 2017, Vol. 37
2017, Vol. 37文章信息
- 王祖艳, 邵元虎, 夏汉平, 傅声雷
- WANG Zuyan, SHAO Yuanhu, XIA Hanping, FU Shenglei.
- 基于土壤线虫群落分析的油页岩废渣地不同植被类型恢复过程中的土壤食物网能流状况
- Energy flow analysis of soil food webs based on soil nematode communities:a case study from an oil shale mined land afforested with different pioneer tree species
- 生态学报. 2017, 37(17): 5612-5620
- Acta Ecologica Sinica. 2017, 37(17): 5612-5620
- http://dx.doi.org/10.5846/stxb201606021067
-
文章历史
- 收稿日期: 2016-06-02
- 网络出版日期: 2017-04-24





2. 中国科学院大学, 北京 100049;
3. 河南大学, 环境与规划学院, 黄河中下游数字地理技术实验室, 开封 475004
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Laboratory of Geospatial Technology for the Middle and Lower Yellow River Regions, College of Environment and Planning, Henan University, Kaifeng 475004, China
线虫作为土壤中数量最丰富的多细胞动物, 主要分为植食性线虫(主要以根系为食源)、食细菌线虫、食真菌线虫和捕食杂食类线虫4类营养类群[1]。土壤线虫是土壤生态系统的重要组成部分[2], 由于线虫具有对环境变化敏感、身体透明、周期性比较短等特点, 它们经常被作为评估环境健康的指示生物[3-4]。
利用线虫区系分析来推断食物网的结构和功能, 定性评价土壤食物网对外界干扰的响应是非常成熟的方法[5]。然而这种方法存在的一些不足, 在比较不同土壤类型的结果时会限制生态指数和方法的指示效果[6], 考虑到土壤线虫个体大小的巨大差异, 用不同类群的生物量去反映或评价土壤食物网的结构和功能应该比用线虫的数量或者密度等指标更加准确。根据食细菌、食真菌和植物寄生线虫的相对生物量做成三角图来分析食物网中的细菌、真菌和植物3种资源渠道, 被认为是一种有效的反映碳能量的来源及其通过食物网流动渠道的工具[7]。这可以更好的估计线虫对生态系统服务和功能方面的贡献, 因此基于不同营养类群的线虫生物量反映食物网能量流动情况越来越受到重视[8-9]。
随着社会经济的快速发展, 土壤污染问题日益突出, 油页岩废渣污染和污染地的修复已经成为严重的社会问题。油页岩废渣是利用油页岩提炼石油后的废弃物残渣, 废渣中含有大量的毒性或潜在毒性物质, 淋溶或扩散后严重污染周围的水源、土地及生物[10], 从而危害居民健康[11]。一般来讲, 栽种植物是污染废弃地恢复的常见手段之一。然而, 植物本身的特性差异往往会导致不同植物对污染地的恢复效果不同。为确定污染地的恢复情况, 可以用线虫作为土壤指示生物来评估不同植被类型的生态修复效果。植物根系及其分泌物会刺激植食性线虫、食细菌和食真菌线虫的增加[12], 这几类线虫进一步还会被更高营养级的线虫捕食;食细菌和食真菌线虫不仅可以通过取食微生物加速其周转, 并且可以通过代谢释放微生物所固持的养分进而促进植物生长[13]。土壤食物网中经过细菌、真菌和植食者的能量分别形成了细菌、真菌和植物3种能流通道[14-16], 线虫群落中的多个营养类群使得它们有条件反映土壤食物网中的能流通道。比如食细菌线虫和食真菌线虫的比值能反映出不同分解通道在分解过程中的相对重要性。当土壤有机质丰富且分解较快, 土壤食物网能量流动更倾向于细菌分解通道;而土壤有机质缺乏且难分解, 土壤食物网能量流动更倾向于真菌分解通道[17]。而用线虫的生物量来描述土壤食物网中的能流, 则可以更加准确的反映土壤食物网结构和养分动态[18]。
本研究拟对不同植被恢复下的废渣地基质进行采样, 提取其中的土壤线虫并通过分析线虫反映的土壤食物网能量流动状况, 进而探讨不同植被恢复条件下的土壤食物网结构及其稳定状况, 以期加深人们对人类活动越来越剧烈的背景下土壤食物网维持机制的理解。
1 研究方法 1.1 研究区域概况茂名市(21°25′—22°42′N, 110°21′—111°46′E)位于广东省西南部, 处于热带植被分布区的北缘, 气候为海洋性季风气候,整个区域的地带性植被为热带雨林。该地区年平均气温23.2℃,最热月均温和最冷月均温分别为28.4℃(7月)和15.5℃(1月),历年平均降雨量1567mm,雨热同期,无降雪或霜冻。研究样地为茂名石化北部的油页岩废渣排放场, 样地面积达6.7 km2。本实验中选取的乌墨、大叶相思、红荷3种纯林以及荒草地内的主要植被类型情况如下:
乌墨样地:树高一般约6—8 m, 胸径约10—15 cm, 郁闭度70%—80%, 林下物种盖度约10%—20%, 林下物种主要包括飞机草(Eupatorrium odoratum)、芒箕(Dicranopteris linearis)、乌毛蕨(Blechnum orientale)等。
大叶相思林样地:树高一般8—10 m, 郁闭度92%。林下层植被稀疏, 林下物种盖度约10%, 常见种类有日本草(Borreria latifolia)、大青(Clerodendron crytophyllum)、土密树(Bridelia tomentosa)等。
红荷林样地:林木层郁闭度80%—90%, 凋落物层厚3—5 cm, 林下物种盖度约10%, 林下层有零星分布的日本草(B. latifolia), 林缘植物主要有飞机草(E. odoratum)、胜红蓟(Ageratum conyzoides)等。
