 2017, Vol. 37
2017, Vol. 37文章信息
- 刘长发, 苑静涵, 刘远, 方蕾, 王艺婷, 陶韦, 李晋
- LIU Changfa, YUAN Jinghan, LIU Yuan, FANG Lei, WANG Yiting, TAO Wei, LI Jin.
- 双台河口翅碱蓬生长与根、茎、叶碳、氮的分配
- Analysis of growth and allocation of carbon and nitrogen in different organs (root, stem, and leaf) of Suaeda heteroptera, a halophyte thriving in Shuangtai Estuary
- 生态学报. 2017, 37(16): 5492-5503
- Acta Ecologica Sinica. 2017, 37(16): 5492-5503
- http://dx.doi.org/10.5846/stxb201605110918
-
文章历史
- 收稿日期: 2016-05-11
- 网络出版日期: 2017-03-25






2. 盘锦市海洋与渔业科学研究所, 盘锦 124010
2. Panjin Institute of Ocean and Fisheries, Panjin 124010, China
生物个体生长过程是其同化作用和异化作用过程中物质和能量吸收、转运、储存、排出平衡的综合结果。因此, 供生物生长的物质和能量在其被生物吸收、转运、储存、排出的生态生理学过程中, 各种物质和能量的供给可能成为生物生长的限制要素。植物生长限制要素可能是空间、光照、水、养分以及可能的环境要素胁迫等。在长期的适应生存和自然选择下, 植物在生态与进化环境中形成了不同的表现, 即生存对策——C-S-R对策(竞争型C;耐胁迫型S;杂草型R)[1]。生长和物质在不同器官间的分配是植物重要的生存对策。植物遗传背景和环境要素相互作用可导致其呈现不同的生长速率。一般地, 生物的生长速率随着生长时间的延长呈现下降趋势, 生长过程则呈“S”形曲线型式[2], von Bertalanffy方程、Gompertz方程和Logistic方程等3参数生长方程, 以及Weibull方程、Monomolecular方程、Richards方程、General von Bertalanffy方程、Schnute-Richards方程、Hossfeld IV方程、Chapman-Richards方程、Levakovic I方程、Levakovic III方程、Korf方程、Yoshida I方程、Sloboda方程等更多参数的方程被用于描述生物生长过程[2-7]。其中, Logistic方程和Gompertz方程分别是Richards方程的特殊形式。3参数方程因简单多用于描述生物个体乃至种群生长。由于多个数学模型均可以描述生物生长过程, 如何选择合适的模型近年来受到重视。在模型选择时, 模型的决定系数(R2)、校正决定系数(adj-R2)、参数估计偏度Hougaard绝对值(h)、赤池信息量准则(AIC)、贝叶斯信息量准则(BIC)、偏离修正AIC(AICC)、均方根误差(RMSE)、尺度准则值Δi分别被用于选择模拟模型[5-6, 8]。其中, 用于小样本量, 基于Kullback-Leibler距离、偏离修正AIC(AICC)和校正决定系数(adj-R2)因考虑了模型参数个数而多用于生物生长模型的选择依据[6, 8-9]。虽然按照AICC和adj-R2可以获得“最佳”模型, 但每种模型的参数推断均有条件约束, 存在一定的不确定性[6, 9]。多模型推断(MMI)提供了一种“无条件的”模型参数选择方法[6, 9]。Katsanevakis[6]建议采用基于赤池权重法计算模型参数平均值, 从而获得模型参数的稳健估计。
植物可将吸收的碳、氮等物质储存在根、茎、叶或特化的储存器官中, 但储存物质的种类、数量和储存的器官依植物种类不同而不同[10]。植物储存分为3类:积累——过渡性储存、储存物形成和再循环——再利用[10]。碳、氮是植物储存并构成其有机体的主要生源要素, 植物体内一般含有相当于其干重38%—46%的碳和1%—7%的氮, 并依植物种类、习性和龄期产生较大变化[11]。植物主要通过叶片光合作用吸收碳、通过根吸收氮, 进入植物体内的碳、氮在不同器官间分配, 则需要碳、氮长距离转运, 以供再利用, 如生长。植物同化物的分配过程是植物生长的关键, 可决定植物的光合作用强弱、生长速率、生长模式、竞争能力、植物结构的形成以及化学防御强度等[12-13]。生物质分配型式影响水生和陆地植被从个体生长与生殖到群落所有水平的物质交换和能量流动[14]。植物同化物的分配储存量与生长间常常表现为相关生长关系[15-19], 采用幂函数方程Y=aXb描述。a为归一化常数, 是温度依赖的, 且生物类群内和生物类群间不相同;b为相关生长指数[20]。b > 1为正异速生长, b=1为等速生长, b<1为负异速生长[15]。相关生长指数b=3/4或2/3, 被认为是“理想值”, 但受温度、生理状态、个体大小和环境资源的可用性等多种因素影响产生变化[21-22]。
翅碱蓬(Suaeda heteroptera)是生长在滨海潮滩湿地的耐盐先锋植物, 可作为植物生物修复物种用于沉积物污染植物修复。生长在滨海潮滩湿地的翅碱蓬植株呈紫红色, 可成为重要的生态工程岸线和旅游景观。种植碱蓬能显著增加各层土壤的总孔隙度、减少各层土壤容重及提高土壤的含水量, 降低各层土壤盐分[23], 改善土壤理化性质和土壤微生物的数量[24-25], 修复重金属[26]、石油烃污染[27]。李征等[28]研究了滨海盐地碱蓬不同生长阶段叶片碳、氮、磷化学计量特征, Mou等[29]研究了黄河口典型盐地碱蓬湿地生态系统的氮循环, 但缺乏对翅碱蓬生长与根、茎、叶碳、氮分配的研究。
