 2016, Vol. 36
2016, Vol. 36文章信息
- 魏亚楠, 党政武, 陈立, 方宇凌, 孔祥波, 胡春祥, 赵莉蔺
- WEI Yanan, DANG Zhengwu, CHEN Li, FANG Yuling, KONG Xiangbo, HU Chunxiang, ZHAO Lilin
- 线虫蛔甙(Ascarosides)信息素研究进展
- Research progress on pheromone ascarosides in nematodes
- 生态学报, 2016, 36(6): 1501-1514
- Acta Ecologica Sinica, 2016, 36(6): 1501-1514
- http://dx.doi.org/10.5846/stxb201409011734
-
文章历史
- 收稿日期: 2014-09-01
- 网络出版日期: 2015-07-29






2. 中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室, 北京 100101;
3. 商洛市林木病虫防治检疫站, 商洛 726000;
4. 中国林业科学研究院森林生态环境与环保研究所国家林业局森林保护重点实验室, 北京 100091
2. State Key Laboratory of Integrated Management of Pest Insects and Rodents, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China;
3. Shangluo Forest Pests Control and Quarantine Station, Shangluo 726000, China;
4. Key Laboratory of Forest Protection of State Forestry Administration, Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
线虫是一类低等无脊椎动物,在自然界分布很广[1]。一部分线虫可以独立生存,称为自由生活线虫;另一部分采用寄生策略,大部分以植物和动物为寄主,对特殊食物、传播媒介和资源加以利用。因为这些线虫通常生活在土壤或寄生物中,没有适宜的视觉或听觉系统,接收环境信号的重要途径就是精细的化学感受系统[2, 3, 4, 5]。通过进化,化学感受系统越来越精细,不仅能引起线虫行为的改变,还能调控线虫的发育途径。在不良环境中,线虫进入扩散周期,形成滞育虫态,从而进一步扩散传播到新的适宜环境[2, 4, 5]。
秀丽隐杆线虫Caenorhabditis elegans一直是线虫化感研究最主要的模式生物,但主要集中在基因调控水平[2, 3, 4, 5],信息素鉴定和结构研究进展缓慢。直到2005年,Jeong等[6]在秀丽隐杆线虫中提取并鉴定出一类诱导滞育虫态形成的新型化合物——蛔甙(ascarosides)。从此,蛔甙的相关研究取得了一系列突破性进展。人们定义蛔甙为线虫的特定信号分子,由脂肪酸等侧链链接到双脱氧蛔糖基(dideoxysugar ascarolyse)上组成。通过侧链长度的改变以及衍生物的差异生成结构多样的蛔甙[7],用以调节线虫的聚集、交配、多尔(dauer)形成等行为。
最近发现,各种寄生线虫中普遍存在蛔甙成分。众所周知,各类寄生线虫危害可造成重大农业经济损失及全球性人类疾病。如果不同线虫种类对不同蛔甙能够特异性识别,那么蛔甙有望发展成为新型药物特异干扰线虫的繁殖和生存。深入研究蛔甙对线虫行为和发育的调节,对于控制重大农业经济损失及人类疾病具有举足轻重的意义。因此,本文对近年来线虫蛔甙信息素研究的突破性进展进行了总结。
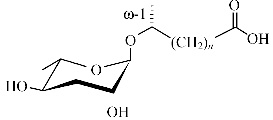