荒草地样地:草层覆盖度80%—90%, 以草本植物为主, 主要种类有:芒箕(Dicranopteris linearis)、白茅(Imperata cylindrical)、飞机草(E. odoratum)等。
1.2 样品采集2008年6月22日, 本实验在4种有植被样地以及无植被的废渣样地均随机选取3块重复样地, 每块重复样地面积约400—600 m2, 每块重复样地内采集4个混合土壤样品, 每个混合土壤样品由3个采样点混合而成, 采样深度为0—5 cm, 采样时几种人工林种植了约8a。采样时, 我们选取树冠垂直投影到地面的一个近似圆形区域的边界进行采样, 采样前把土壤表面的凋落物移除。采集好的土壤样品放入保温箱中保存, 样品带回实验室后转入4℃冰箱。样品用于土壤理化和生物分析前尽可能的去除石头、根系等杂质。
1.3 土壤线虫的提取、分离和鉴定土壤线虫用Baermann湿漏斗法提取[19], 其主要原理是利用线虫在土壤中活动能力较差, 但在浸水的情况下, 土壤中的线虫活动能力加强, 在重力作用下会慢慢向漏斗底部移动, 这样便可在漏斗下方收集线虫。实验时对每个土壤样品用50 g鲜土提取, 提取出来的线虫用热福尔马林溶液(4%甲醛)杀死固定, 然后在倒置显微镜下计数。每个样品随机取100条线虫在光学显微镜下鉴定至科属水平[20-21], 如果样品中不足100条线虫, 则所有样品都要鉴定。根据Yeates等线虫营养类群划分体系将线虫划分为4个营养类群:食细菌类、食真菌类、植物寄生类、捕食杂食类[1]。
1.4 数据统计与处理不同样地之间细菌、真菌和植物能流的百分比之间的差异用SPSS 15.0 (SPSS Inc., Chicago, IL)分析软件进行单因素方差分析, 显著性水平设置为P<0.05, Tukey用来检验不同样地间不同能流所占比重的差异。线虫生物量及重要值计算公式如下:
线虫生物量根据方程B=(D2×L)×(1.6×106)-1进行估算[22], 其中B指每条个体的生物量, 用μg鲜重表示;L是指线虫的体长, 用μm表示;D指身体最宽部分的直径, 用μm表示。某一属的线虫体长和体宽主要参考Goodey, Bongers以及Jairajpuri & Ahmad等线虫分类书籍中介绍的该属不同种的成虫的体长和体宽[20-21, 23], 具体来说是把该属的所有能查到的种的体长和体宽收集后取平均值。如果线虫只鉴定到科, 则按照上述方法把这一科内的所有线虫属估算出体长和体宽, 然后把这些线虫属的体长和体宽再取平均值。

|
土壤食物网能量流通状况分析参考Ferris和Bongers的方法[7]。首先在Excel表中拟定三角形的3个顶点的坐标, 左下角、右下角以及顶点坐标分别为(0, 0), (100, 0), (50, 86.6)。其次是根据几何知识确定每条边的中点及整个图的中心点的坐标。最后的5个样地的点的坐标则根据每个点食细菌、食真菌和植食性线虫的相对生物量确定。比如在乌墨林, 细菌相对生物量是81%, 在Y轴上的坐标点为 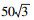
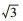
在整个样地来看, 乌墨林和大叶相思林线虫属的数量较多, 分别为54和45个, 线虫重要值最高的是食细菌的棱咽属(Prismatolaimus)和拟丽突属(Acrobeloides);而荷木林和荒草地的线虫属的数量较少, 分别为41和38个, 线虫的重要值最高的是杂食性的真矛线属(Eudorylaimus);废渣地线虫属的数量最少, 仅为34个, 重要值最高的是食真菌的丝尾垫刃属(Filenchus)和滑刃属(Aphelenchoides)(表 1)。
| 次序 Order |
乌墨S. cumini | 大叶相思A. auriculiformis | 红荷S.wallichii | 荒草地Grassland | 废渣Oil shale waste | |||||||||
| 属/科 Genera/Family |
重要值 Importance value |
属/科 Genera/Family |
重要值 Importance value |
属/科 Genera/Family |
重要值 Importance value |
属/科 Genera/Family |
重要值 Importance value |
属/科 Genera/Family |
重要值 Importance value |
|||||
| 1 | Prismatolaimus | 39.762 | Acrobeloides | 36.171 | Eudorylaimus | 23.762 | Eudorylaimus | 45.288 | Filenchus | 30.833 | ||||
| 2 | Protorhabditis | 21.043 | Eudorylaimus | 33.326 | Acrobeloides | 22.455 | Aphelenchoides | 30.621 | Aphelenchoides | 26.559 | ||||
| 3 | Tylenchidae | 16.985 | Protorhabditis | 26.071 | Mononchus | 19.887 | Acrobeloides | 25.881 | Acrobeloides | 24.775 | ||||
| 4 | Rhabditis | 14.133 | Rhabditis | 24.073 | Aphelenchoides | 18.783 | Microdorylaimus | 21.665 | Protorhabditis | 18.992 | ||||