植物生长与碳、氮分配、积累不仅存在个体差异, 而且受环境条件影响。植物为适应环境胁迫可表现出不同的生理生态特征。植物株高是植物的基本形态参数, 植物株高不同, 水分和养分转运距离不同, 获得的光照亦可能不同。并且, 植物在生长和碳、氮储存间的分配也是植物应对环境胁迫的策略之一。因此, 研究翅碱蓬株高生长以及根、茎、叶器官的碳、氮分配, 有助于进一步认识翅碱蓬在双台河口的种群乃至群落动态, 为分析翅碱蓬种群退化和区域物种生态演替提供参考。为此, 我们采集了双台河口滨海潮滩湿地生长的翅碱蓬, 测定了株高和根、茎、叶碳、氮含量, 模拟了翅碱蓬生长, 研究了株高生长与根、茎、叶碳、氮分配间的相关生长关系, 以回答如下问题:(1) 采用什么样的模型模拟双台河口翅碱蓬株高生长;(2) 双台河口潮滩湿地生长的翅碱蓬的渐近株高是多少?(3) 翅碱蓬根、茎、叶碳、氮含量与株高的相关生长关系。
1 材料与方法 1.1 材料翅碱蓬样品于2013年5月15日(0 d)、7月3日(49 d)、8月14日(91 d)、9月23日(134 d)、11月10日(179 d)采集自辽宁省“双台河口国家级自然保护区”滨海潮滩湿地(40°54′—40°58′N, 121°45′—121°50′E)。每次采样设置3个采样区域(40°55′24″N, 121°46′53″E;40°55′34″N, 121°46′47″E;40°55′49″N, 121°46′45″E), 在每个采样区域布设15个1m×1 m样方, 并随机编号。采集样方内所有翅碱蓬样品, 装入自封袋带回实验室。
1.2 测定方法将在每个样方中采集的翅碱蓬样品随机抽取30株用直尺量取株高, 并用纯水洗净后分离根、茎、叶。分离的根、茎、叶于70℃烘干后采用球磨仪(天津东方天净科技有限公司)磨碎, 置于棕色玻璃瓶中待测。测定前混合同一编号的3个采样区域的样品于70℃烘干至衡重, 采用微量电子天平(XP6型, 瑞士Mettler Toledo公司)称重、元素分析仪(vario PYRO cube型, 德国Elementar公司)测定翅碱蓬根、茎、叶碳、氮含量。
1.3 生长方程(模拟模型)3参数生长方程von Bertalanffy方程、Logistic方程、Gompertz方程[2]用于拟合翅碱蓬株高生长。von Bertalanffy生长方程
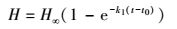
|
(1) |
式中, H表示翅碱蓬株高(cm);H∞表示翅碱蓬渐近株高(cm);k1表示相对生长速率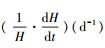
Logistic生长方程
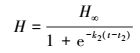
|
(2) |
式中, H表示翅碱蓬株高(cm);H∞表示翅碱蓬渐近株高(cm);k2表示相对生长速率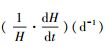
Gompertz生长方程
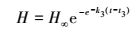
|
(3) |
式中, H表示翅碱蓬株高(cm);H∞表示翅碱蓬渐近株高(cm);k3表示相对生长速率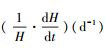
因考虑了参数个数, 用于小样本量, 基于Kullback-Leibler信息(K-L距离)、偏离修正的赤池信息量准则AICC[9]和校正决定系数(adj-R2)用于生长方程选择依据。AICC计算式
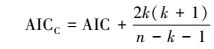
|
(4) |
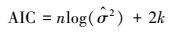
|
(5) |
式中, RSS表示误差平方和, k表示方程参数个数, n表示样本量。
具有最小AICC值(AICC, min)的模拟模型将被选择为“最佳”模型,即优选模型。
尽管按照AICC, min和adj-R2可以选择出“最佳”模型, 但是, 对于各模拟模型得到的渐近株高而言, 采用各模型推断渐进株高仍有条件约束, 存在一定的不确定性。多模型推断(MMI)可被用于计算平均期望渐近株高[9]。方法是分别计算各备选模型的期望渐近株高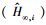
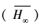
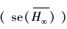
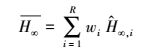
|
(7) |
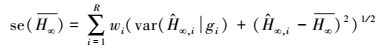
|
(8) |
式中, 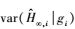
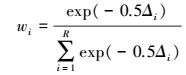
|
(9) |
式中, Δi表示备选模型差值, Δi=AICC, i-AICC, min;R表示备选模型个数。
1.5 拟合率模型拟合率(%)按式(10) 计算[30]

|
(10) |
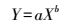
|
(11) |
式中, a为归一化常数, b为相关生长指数。
1.7 碳、氮相对含量植物生源要素相对含量是反映在一定环境条件下植物种群储存生源要素状况相对衡量的综合指标, 作为植物生态关系及其环境适应的间接评价方法, 可用于比较植物种群间的生理生态差异。参考对鱼类相对重量的研究[32], 按式(12) 计算翅碱蓬不同营养器官碳、氮相对含量(Cr-R)
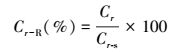
|
(12) |
式中, Cr表示实际测定的翅碱蓬不同营养器官的碳、氮含量, Cr-s表示特定株高翅碱蓬不同营养器官的“标准”碳、氮含量, 简单地, 取在同一采样时间采集的翅碱蓬的测量株高按含量-株高关系计算的平均碳、氮含量的3/4百分位的值。
1.8 相对条件因子相对条件因子同样是反映在一定环境条件下植物种群储存生源要素状况相对衡量的综合指标, 但其主要缺点是需要对每个区域中的生物种群分别依据含量-株高关系计算碳、氮的估计含量。参考对鱼类相对条件因子的研究[33], 按式(13) 计算翅碱蓬不同营养器官碳、氮含量的相对条件因子(Kr)
