮游植物优势种种类及其数量对浮游植物群落结构的稳定性有重要影响, 优势种种类数越多且优势度越小, 则群落结构越复杂、稳定[26], 沙湖每月优势种数量变化表明其群落演替规律为由不稳定到稳定再逐步向不稳定变化的过程。从优势种分类来看, 硅藻和蓝绿藻对环境因素的响应存在明显差异[27]。蓝藻大量生长的时间段为夏季和冬季, 既有相对生物量的增加, 也有种类数的明显增加;夏季爆发可能由于该季节水体状态更适宜蓝藻生长[28], 冬季爆发可能由于2015年11月降雨量增大, 出现适宜蓝藻生长的面源输入[29]。硅藻大量生长的时间为春季和秋季, Sarmento[30]等通过对于浅水湖泊的研究发现, 外源输入减少水位降低时, 硅藻生物量得到明显增加;硅藻夏季种类数比重变化稳定, 但是生物量比重明显下降, 硅藻与蓝藻占总密度百分比为极显著负相关[31]。硅藻和蓝藻优势种的负关联性可能是5、11月浮游植物群落结构演替与夏秋季不同的主要驱动因素。甲藻的生物量最大值出现在3月, 达到6%, 胡韧[32]等研究发现, 在温度不是主要的限制因素时, 春季甲藻的生物量较高, 与硅藻门和绿藻门构成优势类群;12月湖水温度低于10℃, 优势种以金藻门为主, 其特点为耐低温性, 其与其他优势种几乎无种间关联, 可能为冬季群落演替的主要驱动力。
| [1] | Lemma B. Ecological changes in two ethiopian lakes caused by contrasting human intervention. Limnologica-Ecology and Management of Inland Waters, 2003, 33(1): 44–53. DOI:10.1016/S0075-9511(03)80006-3 |
| [2] | Mukankomeje R, Plisnier P D, Descy J P, Massaut L. Lake muzahi, rwanda: limnological features and phytoplankton production. Hydrobiologia, 1993, 257(2): 107–120. DOI:10.1007/BF00005951 |
| [3] | Sommer U, Sommer F. Cladocerans versus copepods: The cause of contrasting top-down controls on freshwater and marine phytoplankton. Oecologia, 2006, 147(2): 183–194. DOI:10.1007/s00442-005-0320-0 |
| [4] | Willis K J, Van den Brink P J, Green J D. Seasonal variation in plankton community responses of mesocosms dosed with pentachlorophenol. Ecotoxicology, 2004, 13(7): 707–720. DOI:10.1007/s10646-003-4430-3 |
| [5] | Huszar V L M, Silva L H S, Domingos P, Marinho M, Melo S. Phytoplankton species composition is more sensitive than OECD criteria to the trophic status of three Brazilian tropical lakes. Hydrobiologia, 1998, 369: 59–71. |
| [6] | 谭香, 夏小玲, 程晓莉, 张全发. 丹江口水库浮游植物群落时空动态及其多样性指数. 环境科学, 2011, 32(10): 2875–2882. |
| [7] | Tilman D, Kilham S S, Kilham P. Phytoplankton community ecology:the role of limiting nutrients. Annual Review of Ecology and Systematics, 1982, 13(1): 349–372. DOI:10.1146/annurev.es.13.110182.002025 |
| [8] | Holopainen I J, Holopainen A L, Hämäläinen H, Rahkola-Sorsa M, Tkatcheva V, Viljanen M. Effects of mining industry waste waters on a shallow lake ecosystem in karelia, north-west russia. Hydrobiologia, 2003, 506(1/3): 111–119. |
| [9] | Mataloni G, Tesolín G, Sacullo F, Tell G. Factors regulating summer phytoplankton in a highly eutrophic antarctic lake. Hydrobiologia, 2000, 432(1/3): 65–72. DOI:10.1023/A:1004045219437 |
| [10] | 庞科, 姚锦仙, 王昊, 刘松涛, 李翀, 吕植. 额尔古纳河流域秋季浮游植物群落结构特征. 生态学报, 2011, 31(12): 3391–3398. |