| 5 | Acrobeloides | 12.529 | Prismatolaimus | 19.270 | Pratylenchus | 18.701 | Thonus | 18.038 | Tylenchidae | 18.601 | ||||
| 6 | Axonchium | 12.289 | Aphelenchoides | 16.922 | Thonus | 16.517 | Protorhabditis | 12.787 | Eudorylaimus | 17.301 | ||||
| 7 | Eudorylaimus | 12.013 | Tylenchidae | 11.996 | Iotonchus | 12.809 | Panagrolaimus | 12.058 | Thornia | 16.525 | ||||
| 8 | Pungentus | 11.828 | Eucephlobus | 11.854 | Tylenchidae | 11.624 | Filenchus | 11.919 | Prismatolaimus | 13.418 | ||||
| 9 | Panagrolaimus | 10.612 | Panagrolaimus | 11.666 | Filenchus | 10.692 | Tylenchidae | 11.483 | Rhabditis | 12.904 | ||||
| 10 | Prodorylaimus | 9.407 | Wilsonema | 10.729 | Belondira | 10.282 | Mononchus | 11.154 | Iotonchus | 9.130 | ||||
| 11 | Aphelenchoides | 9.160 | Filenchus | 8.502 | Dorylaimellus | 10.139 | Eucephalobus | 10.301 | Alaimus | 8.295 | ||||
| 12 | Diplogastrellus | 8.416 | Diplogastrellus | 6.913 | Prismatolaimus | 9.380 | Prismatolaimus | 9.802 | Wilsonema | 8.280 | ||||
| 13 | Eucephalobus | 8.301 | Cervidellus | 6.839 | Protorhabditis | 8.104 | Thornia | 9.167 | Eucephalobus | 7.795 | ||||
| 14 | Filenchus | 8.123 | Helicotylenchus | 6.809 | Panagrolaimus | 8.017 | Pratylenchus | 7.715 | Cervidellus | 7.537 | ||||
| 15 | Criconemella | 6.673 | Acrobeles | 5.598 | Microdorylaimus | 7.635 | Cervidellus | 5.306 | Criconemella | 7.490 | ||||
| 16 | Pratylenchus | 6.516 | Monhystera | 4.552 | Plectus | 6.041 | Rhabditis | 4.447 | Labronema | 6.156 | ||||
| 17 | Plectus | 6.347 | Dorylaimus | 4.501 | Thornia | 5.621 | Criconemella | 4.300 | Pratylenchus | 5.959 | ||||
| 18 | Thornia | 5.587 | Plectus | 3.088 | Cervidellus | 5.519 | Mesodorylaimus | 3.305 | Panagrolaimus | 5.596 | ||||
| 19 | Zygotylenchus | 5.279 | Longidorella | 3.061 | Eucephalobus | 5.340 | Malenchus | 3.177 | Microdorylaimus | 5.373 | ||||
| 20 | Hirschmanniella | 4.440 | Microdorylaimus | 2.862 | Rhabditis | 4.867 | Helicotylenchus | 2.895 | Helicotylenchus | 4.654 | ||||
| 21 | Scutellonema | 4.008 | Paratylenchus | 2.414 | Aporcelaimellus | 4.762 | Monhystera | 2.743 | Tylencholaimus | 3.977 | ||||
| 22 | Dorylaimus | 4.004 | Prodorylaimus | 1.960 | Diphtherophora | 4.342 | Wilsonema | 1.491 | Mononchus | 3.726 | ||||
| 23 | Aphelenchus | 3.274 | Lelenchus | 1.908 | Tylencholaimus | 3.834 | Longidorella | 1.321 | Axonchium | 3.341 | ||||
| 24 | Sectonema | 2.777 | Rhabditonema | 1.623 | Acrolobus | 3.832 | Heterocephalobus | 1.180 | Diplogastrellus | 3.077 | ||||
| 25 | Malenchus | 2.504 | Acrolobus | 1.271 | Helicotylenchus | 3.714 | Aporcelaimellus | 1.154 | Longidorella | 2.825 | ||||