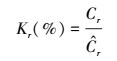
|
(13) |
式中, 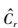
本文采用Χ2-检验方法检验分析了翅碱蓬株高分布;采用非线性回归方法模拟了株高生长, 并对模型参数渐近株高(H∞)进行了点估计和区间估计;采用线性回归方法模拟了相关生长关系, 并分别采用F-检验和t-检验对模型和模型参数b进行了显著性检验;采用离散型数据变异程度分析方法分析了株高生长分异;采用独立样本t-检验方法分析检验了碳、氮含量差异显著性。
2 结果 2.1 生长模拟采用3参数生长方程von Bertalanffy方程、Logistic方程、Gompertz方程拟合的双台河口翅碱蓬株高生长的结果见图 1。由von Bertalanffy方程拟合的翅碱蓬渐近株高H∞为38.95 cm, 相对生长速率k1为0.019 d-1, 理论生长起始时间t0为采样前11.6 d, 即4月30日。由Logistic方程拟合的翅碱蓬渐近株高H∞为37.42 cm, 相对生长速率k2为0.043 d-1, Logistic生长曲线拐点时间t2为32.12 d, 即6月16日。由Gompertz方程拟合的翅碱蓬渐近株高H∞为37.96 cm, 相对生长速率k3为0.030 d-1, Gompertz生长曲线拐点时间t3为16.38 d, 即6月1日。

|
| 图 1 翅碱蓬株高生长模拟曲线 Fig. 1 Fitted growth curves in height of halophyte Suaeda heteroptera |
图 2为按照R=(ln Hi+1-ln Hi)/(ti+1-ti)计算的各采样时间间隔内翅碱蓬株高平均相对生长率随时间变化。由图 2, 翅碱蓬株高生长的相对生长率随时间减小。

|
| 图 2 翅碱蓬株高相对生长率变化 Fig. 2 Changes of relative growth rate in height of halophyte Suaeda heteroptera |
按von Bertalanffy方程计算的翅碱蓬生长性能指数(φ′=2logH∞+logk1)φ′=2log(38.95) + log(0.019)=1.459。按Logistic方程计算的翅碱蓬生长始盛期为5月17日前, 高峰期为5月17日至7月17日, 盛末期为7月17日后。
2.2 生长模型选择模拟翅碱蓬株高生长的3参数方程选择参数和渐近株高见表 1。由表 1, Logistic方程的adj-R2最大、AICC最小, 且拟合率最大, 可以认为Logistic方程是模拟双台河口潮滩湿地翅碱蓬株高生长的优选方程。且Logistic方程的拟合率最高, 为95.5%。
| 模型 Candidate model |
adj-R2 | AICC | Δi | wi | 拟合率 Fitting percentage/% |
理论渐近株高Asymptotic height H∞/cm | |||
| 点估计 Point estimate |
标准差 SD |
95%置信区间 95% CL | |||||||
| von Bertalanffy | 0.872 | 35.04 | 2.16 | 0.180 | 94.3±4.1 | 38.95 | 3.76 | 22.78 | 55.12 |
| Logistic | 0.953 | 32.87 | 0 | 0.534 | 95.5±2.7 | 37.42 | 1.39 | 31.43 | 43.41 |
| Gompertz | 0.916 | 34.12 | 1.25 | 0.286 | 94.6±3.0 | 37.96 | 2.20 | 28.49 | 47.43 |
| adj-R2:校正决定系数;AICC:偏离修正赤池信息量准则;Δi:备选模型差值;wi:赤池权重 | |||||||||
按照多模型推断(MMI)方法计算得到的双台河口潮滩湿地生长的翅碱蓬的平均渐近株高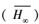
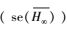
翅碱蓬株高生长分异程度见表 2、图 3。Χ2-检验结果表明翅碱蓬株高符合对数正态分布(0 d:P=0.55;49 d:P=0.30;91 d:P=0.10;134 d:P=0.84;179 d:P=0.77)(图 3), 但离散系数、偏度和峰度均较大, 其中初次和末次采集的翅碱蓬株高离散系数更大, 其离散系数为17.1%—30.8%。可见翅碱蓬株高生长产生分异, 表明双台河口潮滩湿地翅碱蓬的株高生长不同步, 可能受环境条件影响较大。
| 采样时间/d Sampling time |
平均值 Mean |
中位数 Median |
范围 Range |
标准差 SD |
离散系数 CV |
偏度 Skewness |
峰度 Kurtosis |
| 0 | 8.10 | 7.75 | 14.86 | 2.50 | 0.31 | 0.70 | 0.61 |
| 49 | 24.17 | 24.09 | 31.96 | 5.25 | 0.22 | 0.14 | 0.08 |
| 91 | 36.91 | 36.84 | 39.43 | 6.94 | 0.19 | 0.14 | -0.37 |
| 134 | 36.29 | 35.89 | 39.44 | 6.22 | 0.17 | 0.28 | 0.23 |
| 179 | 36.62 | 35.60 | 47.94 | 8.67 | 0.24 | 0.66 | 0.61 |

|
| 图 3 不同生长时间翅碱蓬株高对数正态分布拟合 Fig. 3 The fitting of log-normal distribution in height of halophyte Suaeda heteroptera at the observation day |