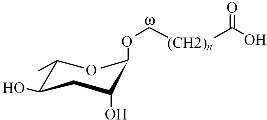
1 秀丽隐杆线虫蛔甙结构鉴定与功能线虫对蛔甙这种小分子信号的利用,让人联想到细菌群体感应系统通过检测复杂的环境条件来调节自身的行为和发展。蛔甙信息素主结构均由蛔糖基连接一个脂肪酸侧链形成,因侧链的长度和衍生物的不同而形成结构和功能的多样性。蛔甙的总体结构见图 1。根据发现的先后顺序,依次命名为ascr#1,2,3,…,n。不同的蛔甙或蛔甙组合调节不同的表型特征,在化学结构上即使很小的差异都可能极大地改变线虫的生理活动[8]。蛔甙的活性和特异性依赖于化学结构和浓度。因此蛔甙信号特异性可以通过定性、定量来表达。

|
| 图1 蛔甙结构示意图 Fig.1 Overall structure of ascarosides 蛔糖(ascarylose sugar)编号的位置显示已知的修饰官能团 |
Choe[9]总结秀丽隐杆线虫包含三类蛔甙结构:一种是结构简单的蛔甙,简写为ascr,一种是吲哚蛔甙(indole ascarosides),简写为icas,另外一种是ω-1氧化ascr常伴随的ω氧化异构体,简写为oscr(化学结构见表 4)。
1.1 诱导秀丽隐杆线虫多尔幼虫形成的信息素结构及其鉴定线虫信息素蛔甙是在多尔幼虫形成研究中被发现的。多尔是秀丽隐杆线虫幼虫在生长发育过程中出现的滞育虫态,此虫态的形成与多尔信息素诱导密切相关。
不论自然界中的环境好坏,秀丽隐杆线虫都能适应并生存。在环境适宜的条件下,秀丽隐杆线虫仅需3—5 d就能由受精卵经过L1—L4四龄幼虫阶段发育为成虫,并产生约300个后代。在L1或L2初期,当遇到食物缺乏、种群密度增加、温度升高等恶劣环境时,秀丽隐杆线虫将进入滞育,形成持久型L3幼虫,称为多尔幼虫[10]。线虫进入滞育阶段会停止取食和发育[11]。滞育阶段能维持数月,当环境适宜时(如食物充足、低浓度信息素),线虫恢复发育,蜕皮成繁殖型L4幼虫,进入繁殖周期[11, 12]。
早在20世纪80年代,科学家们就发现秀丽隐杆线虫的多尔形成是受化学信息素调控的,认为此类信息素是亲水性的双脱氧蛔糖基衍生物,但一直难以明确其结构组成[13, 14]。直到2005年,Jeong等突破了提取和鉴定方法,在300 L大型发酵罐中进行了秀丽隐杆线虫的大规模液体培养,并用乙酸乙酯提取液体培养液进行多尔信息素的活性物质分离纯化。然后进行色谱分离,结合生物活性测试获得具有显著诱导效果馏分后,用标准核磁共振NMR(nuclear magnetic resonance)和质谱技术鉴定出一类新型物质——ascr#1(即daumone)[6]。Butcher等[15]用类似的方法通过多尔恢复试验纯化鉴定了ascr#2,并用核磁共振谱分析找到另一个蛔甙,即ascr#3。Ascr#1及ascr#3的荧光探针显示,在多尔形成过程中集中在咽(pharynx)表皮,ascr#3探针甚至能移动到化感器神经元(amphid neurons)[16]。
随后,Gallo和Riddle[17]比较了ascr#1合成物[6]和秀丽隐杆线虫提取物[18]的生理特性,发现ascr#1并不是形成多尔信息素的主要成分,而且Jeong已报道的浓度[6]对秀丽隐杆线虫有毒,接触后会降低其寿命[17]。
虽然业界对Jeong的研究有所质疑,但它依然为蛔甙的后续研究开辟了一个新的天然产物研究方向,大规模的蛔甙分离鉴定工作展开,甚至发现了蛔甙衍生物,如icas#9[19]。这些蛔甙在不同的发展阶段产生,并在各种恶劣条件诱导下增加[20]。多个蛔甙(ascr#1-3,5,8,icas#9)(表 1)协同作用形成一个诱导型的混合物。其中,一种侧链为3-羟丙酸(3-hydroxypropionatea)的蛔甙是多尔信息素中一个非常重要的成分[21],ascr#2和ascr#3比最初确定的ascr#1功能更强[15,20- 21]。诱导多尔幼虫形成的蛔甙结构见表 1。