| 26 | Lelenchus | 2.286 | Aphelenchus | 1.131 | Rhabditonema | 3.146 | Acrolobus | 1.029 | Acrolobus | 2.554 | ||||
| 27 | Helicotylenchus | 2.280 | Apocelaimellus | 0.837 | Wilsonema | 2.793 | Plectus | 0.952 | Aphelenchus | 2.539 | ||||
| 28 | Diplogasteroides | 2.068 | Pratylenchus | 0.778 | Teratorhabditis | 2.680 | Prodorylaimus | 0.942 | Aporcelaimellus | 1.336 | ||||
| 29 | Cervidellus | 1.763 | Pelodera | 0.676 | Criconemella | 2.368 | Cephalobus | 0.705 | Mesodorylaimus | 1.301 | ||||
| 30 | Alaimus | 1.493 | Alaimus | 0.654 | Cephalobus | 2.258 | Lelenchus | 0.647 | Trichodorus | 1.118 | ||||
| 31 | Xiphinema | 1.449 | Mesorhabditis | 0.640 | Cruznema | 1.284 | Tylencholaimus | 0.571 | Rhabditonema | 0.972 | ||||
| 32 | Pelodera | 1.306 | Rotylenchus | 0.631 | Diplogastrellus | 1.275 | Acrobeles | 0.432 | Acrobeles | 0.922 | ||||
| 33 | Iotonchus | 1.083 | Heterocephalobus | 0.611 | Diplogasteroides | 0.817 | Diphtherophora | 0.391 | Malenchus | 0.537 | ||||
| 34 | Tylenchorhynchus | 1.043 | Diplogasteritus | 0.589 | Acrobeles | 0.812 | Rhabditonema | 0.344 | Paratylenchus | 0.490 | ||||
| 35 | Heterocephalobus | 0.949 | Diplogasteriana | 0.507 | Alaimus | 0.782 | Diplogastrellus | 0.302 | - | - | ||||
| 36 | Euteratocephalus | 0.939 | Cephalobus | 0.479 | Diplogasteritus | 0.655 | Alaimus | 0.295 | - | - | ||||
| 37 | Monhystera | 0.925 | Discomyctus | 0.332 | Aphelenchus | 0.639 | Diplogasteritus | 0.265 | - | - | ||||
| 38 | Diplogasteritus | 0.630 | Thonus | 0.304 | Kochinema | 0.624 | Dorylaimellus | 0.170 | - | - | ||||
| 39 | Diphtherophora | 0.591 | Thornia | 0.225 | Teratocephalus | 0.525 | - | - | - | - | ||||
| 40 | Dorylaimoides | 0.585 | Nothotylenchus | 0.196 | Diplogaster | 0.327 | - | - | - | - | ||||
| 41 | Diplogaster | 0.466 | Drilocephalobus | 0.145 | Malenchus | 0.289 | - | - | - | - | ||||
| 42 | Nygolaimus | 0.428 | - | - | Prodorylaimus | 0.245 | - | - | - | - | ||||
| 43 | Mesorhabditis | 0.395 | - | - | Epidorylaimus | 0.233 | - | - | - | - | ||||
| 44 | Psilenchus | 0.362 | - | - | Paratylenchus | 0.195 | - | - | - | - | ||||
| 45 | Rhabditoides | 0.342 | - | - | Lelenchus | 0.137 | - | - | - | - | ||||
| 46 | Rotylenchus | 0.305 | - | - | - | - | - | - | - | - | ||||
| 47 | Rhabditonema | 0.283 | - | - | - | - | - | - | - | - | ||||
| 48 | Anaplectus | 0.267 | - | - | - | - | - | - | - | - | ||||
| 49 | Microdorylaimus | 0.216 | - | - | - | - | - | - | - | - | ||||
| 50 | Tripyla | 0.208 | - | - | - | - | - | - | - | - | ||||