翅碱蓬根、茎、叶碳、氮含量见表 3、表 4。由表 3、表 4, 可见翅碱蓬根碳、氮含量在不同生长时期有相似的差异显著性关系, 即较小个体与较大个体根碳、氮含量间存在极显著差异。早期快速生长阶段, 翅碱蓬根对碳、氮的积累明显不同于其它生长时期, 积累了较少的碳、但积累了较多的氮。即使是相同生长时期, 翅碱蓬个体间的根、茎、叶碳、氮含量仍存在一定差异。其离散系数:根碳含量间为2.8%—15.5%;茎碳含量间为2.5%—13.8%;叶碳含量间为3.4%—15.7%;根氮含量间为11.6%—8.4%;茎氮含量间为9.2%—23.5%;叶氮含量间为8.3%—18.2%。从平均含量看, 随着生长时间的延长, 翅碱蓬根、茎碳含量呈增加趋势, 叶碳含量则先降低再增加;根氮含量呈明显的减小趋势, 茎氮含量先降低再增加后再降低, 叶氮含量则先降低再增加。
| 采样时间/d Sampling time |
根Roots | 茎Stems | 叶Leaves | ||||||||||||
| 0 | 49 | 91 | 134 | 179 | 0 | 49 | 91 | 134 | 179 | 0 | 49 | 91 | 134 | 179 | |
| 0 | 30.5±4.7 | c | c | c | c | 36.6±5.0 | a | c | c | c | 28.8±4.5 | c | a | a | — |
| 49 | 38.7±1.6 | b | a | c | 38.5±1.0 | c | c | c | 24.5±1.4 | c | c | — | |||
| 91 | 40.0±1.3 | a | a | 41.4±1.0 | c | c | 29.0±1.0 | c | — | ||||||
| 134 | 39.5±1.8 | a | 40.6±1.0 | c | 30.8±1.9 | — | |||||||||
| 179 | 41.0±1.2 | 43.4±1.5 | — | ||||||||||||
| 独立样本t-检验a:差异不显著;b:差异显著(α=0.05);c:差异极显著(α=0.01) | |||||||||||||||
| 采样时间/d Sampling time |
根Roots | 茎Stems | 叶Leaves | ||||||||||||
| 0 | 49 | 91 | 134 | 179 | 0 | 49 | 91 | 134 | 179 | 0 | 49 | 91 | 134 | 179 | |
| 0 | 1.34±0.24 | c | c | c | c | 1.45±0.16 | c | c | c | c | 1.24±0.22 | c | a | c | — |
| 49 | 0.55±0.06 | a | a | c | 0.70±0.10 | a | c | a | 1.08±0.09 | c | c | — | |||
| 91 | 0.50±0.09 | a | a | 0.79±0.12 | c | a | 1.32±0.11 | a | — | ||||||
| 134 | 0.54±0.10 | a | 1.08±0.10 | c | 1.40±0.14 | — | |||||||||
| 179 | 0.46±0.08 | 0.70±0.16 | — | ||||||||||||
| 独立样本t-检验a:差异不显著;b:差异显著(α=0.05);c:差异极显著(α=0.01) | |||||||||||||||
翅碱蓬根、茎、叶碳、氮含量(Cr)与株高(H)相关生长关系指数方程线性转化式log10 Cr=log10a + b log10 H的拟合方程参数、决定系数R2、拟合方程F-检验F值见表 5。由表 5, 只有根碳、氮含量与株高间存在较好的相关生长关系, 且b<1, 表现为负相关生长关系。叶碳、氮含量与株高间无相关生长关系。对线性化的相关生长方程进行F-检验的结果, 表明只有翅碱蓬根碳、氮含量与株高间相关生长关系极显著(F>F0.01)。对拟合方程参数b(相关生长指数)进行t-检验, 也得到了相同结果。翅碱蓬根碳、氮与株高的相关生长指数b及其95%置信区间分别为0.182(0.122—0.243)、-0.649(-0.916—-0.382)。
| 参数Parameter | 碳Carbon | 氮Nitrogen | ||||
| 根Roots | 茎Stems | 叶Leaves | 根Roots | 茎Stems | 叶Leaves | |
| a | 21.009 | 30.179 | 26.736 | 5.029 | 2.854 | 1.065 |
| b | 0.182 | 0.088 | 0.017 | -0.649 | -0.358 | 0.053 |
| R2 | 0.968 | 0.764 | 0.015 | 0.952 | 0.531 | 0.110 |
| F | 92.15** | 9.71* | 0.03 | 60.01** | 3.39 | 0.25 |
| t(b) | 9.60** | 3.12 | 0.17 | -7.75** | -1.84 | 0.50 |
| *α=0.05;**α=0.01 | ||||||
由于仅翅碱蓬根碳、氮含量与翅碱蓬株高间存在显著的相关生长关系, 因此, 仅计算翅碱蓬根碳、氮相对含量和相对条件因子(表 6)。由表 6, 可见根碳含量较为接近“标准”根碳含量, 而根氮含量则变化较大。相对条件因子也表现出类似情况。显然, 与株高相比, 翅碱蓬根碳、氮积累受到了环境条件限制。按含量-株高关系计算的翅碱蓬根碳、氮估计含量, 即具有相同平均株高的翅碱蓬根碳、氮平均含量与实际测定的根碳、氮平均含量基本相同。
| 采样时间 Sampling time |
相对含量Relative content/% | 相对条件因子Relative condition factor/% | ||
| 根碳Carbon in Root | 根氮Nitrogen in Root | 根碳Carbon in Root | 根氮Nitrogen in Root | |