| 名称 Name | 又名 Other name(s) | 化学结构 Chemical structure | 诱导多尔形成活性 Dauer activity | 参考文献 References |
| ascr#1 | ascaroside C7/asc-C7/ daumone-1 | 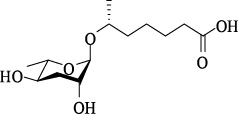 | 非常低 | [6] |
| ascr#2 | ascaroside C6/ daumone-2 | 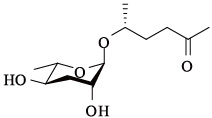 | 高 | [15] |
| ascr#3 | ascaroside C9/ asc-△C9/daumone-3 | 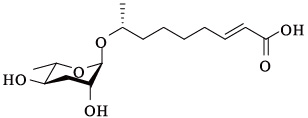 | 高 | [15] |
| ascr#4 | — |  | 低 | [22] |
| ascr#5 | ascaroside C3/ asc-ωC3/daumone-5 | 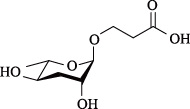 | 高,与ascr# 2,3 表现协同作用 | [21] |
| ascr#8 | — | 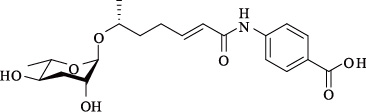 | 适中 | [23] |
| icas#9 | indolecarboxyl ascaroside C5/ascaroside C5/ IC-asc-C5 | 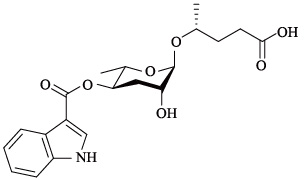 | 适中,反应为钟形 | [24] |
最近的研究发现,低浓度蛔甙能够诱导秀丽隐杆线虫的交配行为。科学家们通过多种手段,如核磁共振光谱分析(NMR spectroscopic analyses)方法及二维核磁共振光谱DANS(differential analysis of 2D NMR spectra)鉴定方法得到能够诱导交配活性的蛔甙结构,包括ascr#4[22],ascr#6.1,6.2,8[23]。
当Jeong等人都使用标准液体细菌和线虫的混合培养液作为原材料时,Srinivasan等则将特定发育阶段的线虫转移到水中来专门收集线虫的化合物,以防止细菌污染[22]。这种策略可达到3个重要优势目标:首先,它大大简化了雄虫的趋化生物测定,因为线虫也会被很多细菌小分子所吸引而增加生测难度;其次,它简化了原材料的化学成分鉴定;最后,明确了虫态特异性。因为雄虫活动只出现在L4龄幼虫、年轻成虫以及成虫3个阶段[22],因此虫态特异性对性信息素的鉴定尤为重要,这种策略集合了每个发育阶段所产生的特定物质组成,而不是同步培养时随着时间推移从而积累所有的化合物。
吸引雄性的信息素是由一个以上的化合物组成的,单个的色谱成分不能充分引起特异雄性的反应,但重组成分却能使其恢复活力[22]。其中一个主要成分是被葡萄糖修饰的ascr#2衍生物,这个化合物称作ascr#4。Ascr#2、ascr#3和ascr#4表现为协同作用,在pmol/L浓度下活性强,特别吸引雄性。
从此,鉴定蛔甙的手段越来越先进,以Paul W. Sternberg[23]为首的实验室采用二维核磁共振光谱DANS鉴定方法得到另一些蛔甙结构,包括能够诱导交配活性的ascr#6.1、ascr#6.2,以及功能未知的ascr#7(结构见表 4)。他们通过这种方法实现了蛔甙的核磁共振痕量检测,并且通过DANS差异分析,简化了连接的小分子代谢物鉴定和它们的生物学功能研究[23]。
秀丽隐杆线虫雄虫也会产生蛔甙,以饱和ascr#10为主,而雌雄同体线虫产生的蛔甙为α,β-不饱和蛔糖苷,ascr#3。虽然两者的化学结构只有微小差别,但生理活性有很大不同:雌雄同体线虫产生的ascr#3能够排斥其他雌雄同体个体、吸引雄虫,而雄虫产生的ascr#10能够强烈吸引雌雄同体线虫[25]。同时,Srinvisan等[22]的研究表明,很多蛔甙的混合物,特别是ascr#3和ascr#8,能够在超低浓度下吸引强壮的雄性个体。在较高浓度下,相同的混合物对雄性丧失吸引力,取而代之的是对雌雄同体的排斥。蛔甙性信息素的结构见表 2。