| 51 | Amphidirhabditis | 0.193 | - | - | - | - | - | - | - | - | ||||
| 52 | Panagrobelus | 0.183 | - | - | - | - | - | - | - | - | ||||
| 53 | Alirhabditis | 0.164 | - | - | - | - | - | - | - | - | ||||
| 54 | Wilsonema | 0.143 | - | - | - | - | - | - | - | - | ||||
细菌能流通道的比重最高的是大叶相思林和乌墨林, 而真菌能流通道比重最高的是废渣地, 红荷林和荒草地居中, 各样地植物能流通道的比重都比较小(图 1)。进一步根据食细菌、食真菌和植物寄生线虫的相对生物量进行土壤食物网能流分析后发现各样地细菌通道所占的比重范围在56%—88%之间, 真菌通道的能流通道所占的比重范围在10%—39%之间, 植物能流通道所占的比重范围在2%—10%之间, 各样地植物能流通道的比重没有显著的差异(图 1B)。

|
| 图 1 不同样地的能流分析 Fig. 1 Energy flow analysis for different sites A:根据食细菌、食真菌和植物寄生性线虫的相对生物量反映的土壤食物网中的细菌、真菌和植物三种能流通道;B:不同样地下3种能流的百分比,B中显示的是平均值±标准误; 图中的字母不同表示各样地之间有显著性差异 |
线虫的重要值可以更加客观的反映样地不同线虫的重要性, 根据线虫的不同, 我们也就可以更加准确地了解线虫反映的土壤食物网状况。本研究中, 在乌墨林和大叶相思林, 线虫重要值最高的是食细菌的棱咽属(Prismatolaimus)和拟丽突属(Acrobeloides), 而食细菌线虫取食细菌之后往往能促进土壤的氮矿化[25], 进而可以影响植物的生长。表明乌墨和大叶相思恢复过程中这两种植物通过根系分泌物的输入刺激了细菌能流通道, 反过来细菌能流通道快速的养分周转也可能会刺激植物生长。而细菌能流通道的比重较高时, 食物网中能量流动速度较快, 食物网受到干扰后比较容易恢复。在红荷林和荒草地, 重要值最高的是杂食性的真矛线属(Eudorylaimus), 这类线虫代表了食物网中较高的营养级, 表明食物网内的关系比较复杂。在废渣样地, 重要值最高的是食真菌的丝尾垫刃属(Filenchus)和滑刃属(Aphelenchoides), 有研究表明养分条件比较差的时候, 食物网中真菌的代谢作用比细菌更大[26], 因为真菌生物量要比细菌生物量周转更慢, 并且土壤动物消耗的真菌生物量的比例低于细菌生物量的比例, 所以真菌通道占优势的食物网在保持土壤养分方面更有效[14, 27-28]。废渣样地因为没有植物资源输入, 土壤受干扰强烈, 土壤养分状况更差, 这个时候更高比重的真菌能流通道更加有利于保持土壤养分。另外, 真菌为主的能流通道一般是相对慢速的能流通道, 大多数食物网的分析[29-32]表明相对慢速的能流通道的食物网内的关系都比较弱[33], 这意味着土壤食物网受到外界影响时, 这些影响会通过改变食物网能量组织形式(如细菌通道和真菌通道相对大小)来改变土壤食物网稳定性[34], 其中各种生物的关系可能会发生变化, 这种变化可能会导致土壤食物网的结构发生转变, 进而会对土壤生态系统的稳定性造成影响。比如本研究中的真菌通道权重最大的废渣地, 受到外界影响后有最大的恢复潜力, 但也是最难恢复的, 较慢速的真菌能流通道在土壤食物网中的相互关系较弱, 而这种弱的关系对于维持油页岩废渣地的生态系统土壤食物网稳定性至关重要, 这与Rooney、Andrés等人的观点相一致[30, 32-33]。整个实验样地, 植食性线虫的重要值都不是特别高, 一方面是因为那些个体较小的植食性线虫, 比如垫刃科(Tylenchidae)中的一些植食性线虫是兼性植食的, 它们可以取食植物的根毛也可以取食根系分泌物或少量真菌[12]。那些个体较大的植物寄生线虫经常会把口器刺入到植物根系的深层细胞进行取食[35]。而对大多数的植食者来说养分是限制因子,尤其是氮[36], 油页岩废渣这种特殊的基质中含有的有害金属元素会对土壤及土壤生物造成一定的影响, 使土壤养分比较贫瘠、有效性也比较差, 从养分和能量需求的角度看, 这些大个体的植食性线虫可能更加容易受到养分贫瘠的影响。另一方面, 很多植物寄生性线虫喜欢高pH的土壤[37], 根据我们早期的研究结果发现, 整个样地的pH在4.0—4.4之间[38], 较低的pH值可能也对植食性线虫产生了影响。
几种食物网能流分析表明:在整个油页岩废渣场各个样地, 植物能流通道所占的比重都比较小, 这与植食性线虫的重要值反映的规律基本一致。细菌和真菌能流通道所代表的营养系统是腐屑为基础的营养系统, 与植物为基础的营养系统相比, 这种营养系统能将更高比例的可利用能量转化为生产量[39], 在我们的研究中, 细菌和真菌能流通道的权重更高, 这可能有利于这种特殊基质的恢复。
不同样地在土壤线虫重要值以及土壤食物网能流方面的差异可能是受种植8年的不同植物的影响, 不同植被类型下的土壤食物网健康状况因植被的不同而产生了植被根际生态学、土壤养分特征、pH或者污染等方面的差异[40], 并且影响到了土壤线虫的群落组成以及多度。本研究中发现乌墨林线虫的科属数量是最多的, 而我们早期的研究发现不同植被类型下, 土壤线虫密度差别很大。具体来说, 大叶相思林线虫密度最高, 主要是大叶相思林有更高密度的低营养级水平的食细菌和食真菌线虫, 红荷林的植食性线虫以及高营养级的捕食杂食性线虫密度更高, 乌墨林食真菌线虫密度最低, 但其食物网健康状况更好[38]。说明植物根系碳的输入不仅可以刺激植食性线虫的增加, 也可能通过根系分泌物的输入刺激食细菌和食真菌线虫的增加[12], 从而进一步影响更高营养级的捕食和杂食性线虫。从这个角度看, 植物资源的输入对细菌、真菌以及植物能流通道均产生了较大的影响, 尽管植物能流通道在各样地所占的比重比较小, 但植被仍是土壤线虫群落发展的主要驱动力。另外, 不同植被类型下的土壤微生物群落、土壤理化以及土壤污染物属性也表现出不同的特征。比如, 乌墨林下有更高的菌根真菌含量和较低的总的铝含量;尽管废渣地有较高的总铝含量, 但活性铝含量不是特别高; 大叶相思作为固氮树种, 土壤总氮含量更高[38], 这些都说明了植物树种本身的属性与油页岩废渣地的恢复关系密切。比如在油页岩这种养分贫瘠并且铝毒污染比较严重的基质进行植被恢复, 菌根植物和固氮植物可能是比较适合的选择, 而土壤线虫反映不同植被恢复条件下的土壤食物网健康和养分循环情况, 可能有利于油页岩废渣地的恢复过程中植物物种的筛选。