| 0 | 96.6 | 90.3 | 99.1 | 103.8 |
| 49 | 100.8 | 78.6 | 103.2 | 87.2 |
| 91 | 96.5 | 94.5 | 98.7 | 104.4 |
| 134 | 96.0 | 102.7 | 97.8 | 111.3 |
| 179 | 98.8 | 85.0 | 101.2 | 95.1 |
| Mean±SD | 97.8±2.1 | 90.2±9.2 | 100.0±2.2 | 100.4±9.3 |
由于受机制自我调整、成熟等内在因素和竞争、有限的资源、呼吸、胁迫等外在因素的限制[4], 生物生长速率随生长时间呈下降趋势。翅碱蓬株高平均相对生长率亦如此(图 2)。生物学家采用了多种形式的数学模型描述生物生长随时间的变化曲线。数学模型的待拟合参数个数也不断增加。但随着参数个数增多, 模型趋于复杂, 拟合时, 对数据量的要求也越多。3参数生长方程von Bertalanffy方程、Gompertz方程和Logistic方程相对较简单多用于模拟动物、植物个体甚至是种群生长[2, 4-5, 7]。其中, von Bertalanffy方程的理论基础是生物合成代谢与分解代谢的综合生理过程机制;Logistic方程的相对生长速率(大小增长量与自身大小之比)随生物个体大小线性降低, 曲线拐点位于最终大小的1/2处;Gompertz方程的相对生长速率(大小增长量与自身大小之比)为生长时间的指数函数, 曲线拐点位于最终大小的约1/3处[4]。但采用哪个模型较为合适?基于信息距离、针对小样本量进行偏离修正的AICC以及校正决定系数adj-R2, 因考虑了参数个数多用于模型选择[9], 并在生物生长模型选择中也得到了应用[5]。采用AICC和adj-R2为模型选择依据, Logistic方程可“最佳”地模拟双台河口滨海潮滩湿地翅碱蓬株高生长。张彩琴等的研究表明, 内蒙古典型草原生长的羊草(Leymus chinensis)、大针茅(Stipa grandis)、冷蒿(Artemisia frigida)、冰草(Agropyron michnoi)的生长也符合Logistic增长[7]。但是, 由于各模型的提出均有其自身的理论基础, 一个生长方程不可能适用于描述所有生物的生长过程[4], 为消除采用单一模型推断模型参数的不确定性, MMI方法是一种较为可信的多模型参数推断方法[9], 该方法曾被用于推断鱼类生长的理论渐近体长[6]。本文采用MMI方法推断的翅碱蓬理论渐近株高为(38.11±2.59) cm。
双台河口翅碱蓬株高符合对数正态分布。由于可能受到了盐胁迫、氮、磷等养分缺乏等河口潮滩湿地较为恶劣的环境因素影响, 株高生长分异明显。研究表明, 植物生长具有不确定性, 生长速率极易受环境条件剧烈影响[17]。
翅碱蓬根、茎、叶碳平均含量分别为(37.9±4.4)%、(40.0±3.1)%、(28.3±3.4)%;氮平均含量分别为(0.68±0.35)%、(0.94±0.31)%、(1.26±0.19)%, 低于大多数植物的碳含量(38%—46%)[11]和氮含量(1%—7%)[11]。李征等[28]测定了采集于江苏盐城滨海湿地的盐地碱蓬(S. salsa)叶片碳、氮含量后也得到了类似结果, 叶片碳平均含量26.29±4.29%、氮平均含量(1.156±0.633)%。杭州湾滨海湿地生长的3种草本植物海三棱藨草(Scirpus mariqueter)、糙叶薹草(Carex scabrifolia)和芦苇(Phragmites australis)叶片氮含量分别为(1.169±0.266)%、(1.017±0.153)%和(1.156±0.319)%[34]。福建闽江河口湿地生长的芦苇叶片碳、氮含量分别为(41.96±0.73)%、(3.00±0.42)%, 茎碳、氮含量分别为(41.89±0.23)%、(1.06±0.12)%, 0—15 cm根碳、氮含量分别为(38.59±0.61)%、(1.01±0.04)%, 15—30 cm根碳、氮含量分别为(35.99±1.37)%、(0.93±0.03)%;短叶茳芏(Cyperus malaccensis var. brevifolius)叶片碳、氮含量分别为(41.02±1.17)%、(2.04±0.21)%, 茎碳、氮含量分别为(39.60±0.52)%、(1.21±0.15)%, 0—15 cm根碳、氮含量分别为(39.86±1.67)%、(0.89±0.04)%, 15—30 cm根碳、氮含量分别为(36.89±1.78)%、(0.77±0.04)%。30—60 cm根碳、氮含量分别为(32.17±2.41)%、(0.67±0.04)%[35]。牟晓杰等[36]测定了采集于黄河口湿地的翅碱蓬(S. salsa)根、茎、叶(+果实)氮含量, 发现叶氮含量>1%, 氮含量表现为叶>茎>根, 生长在中潮滩和低潮滩2种生境的碱蓬根、茎、叶氮含量并不一致, 且均有明显的季节变化。Schat的研究显示, 水淹可以降低植被的养分吸收或者减弱植被汲取养分用于生长的能力[37]。
由于植物获得的碳以及氮、磷等养分有限, 植物形成储存物和生长间是矛盾的、竞争的。植物如何分配其获得的碳及氮、磷等养分, 可以指示植物间竞争能力的强弱, 也是减少植物物种竞争相同资源的生物生存策略。植物可以通过降低生长率和改变资源分配型式以响应不利的环境条件, 最大限度地减少单个环境因子对其生长的限制, 因此, 植物可以在生长与储存间最优分配所获得的资源[10, 38]。这种调节不仅有氮、磷等养分根吸收的调节作用[39], 也有根系和叶片之间的碳及氮、磷养分转运的调节作用[10]。并且某些化学信号起到了调节作用[10, 40]。在某些阶段, 植物从环境中快速地吸收碳及氮、磷等养分, 并将新获得的碳和养分由生长或呼吸转向积累或形成储存物。而当植物需要快速生长时, 又可再利用储存物。形成储存物可以降低植物对当时光合作用或从土壤中吸收养分的依赖, 也可使植物从营养生长转为生殖生长[10]。双台河口湿地生长的翅碱蓬以及江苏盐城湿地生长的盐地碱蓬[28]、黄河口湿地生长的翅碱蓬[36]、杭州湾湿地生长的海三棱藨草、糙叶薹草和芦苇[34]营养器官的碳、氮含量随生长时期变化可能是潮滩湿地生长的植物的生存策略之一。