| 名称 Name | 又名 Other name(s) | 化学结构 Chemical structure | 交配活性 Mating activity | 参考文献 References |
| ascr#2 | ascaroside C6/ daumone-2 |  | 适中,与ascr#3,4表现为协同作用 | [15] |
| ascr#3 | ascaroside C9/ asc-△C9/ daumone-3 | 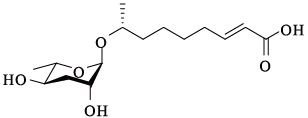 | 高,与ascr#2,4表现为协同作用 | [15] |
| ascr#4 | daumone-4 | 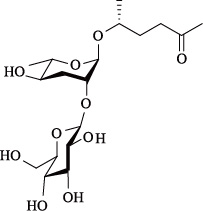 | 本身没有活性,但与ascr#2,3表现为协同作用 | [22] |
| ascr#6.1 | — | 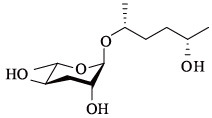 | 适中 | [23] |
| ascr#6.2 | — | 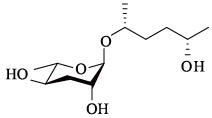 | 未证实 | [23] |
| ascr#8 | — | 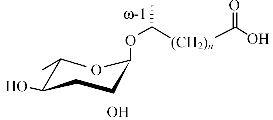 | 高,与ascr#2,3表现为协同作用 | [23] |
| ascr#10 | asc-C9 | 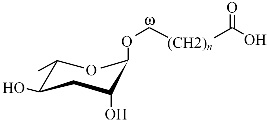 | 高 | [25] |
蛔甙还能够诱导秀丽隐杆线虫的聚集行为。这类蛔甙含有色氨酸吲哚衍生物(tryptophan-derived indolemoeities-indole ascarosides),其中一个命名为indolecarboxyl-ascaroside C5,即icas#9[19]。秀丽隐杆线虫的雌雄同体能够被浓度为fmol/L到pmol/L的icas#3(表 3)和icas#9强烈吸引,造成聚集现象[26]。
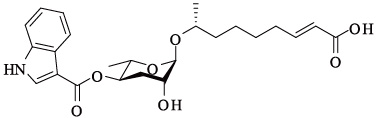
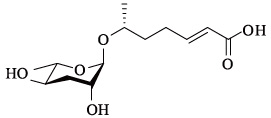
蛔甙还能调节其它行为和趋化反应。如最近的几项研究[22, 26, 27, 28]都提到了调控不同行为的蛔甙信号,包括特异性别吸引、排斥、聚合以及嗅觉可塑性。蛔甙除了直接影响线虫的行为之外,还能引起对其他异嗅物质的趋化反应,如野生型线虫对苯甲醛气味的反应依赖于种群密度[27]。值得注意的是,诱导这种行为产生的部分蛔甙与调节多尔形成的蛔甙是相同的,诱导多尔形成需要的浓度为nmol/L—μmol/L级别,而行为反应的浓度可能只需要fmol/L—pmol/L。目前,除了上述一些短链蛔甙功能被明确,表 4列出的长链蛔甙功能还未知,尚有很大的研究潜力和空间。
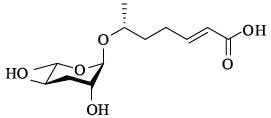
总之,线虫蛔甙方面的最新发现,为研究信息素交流及其演变提供了前所未有的实验依据。在功能上,蛔甙信息素主要分为多尔幼虫信息素、性信息素、聚集信息素等三类。蛔甙根据结构的不同功能也有所不同。结构相同的信息素在功能上又有重叠,例如ascr#1,#2,#3,#5,#8既是多尔诱导信息素也是雄性诱导信息素,而icas#9既是多尔诱导信息素又是聚集信息素。秀丽隐杆线虫的其他一些蛔甙信息素(见本文1.4)功能还有待进一步深入探索。