| [1] | Yeates G W, Bongers T, De Goede R G, Freckman D W, Georgieva S S. Feeding habits in soil nematode families and genera-an outline for soil ecologists. Journal of Nematology, 1993, 25(3): 315–331. |
| [2] | Freckman D W. Bacterivorous nematodes and organic-matter decomposition. Agriculture, Ecosystems and Environment, 1988, 24(1/3): 195–217. |
| [3] | 梁文举, 闻大中. 土壤生物及其对土壤生态学发展的影响. 应用生态学报, 2001, 12(1): 137–140. |
| [4] | Neher D A, Wu J, Barbercheck M E, Anas O. Ecosystem type affects interpretation of soil nematode community measures. Applied Soil Ecology, 2005, 30(1): 47–64. DOI:10.1016/j.apsoil.2005.01.002 |
| [5] | Ferris H, Bongers T, De Goede R G M. A framework for soil food web diagnostics:extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13–29. DOI:10.1016/S0929-1393(01)00152-4 |
| [6] | Yeates G W. Nematodes as soil indicators:functional and biodiversity aspects. Biology and Fertility of Soils, 2003, 37(4): 199–210. |
| [7] | Ferris H, Bongers T. Nematode indicators of organic enrichment. Journal of Nematology, 2006, 38(1): 3–12. |
| [8] | Scharroba A, Kramer S, Kandeler E, Ruess L. Spatial and temporal variation of resource allocation in an arable soil drives community structure and biomass of nematodes and their role in the micro-food web. Pedobiologia, 2016, 59(3): 111–120. DOI:10.1016/j.pedobi.2016.03.005 |
| [9] | Zhang X K, Guan P T, Wang Y L, Li Q, Zhang S X, Zhang Z Y, Bezemer T M, Liang W J. Community composition, diversity and metabolic footprints of soil nematodes in differently-aged temperate forests. Soil Biology and Biochemistry, 2015, 80: 118–126. DOI:10.1016/j.soilbio.2014.10.003 |
| [10] | 夏汉平, 黄娟, 孔国辉. 油页岩废渣场的生态恢复. 生态学报, 2004, 24(12): 2887–2893. DOI:10.3321/j.issn:1000-0933.2004.12.033 |
| [11] | 肖彦春. 油页岩及废渣农业利用研究进展. 辽宁农业职业技术学院学报, 2014, 16(4): 12–14. |
| [12] | Yeates G W. Effects of plants on nematode community structure. Annual Review of Phytopathology, 1999, 37(37): 127–149. |
| [13] | 吴纪华, 宋慈玉, 陈家宽. 食微线虫对植物生长及土壤养分循环的影响. 生物多样性, 2007, 15(2): 124–133. |
| [14] | Moore J C, Hunt H W. Resource compartmentation and the stability of real ecosystems. Nature, 1988, 333(6170): 261–263. DOI:10.1038/333261a0 |