植物的一些属性与其生长间存在相关生长, 即异速生长关系[16, 22], 并且, 机体结构和生理过程中许多特征属性与其以个体质量表示的机体大小间的相关生长指数为1/4的整数倍关系[41]。同样, 作为储存物的碳以及氮、磷等与其生长间亦存在相关生长关系。本项研究中, 只有翅碱蓬根碳、根氮含量与株高间存在显著的相关生长关系, 相关生长因子b分别为0.182、-0.649, 茎、叶碳、氮含量与株高间不具有规律的相关生长关系。研究表明, 植物分配资源被视为是依赖其大小的过程, 即相关生长是生长和分配间的定量关系, 相关生长分析广泛用于研究植物结构和各种生理过程的大小依赖性[42]。资源分配可塑性是植物相关生长轨迹对生存环境的响应[17]。常见的植物资源分配方式源于其异速生长。植物早期发育阶段或低养分条件下, 植物根分配资源最多, 随后是叶, 最后是茎[42]。植物在发育初期大多是根型的, 更大一点后变成茎型。因为, 植物萌发后首先出现根, 之后, 茎长得比根快[43]。
翅碱蓬不同营养器官碳、氮相对含量是基于相对重量[32]提出的, 并与相对条件因子[33]一起, 可以用于间接评价植物的生态学关系和管理策略对生物的影响。由于相对重量测量已经成为一种广义的生物“健康状态”测量的替代指标, 因而广受关注[44]。相对条件因子可用于比较来自于同一批采样的、长度(高度)独立的不同样本间的生长条件比较, 但是, 一般不用于总体间的比较, 总体和物种间的生长条件比较可以采用相对重量[45]。因此, 植物不同器官的碳、氮相对含量可以用于比较不同总体和物种间的生长条件适宜性。相对条件因子也仅用于来自于同一总体的同种植物具有相同高度时样本间的生长条件比较, 局限性较大。生长于双台河口潮滩湿地的翅碱蓬根碳、氮相对含量分别为(97.8±2.1)%、(90.2±9.2)%;相对条件因子分别为(100.0±2.25)%、(100.4±9.3)%。
4 结论3参数方程可以较好地描述双台河口潮滩湿地翅碱蓬的株高生长, 其中Logistic方程是以AICC和adj-R2为准则选择的“最佳”模拟方程——优选方程, 但翅碱蓬株高间分异明显, 可能受环境条件影响较大。采用多模型推断方法可以计算翅碱蓬平均理论渐近株高和非条件标准误。随着翅碱蓬生长, 根、茎碳含量呈增加趋势, 叶碳含量则先降低再增加;根氮含量呈明显的减少趋势, 茎氮含量先降低再增加后再降低, 叶氮含量则先降低再增加。只有根碳、氮含量与株高间呈极显著的相关生长关系。可以采用植物不同营养器官碳、氮相对含量及相对条件因子间接评价植物的生态学关系和植物的环境适应。
| [1] | Hodgson J G, Wilson P J, Hunt R, Grime J P, Thompson K. Allocating C-S-R plant functional types:a soft approach to a hard problem. Oikos, 1999, 85(2): 282–294. DOI:10.2307/3546494 |
| [2] | Yin X Y, Goudriaan J, Lantinga E A, Vos J, Spiertz H J. A flexible sigmoid function of determinate growth. Annals of Botany, 2003, 91(3): 361–371. DOI:10.1093/aob/mcg029 |
| [3] | Erickson R O. Modeling of plant growth. Annual Review of Plant Physiology, 1976, 27: 407–434. DOI:10.1146/annurev.pp.27.060176.002203 |
| [4] | Zeide B. Analysis of growth equations. Forest Science, 1993, 39(3): 594–616. |
| [5] | Hernandez-Llamas A, Ratkowsky D A. Growth of fishes, crustaceans and molluscs:estimation of the von Bertalanffy, Logistic, Gompertz and Richards curves and a new growth model. Marine Ecology Progress Series, 2004, 282: 237–244. DOI:10.3354/meps282237 |
| [6] | Katsanevakis S. Modelling fish growth:Model selection, multi-model inference and model selection uncertainty. Fisheries Research, 2006, 81(2/3): 229–235. |
| [7] | 张彩琴, 杨持. 内蒙古典型草原生长季内不同植物生长动态的模拟. 生态学报, 2007, 27(9): 3618–3629. |
| [8] | Zhu L X, Li L F, Liang Z L. Comparison of six statistical approaches in the selection of appropriate fish growth models. Chinese Journal of Oceanology and Limnology, 2009, 27(3): 457–467. DOI:10.1007/s00343-009-9236-6 |
| [9] | Burnham K P, Anderson D R. Model Selection and Multimodel Inference:A Practical Information-Theoretic Approach. 2nd ed. New York:Springer Science + Business Media, 2002:1-488. |