| [11] | 田永强, 俞超超, 王磊, 黄邦钦. 福建九龙江北溪浮游植物群落分布特征及其影响因子. 应用生态学报, 2012, 23(9): 2559–2565. |
| [12] | 王丽卿, 施荣, 季高华, 范志锋, 程婧蕾. 淀山湖浮游植物群落特征及其演替规律. 生物多样性, 2011, 19(1): 48–56. |
| [13] | 王静雅, 汪志聪, 李翀, 叶少文, 连玉喜, 刘家寿, 张堂林, 李钟杰. 三峡水库坝前水域浮游植物群落时空动态研究. 水生态学杂志, 2015, 39(5): 877–884. |
| [14] | 许晴, 张放, 许中旗, 贾彦龙, 尤建民. Simpson指数和Shannon-Wiener指数若干特征的分析及"稀释效应". 草业科学, 2011, 28(4): 527–531. |
| [15] | 周洪华, 陈亚宁, 李卫红. 塔里木河下游绿洲——荒漠过渡带植物多样性特征及优势种群分布格局. 中国沙漠, 2009, 29(4): 688–696. |
| [16] | 刘文盈, 张秋良, 高润宏, 铁牛, 崔琥. 盐沼湿地浮游植物多样性与环境关系研究. 内蒙古林业科技, 2012, 38(1): 5-7, 22-22. |
| [17] | 郝媛媛, 孙国钧, 张立勋, 龚雪平, 许莎莎, 刘慧明, 张芬. 黑河流域浮游植物群落特征与环境因子的关系. 湖泊科学, 2014, 26(1): 121–130. DOI:10.18307/2014.0115 |
| [18] | 徐敏, 高翔, 吕光辉, 傅德平, 杨晓东. 克拉玛依典型植物群落种间关联分析. 新疆农业科学, 2010, 47(5): 1041–1046. DOI:10.6048/j.issn.1001-4330.2010.05.037 |
| [19] | 周世兵, 徐振源, 唐旭清. 新的k-均值算法最佳聚类数确定方法. 计算机工程与应用, 2010, 46(16): 27–31. |
| [20] | 陈家长, 孟顺龙, 尤洋, 胡庚东, 瞿建宏, 吴伟, 范立民, 马晓燕. 太湖五里湖浮游植物群落结构特征分析. 生态环境学报, 2009, 18(4): 1358–1367. |
| [21] | 刘春光, 邱金泉, 王雯, 庄源益. 富营养化湖泊治理中的生物操纵理论. 农业环境科学学报, 2004, 23(1): 198–201. |
| [22] | Li H M, Tang H J, Shi X Y, Zhang C S, Wang X L. Increased nutrient loads from the Changjiang (Yangtze) River have led to increased Harmful Algal Blooms. Harmful Algae, 2014, 39: 92–101. DOI:10.1016/j.hal.2014.07.002 |
| [23] | 孙谷畴. 镁离子对柱孢鱼腥藻(Anabaena cylindrica)类囊体膜吸收光谱和荧光光谱的影响. 植物生理学报, 1983, 9(4): 339–345. |
| [24] | 刘文盈, 铁牛, 张秋良, 高润宏. 藻类在盐沼湿地分布的关键限制因子研究. 内蒙古林业科技, 2012, 38(4): 16-18, 26-26. |
| [25] | 邓文丽, 刘均平, 王晓星, 杜桂森, 洪剑明. 北京野鸭湖浮游植物群落结构与水质关系研究. 湿地科学, 2013, 11(1): 27–34. |
| [26] | 裴国霞, 郭琦, 魏乐, 李汗青. 金海水库浮游植物调查及群落结构研究. 水生态学杂志, 2014, 35(2): 68–73. |
| [27] | 王宇飞, 赵秀兰, 何丙辉, 黄琪. 汉丰湖夏季浮游植物群落与环境因子的典范对应分析. 环境科学, 2015, 36(3): 922–927. |
| [28] | 郑吴柯, 刘宪斌, 赵兴贵, 陈曦. 于桥水库浮游植物群落特征. 中国环境监测, 2015, 31(1): 35–40. |
| [29] | Rakko A, Laugaste R, Ott I. Algal blooms in estonian small lakes//Evangelista V, Barsanti L, Frassanito A M, Passarelli V, Gualtieri P, eds. Algal Toxins: Nature, Occurrence, Effect and Detection. Netherlands: Springer, 2008: 211-220. |
| [30] | Sarmento H, Isumbisho M, Descy J P. Phytoplankton ecology of Lake Kivu (Eastern Africa). Journal of Plankton Research, 2006, 28(9): 815–829. DOI:10.1093/plankt/fbl017 |
| [31] | 李娣, 李旭文, 牛志春, 王霞, 师伟, 于红霞. 太湖浮游植物群落结构及其与水质指标间的关系. 生态环境学报, 2014, 23(11): 1814–1820. DOI:10.3969/j.issn.1674-5906.2014.11.014 |
| [32] | 胡韧, 雷腊梅, 韩博平. 南亚热带大型贫营养水库浮游植物群落结构与季节变化——以新丰江水库为例. 生态学报, 2008, 28(10): 4652–4664. DOI:10.3321/j.issn:1000-0933.2008.10.005 |