| [15] | De Vries F T, Thébault E, Liiri M, Birkhofer K, Tsiafouli M A, Bjørnlund L, Bracht Jørgensen H, Brady M V, Christensen S, De Ruiter P C, d'Hertefeldt T, Frouz J, Hedlund K, Hemerik L, Hol W H G, Hotes S, Mortimer S R, Setälä H, Sgardelis S P, Uteseny K, Van Der Putten W H, Wolters V, Bardgett R D. Soil food web properties explain ecosystem services across European land use systems. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(35): 14296–14301. DOI:10.1073/pnas.1305198110 |
| [16] | Neutel A M, Heesterbeek J A, De Ruiter P C. Stability in real food webs:weak links in long loops. Science, 2002, 296(5570): 1120–1123. DOI:10.1126/science.1068326 |
| [17] | Ingwersen J, Poll C, Streck T, Kandeler E. Micro-scale modelling of carbon turnover driven by microbial succession at a biogeochemical interface. Soil Biology and Biochemistry, 2008, 40(4): 864–878. DOI:10.1016/j.soilbio.2007.10.018 |
| [18] | Moore J C, De Ruiter P C. Temporal and spatial heterogeneity of trophic interactions within below-ground food webs. Agriculture Ecosystems and Environment, 1991, 34(1/4): 371–397. |
| [19] | Barker K R, Carter C C, Sasser J N. An Advanced Treatise on Meloidogyne. Volume Ⅱ:Methodology. Raleigh:North Carolina University Graphics, 1985. |
| [20] | Bongers T. De Nematoden van Nederland. Utrecht, Netherlands:Stichting Uitgeverij Koninklijke Nederlandse Natuurhistorische Vereniging, 1988. |
| [21] | Loof P A A, Jairajpuri M S, Ahmad W. Dorylaimida:Free-Living, Predaceous and Plant-Parasitic Nematodes. Fundamental and Appilied Nematology, 1992, 3: 286–287. |
| [22] | Andrassy I. Die rauminhalts-und gewichtsbestimmung der fadenwürmer (Nematoden). Acta Zoologica Academiae Scientiarum Hungaricae, 1956, 2: 1–15. |
| [23] | Goodey T, Goodey J B. Soil and Freshwater Nematodes. New York: John Wiley and Sons, 1963: 219–220. |
| [24] | Norton D C, Schmitt D P. Community analyses of plant-parasitic nematodes in the Kalsow Prairie, Iowa. Journal of Nematology, 1978, 10(2): 171–176. |
| [25] | Anderson R V, Coleman D C, Cole C V. Effects of saprotrophic grazing on net mineralization//Clark F E, Rosswall T, eds. Terrestrial Nitrogen Cycles:Processes, Ecosystem Strategies, and Management Impacts. Stockholm:Swedish Natural Science Research Council, 1981:201-216. |
| [26] | Williamson W M, Wardle D A, Yeates G W. Changes in soil microbial and nematode communities during ecosystem decline across a long-term chronosequence. Soil Biology and Biochemistry, 2005, 37(7): 1289–1301. DOI:10.1016/j.soilbio.2004.11.025 |
| [27] | Coleman D C, Reid C P P, Cole C V. Biological strategies of nutrient cycling in soil systems. Advances in Ecological Research, 1983, 13(4): 1–55. |