| [10] | Lambers H, Chapin F S Ⅲ, Pons T L. Plant Physiological Ecology. 2nd ed. New York:Springer, 2008:321-374. |
| [11] | Pate J S, Layzell D B. Carbon and nitrogen partitioning in the whole plant-a thesis based on empirical modeling//Bewley J D, ed. Nitrogen and Carbon Metabolism. Netherlands:Springer, 1981:94-134. |
| [12] | 刘颖慧, 贾海坤, 高琼. 植物同化物分配及其模型研究综述. 生态学报, 2006, 26(6): 1981–1992. |
| [13] | 史作民, 唐敬超, 程瑞梅, 罗达, 刘世荣. 植物叶片氮分配及其影响因子研究进展. 生态学报, 2015, 35(18): 5909–5919. |
| [14] | Niklas K J. A phyletic perspective on the allometry of plant biomass-partitioning patterns and functionally equivalent organ-categories. New Phytologist, 2006, 171(1): 27–40. DOI:10.1111/nph.2006.171.issue-1 |
| [15] | Huxley J S, Teissier G. Terminology of relative growth. Nature, 1936, 137(3471): 780–781. |
| [16] | Enquist B J. Universal scaling in tree and vascular plant allometry:toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiology, 2002, 22(15-16): 1045–1064. DOI:10.1093/treephys/22.15-16.1045 |
| [17] | Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, 2004, 6(4): 207–215. DOI:10.1078/1433-8319-00083 |
| [18] | Niklas K J. Plant allometry, leaf nitrogen and phosphorus stoichiometry, and interspecific trends in annual growth rates. Annals of Botany, 2006, 97(2): 155–163. |
| [19] | Picard N, Rutishauser E, Ploton P, Ngomanda A, Henry M. Should tree biomass allometry be restricted to power models?. Forest Ecology and Management, 2015, 353: 156–163. DOI:10.1016/j.foreco.2015.05.035 |
| [20] | Agutter P S, Tuszynski J A. Analytic theories of allometric scaling. Journal of Experimental Biology, 2011, 214: 1055–1062. DOI:10.1242/jeb.054502 |
| [21] | Price C A, Enquist B J, Savage V M. A general model for allometric covariation in botanical form and function. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(32): 13204–13209. DOI:10.1073/pnas.0702242104 |
| [22] | 韩文轩, 方精云. 幂指数异速生长机制模型综述. 植物生态学报, 2008, 32(4): 951–960. |
| [23] | 赵可夫, 范海, 江行玉, 宋杰. 盐生植物在盐渍土壤改良中的作用. 应用与环境生物学报, 2002, 8(1): 31–35. |
| [24] | 林学政, 陈靠山, 何培青, 沈继红, 黄晓航. 种植盐地碱蓬改良滨海盐渍土对土壤微生物区系的影响. 生态学报, 2006, 26(3): 801–807. |
| [25] | 邹桂梅, 苏德荣, 黄明勇, 刘虎, 蔡飞. 人工种植盐地碱蓬改良吹填土的试验研究. 草业科学, 2010, 27(4): 51–56. |
| [26] | 何洁, 王庆芝, 吉志新, 陈旭, 刘长发, 周一兵. 翅碱蓬-沙蚕对Cu污染沉积质理化性质的影响研究. 环境污染与防治, 2014, 36(11): 22–29. DOI:10.3969/j.issn.1001-3865.2014.11.005 |
| [27] | 许崇彦, 刘宪斌, 刘占广, 王娟, 姜中鹏, 曹佳莲. 翅碱蓬对石油烃污染的海岸带修复的初步研究. 安全与环境学报, 2007, 7(1): 37–39. |
| [28] | 李征, 韩琳, 刘玉虹, 安树青, 冷欣. 滨海盐地碱蓬不同生长阶段叶片C、N、P化学计量特征. 植物生态学报, 2012, 36(10): 1054–1061. |