2 寄生线虫蛔甙结构鉴定与功能属于线虫门的蛔虫属(Ascaris)也存在蛔甙,包括马蛔虫Parascaris equorum[29]、人体蛔虫Ascaris lumbricoides、猪蛔虫Ascaris suum[30, 31, 32]。主要存在于蛔虫属卵中,抵抗由质变而导致的生理退化,使蛔虫卵在潮湿的土壤中存活数年[33]。蛔虫卵具有3层结构,外层是蛋白质、中间是甲壳素,蛔甙包被在内部。存在于最内层的是蛔甙长链混和物,具有非渗透性(结构见图 2)[34]。

蛔甙既存在于蛔虫属,也存在于秀丽隐杆线虫,二者虽然都属于线虫门,但蛔虫目Ascaridida和小杆目Rhabditida在形态学、生理学特征上有显著区别[32],导致多数人认为寄生线虫的长链蛔甙与秀丽隐杆线虫的蛔甙信息素组成是不同的。直到2009年才研究发现两者是非常相似的[8, 9],其功能上也承担了信息传递、幼虫滞育的功能。
蛔甙结构决定其在蛔虫属卵发育的特定功能。图 2为猪蛔虫Ascaris suum卵中的6种长链蛔甙结构,类似的长链结构在秀丽隐杆线虫N2(野生型)和daf-22中有报道[23]。蛔虫和秀丽隐杆线虫的蛔甙在功能上有异同之处:秀丽隐杆线虫从卵中孵化的阶段作为L1幼虫,而猪蛔虫在卵中要经历两次蜕皮。当蛔虫的卵被携带进入新的寄主时,孵化得到L3龄幼虫,类似于秀丽隐杆线虫的多尔幼虫阶段[35]。卵中的蛔甙与幼虫有直接关系,与秀丽隐杆线虫的蛔甙在多尔形成(或至少在多尔维持)上扮演着相同角色[36]。因此,调查研究动物寄生线虫的蛔甙功能将为控制线虫的侵染提供新见解。
蛔甙结构特征在不同线虫种类进化中出现差异。Choe等[9]通过对比自由生活线虫、动物、昆虫和植物寄生性线虫等在进化关系上较远的19种线虫的蛔甙结构特征[37](表 5),发现不同线虫种类演变为占据不同生态位的种群,尽管都显示出利用蛔甙作为信号分子的保守性,但在蛔甙种类和组成上有很大差异。甚至4种因自然隔离而导致基因差异的秀丽隐杆线虫产生的蛔甙也是不同的。当然,这些种内差异与种间差异相比是非常小的。
| 生活方式 Lifestyle | 名称 Name | 性别 Reproductive mode | 分支 Clade | 蛔甙种类 Species ofascarosides |
| 食细菌和/或真菌 Bacteriovore and/or | 全齿复活线虫 Panagrellus redivivus | ♂ ♀ | 10 | ascr#1,3,9,10,14,18 |
| fungivore | 类圆小杆线虫 Pelodera strongyloides | ♂ ♀ | 9 | ascr#18,20,22,24,26 |
| 秀丽隐杆线虫 Caenorhabditis elegans | 0 | 9 | ascr#1,2,5,7,9,10,12,13, 18,26; icas#9; oscr#9,10 | |
| Caenorhabditis sp. | ♂ ♀ | 9 | ascr#1,7,9,10,12,13,14, 18;icas#9; oscr#9 | |
| Caenorhabditis sp.7 (D) | ♂ ♀ | 9 | ascr#1,3,10,12,14,18; icas#9 | |
| Rhabditis sp. | 0 | 9 | ascr#1,9,10,12; oscr#9,10 | |
| 植物寄生Plant parasite | 穿刺短体线虫 Pratylenchus penetrans | ♂ ♀ | 12 | 未知 |
| 昆虫病原寄生 Entomopathogenic insect | 嗜菌异小杆线虫 Heterorhabditis bacteriophora | ♂ ♀ 0 | 9 | ascr#9,12,18,20,22,24,26 |
| parasite | Oscheius carolinensis (A) | ♂ ♀ | 9 | ascr#1,9,10,14,16,18,22,26; icas#9 |