| [28] | Wardle D A, Yeates G W. The dual importance of competition and predation as regulatory forces in terrestrial ecosystems:evidence from decomposer food-webs. Oecologia, 1993, 93(2): 303–306. DOI:10.1007/BF00317685 |
| [29] | Moore J C, De Ruiter P C, Hunt H W, Coleman D C, Freckman D W. Microcosms and soil Ecology:Critical linkages between field studies and modelling food webs. Ecology, 1996, 77(3): 694–705. DOI:10.2307/2265494 |
| [30] | Rooney N, Mccann K S. Integrating food web diversity, structure and stability. Trends in Ecology and Evolution, 2012, 27(1): 40–46. DOI:10.1016/j.tree.2011.09.001 |
| [31] | Zhao J, Neher D A. Soil energy pathways of different ecosystems using nematode trophic group analysis:A meta analysis. Nematology, 2014, 16(4): 379–385. DOI:10.1163/15685411-00002771 |
| [32] | Andrés P, Moore J C, Simpson R T, Selby G, Cotrufo F, Denef K, Haddix M L, Shaw E A, De Tomasel C M, Molowny-Horas R, Wall D H. Soil food web stability in response to grazing in a semi-arid prairie:The importance of soil textural heterogeneity. Soil Biology and Biochemistry, 2016, 97: 131–143. DOI:10.1016/j.soilbio.2016.02.014 |
| [33] | Rooney N, Mccann K, Gellner G, Moore J C. Structural asymmetry and the stability of diverse food webs. Nature, 2006, 442(7100): 265–269. DOI:10.1038/nature04887 |
| [34] | 陈云峰, 唐政, 李慧, 韩雪梅, 李钰飞, 胡诚. 基于土壤食物网的生态系统复杂性-稳定性关系研究进展. 生态学报, 2014, 34(9): 2173–2186. |
| [35] | Bongers T, Bongers M. Functional diversity of nematodes. Applied Soil Ecology, 1998, 10(3): 239–251. DOI:10.1016/S0929-1393(98)00123-1 |
| [36] | Mattson Jr W J. Herbivory in relation to plant nitrogen content. Annual Review of Ecology and Systematics, 1980, 11(1): 119–161. DOI:10.1146/annurev.es.11.110180.001003 |
| [37] | Brzeski M W, Dowe A. Effect of pH on Tylenchorhynchus dubius (Nematoda, Tylenchidae). Nematologica, 1968, 15(3): 403–407. |
| [38] | Shao Y H, Zhang W X, Liu Z F, Sun Y X, Chen D M, Wu J P, Zhou L X, Xia H P, Neher D A, Fu S L. Responses of soil microbial and nematode communities to aluminum toxicity in vegetated oil-shale-waste lands. Ecotoxicology, 2012, 21(8): 2132–2142. DOI:10.1007/s10646-012-0966-4 |
| [39] | Chapin F S I, Matson P A I, Mooney H A. Principles of Terrestrial Ecosystem Ecology. New York: Springer Berlin, 2002. |
| [40] | Bongers T, Ferris H. Nematode community structure as a bioindicator in environmental monitoring. Trends in Ecology and Evolution, 1999, 14(6): 224–228. DOI:10.1016/S0169-5347(98)01583-3 |