| [29] | Mou X J, Sun Z G, Wang L L, Wang C Y. Nitrogen cycle of a typical Suaeda salsa marsh ecosystem in the Yellow River estuary. Journal of Environmental Sciences, 2011, 23(6): 958–967. DOI:10.1016/S1001-0742(10)60530-X |
| [30] | 孙红叶, 张希明, 李利, 马建兵. 塔里木盆地南缘不同生境盐生草种群分布特征及地上生物量初步估测. 干旱区资源与环境, 2008, 22(4): 193–197. |
| [31] | Voje K L, Hansen T F, Egset C K, Bolstad G H, Pélabon C. Allometric constraints and the evolution of allometry. Evolution, 2014, 68(3): 866–885. DOI:10.1111/evo.12312 |
| [32] | Murphy B R, Brown M L, Springer T A. Evaluation of the relative weight (Wr) index, with new applications to walleye. North American Journal of Fisheries Management, 1990, 10(1): 85–97. DOI:10.1577/1548-8675(1990)010<0085:EOTRWW>2.3.CO;2 |
| [33] | Le Cren E D. The length-weight relationship and seasonal cycle in gonad weight and condition in the perch (Perca fluviatilis). Journal of Animal Ecology, 1951, 20(2): 201–219. DOI:10.2307/1540 |
| [34] | 吴统贵, 吴明, 刘丽, 萧江华. 杭州湾滨海湿地3种草本植物叶片N、P化学计量学的季节变化. 植物生态学报, 2010, 34(1): 23–28. |
| [35] | 王维奇, 徐玲琳, 曾从盛, 仝川, 张林海. 河口湿地植物活体-枯落物-土壤的碳氮磷生态化学计量特征. 生态学报, 2011, 31(23): 7119–7124. |
| [36] | 牟晓杰, 孙志高, 王玲玲, 董洪芳. 黄河口滨岸潮滩不同生境下翅碱蓬氮的累积与分配特征. 湿地科学, 2010, 8(1): 57–66. |
| [37] | Schat H. A comparative ecophysiological study on the effects of waterlogging and submergence on dune slack plants:growth, survival and mineral nutrition in sand culture experiments. Oecologia, 1984, 62(2): 279–286. DOI:10.1007/BF00379027 |
| [38] | Chiariello N R, Mooney H A, Williams K. Growth, carbon allocation and cost of plant tissues//Pearcy R W, Ehleringer J R, Mooney H A, Rundel P W, eds. Plant Physiological Ecology:Field methods and instrumentation. Netherlands:Springer, 2000:327-365. |
| [39] | Gojon A, Nacry P, Jean-Claude D. Root uptake regulation:a central process for NPS homeostasis in plants. Current Opinion in Plant Biology, 2009, 12(3): 328–338. DOI:10.1016/j.pbi.2009.04.015 |
| [40] | 李冀南, 李朴芳, 孔海燕, 熊俊兰, 王绍明, 熊友才. 干旱胁迫下植物根源化学信号研究进展. 生态学报, 2011, 31(9): 2610–2620. |
| [41] | 韩文轩, 方精云. 相关生长关系与生态学研究中的尺度转换. 北京大学学报:自然科学版, 2003, 39(4): 583–593. |
| [42] | Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(2): 115–127. DOI:10.1078/1433-8319-00007 |
| [43] | 陆霞梅, 周长芳, 安树青, 方超, 赵晖, 杨茜, 颜超. 植物的表型可塑性、异速生长及其入侵能力. 生态学杂志, 2007, 26(9): 1438–1444. |
| [44] | Blackwell B G, Brown M L, Willis D W. Relative weight (Wr) status and current use in fisheries assessment and management. Reviews in Fisheries Science, 2000, 8(1): 1–44. DOI:10.1080/10641260091129161 |
| [45] | Froese R. Cube law, condition factor and weight-length relationships:history, meta-analysis and recommendations. Journal of Applied Ichthyology, 2006, 22(4): 241–253. DOI:10.1111/jai.2006.22.issue-4 |