| Oscheius carolinensis (IJ) | ♂ ♀ | 9 | ascr#1,7,9,10,18,22; icas#9 | |
| 小卷蛾斯氏线虫(A) Steinernema carpocapsae (A) | ♂ ♀ | 10 | ascr#1,9,10,11,12,18 | |
| 小卷蛾斯氏线虫(IJ) Steinernema carpocapsae (IJ) | ♂ ♀ | 10 | ascr#9,10 | |
| Steinernema scapterisci (IJ) | ♂ ♀ | 10 | ascr#9,10,11,14 | |
| Steinernema riobrave (IJ) | ♂ ♀ | 10 | ascr#9,10,14,16,18 | |
| Steinernema glaseri (A) | ♂ ♀ | 10 | ascr#1,9,12 | |
| Steinernema glaseri (IJ) | ♂ ♀ | 10 | ascr#9,11,12 | |
| 非昆虫病原寄生 Non-entomopathogenic | 应氏罗曼索线虫(A) Romanomermis iyengari (A) | ♂ ♀ | 2 | 未知 |
| insect parasite | 应氏罗曼索线虫(IJ) Romanomermis iyengari (IJ) | ♂ ♀ | 2 | 未知 |
| 食蚊罗曼索线虫(A) Romanomermis culicivorax (A) | ♂ ♀ | 2 | 未知 | |
| 与昆虫相关,食细菌或 | Oscheius tipulae | 0 | 9 | ascr#9,12 |
| 真菌 | Pristionchus pacificus | 0 | 9 | ascr#1,9,12 |
| Insect-associated,bacteriovore/fungivore | Koernia sp. | ♂ ♀ | 9 | ascr#1,3,14 |
| 脊椎动物寄生 Vertebrate parasite | 巴西日圆线虫(A) Nippostrongylus brasiliensis (A) | ♂ ♀ | 9 | ascr#3,7,10 |
| 巴西日圆线虫(IJ) Nippostrongylus brasiliensis (IJ) | ♂ ♀ | 9 | ascr#3,10 | |
| 猪蛔虫 Ascaris suum | ♂ ♀ | 8 | 未知 | |
| A: 成虫,IJ: 感染性幼虫,D: 多尔幼虫,♂: 雄虫,♀: 雌虫,0: 雌雄同体; 部分种对应中文种名未知 | ||||
19种线虫样本中都含有蛔甙,且由两个或两个以上化合物组成[9]。除了小杆科Rhabditidae的几个未知种:Caenorhabditis sp.7和Rhabditis sp.(突变种)产生的蛔甙在碳末端上连接着蛔糖(即“ω”位置,如oscr#9和oscr#10)[7],大多数线虫产生的蛔甙都在倒数第2个碳上连接着蛔糖(即“ω-1”位置)。同样地,作为秀丽隐杆线虫聚集信号的吲哚蛔甙——icas#9[26],也只在小杆科的几个种:Caenorhabditis spp.(多个未知种)和Oscheius carolinensis中出现。这些发现表示,在碳末端上连接蛔糖的蛔甙或是蛔甙的吲哚羰基衍生物可能代表着第9分支(其中包括小杆科家族的成员)衍生的一个新的里程碑[37]。他们还发现第9分支的线虫(木耳线虫属未知种Pelodera sp.,小杆线虫属未知种Oscheius.sp和异小杆线虫属未知种Heterorhabditi sp.)能大量产生含有12—15个碳原子的长链蛔甙,而第10分支的线虫(昆虫病原斯氏线虫属未知种Steinernema spp.和Panagrellus sp.)不产生这些蛔甙。值得注意的是,小杆线虫属Oscheius和斯式线虫科(Steinernematidae)、异小杆线虫科(Heterorhabditidae)的成员包含了所有已知的昆虫寄生线虫[38],它们进化为不同分支是由蛔甙的侧链长度决定的,而不是生活方式。具有蛔甙相同结构特征的种有着相似的生态位[9]。因此推断,蛔甙生物合成的模式似乎与亲缘关系以及生活方式或生态位在某种程度上是相关的。
综上所述,通过蛔甙在蛔虫属等寄生性线虫中的不同结构研究,发现不同物种通过特异蛔甙信号进行识别,进而干扰蛔虫属、寄生性线虫的繁殖和生存,但是目前对于不同寄生线虫种类的滞育诱导信息素、性信息素、聚集信息素功能的特异性蛔甙组分尚待进行深入研究。
3 蛔甙生物合成与代谢调控从结构研究表明蛔甙信息素组分不同,但功能相似。蛔甙在生物合成和代谢通路中所处的位点、生理生化功能不同。蛔甙生物合成及代谢包括对糖类和脂肪酸两方面的调控。在生物合成方面已明确了在脂肪链合成过程中的两个蛋白参与其代谢过程,其他方面的功能有待进一步研究确定。
3.1 蛔甙生物合成蛔甙的合成主要是通过编码蛋白的两种基因daf-22(thiolase)和dhs-28(dehydrogenase)发现的。Daf-22编码一种人体固醇载体蛋白(human sterol carrier protein)的同系物SCPx(sterol carrier protein X)[15]。这种酶能催化过氧化物酶反应中脂肪酸β-氧化(β-oxidation)反应的最后一步[19]。Dhs-28基因编码人类双功能蛋白(human D-bifunctional protein)的一个同族蛋白质,这种蛋白质只能合成SCPx的上游部分。该蛋白在肠道中表达,而肠道是秀丽隐杆线虫中蛔甙的合成位点[19]。这个途径使得短链、中链、长链的脂肪酸降解为乙酰辅酶A(acetyl-CoA)和胆汁酸中间体(bile acid intermediates)。将daf-22和dhs-28线虫在液体培养基下培养10d、20d,发现培养了10d的两种突变体多尔形成都不活跃,而培养20d的突变体在多尔形成中十分活跃[15]。培养20d的dhs-28线虫中发现了一些长链蛔甙,其中最长的含有15个碳,表明这些蛔甙就是信息素生物合成的中间体。蛔甙生物合成的许多步骤仍有待阐明,包括长链蛔甙的初始形成阶段以及特定官能团如ascr # 8中的芳酰胺(aromatic a-mide)[23]以及ascr # 9中的吲哚(indole)官能团[24]的细节性描述。蛔甙生化合成通路如何使其生物学功能得以实现也有待进一步研究。
3.2 蛔甙代谢调控通过生化代谢过程,形成蛔甙的前体分子,为蛔甙的生物合成提供原料。秀丽隐杆线虫繁殖周期线虫靠取食细菌来摄取能量和生物合成的前体分子[39]。糖类和一些核酸、氨基酸首先通过糖酵解途径代谢产生乙酰辅酶A;而脂类物质则通过β-氧化代谢产生乙酰辅酶A。乙酰辅酶A再通过三羧酸循环TCA(tricarboxylic acid)和氧化磷酸化(oxidative phosphorylation)产生ATP(adenosine triphosphate)[39, 40]。在秀丽隐杆线虫繁殖型线虫中,糖酵解、β-氧化、三羧酸循环和氧化磷酸化这四种代谢途径都处于活跃状态。而在多尔幼虫阶段,由于线虫不再进食,必须依靠身体内部储存的物质为机体提供能量和生物合成前体。在多尔形成过程中,最主要的能量物质是那些在皮下和肠道部位积累的脂肪。这些脂肪主要以三酰甘油(triacylglycerides)的形式存在,由3个长链脂肪酸分子以酰基的方式连接到甘油的三个羟基上。不同于繁殖型线虫,多尔必须依靠脂肪来提供能量和生物合成前体,这就需要脂肪酸通过β-氧化产生乙酰辅酶A进入后续TCA循环和氧化磷酸化产生ATP;同时通过β-氧化、乙醛酸循环(glyoxylate cycle)和糖异生(gluconeogenesis)途径来产生供蛔甙生物合成用的前体分子[39, 40](图 3)。
4 蛔甙化学感受信号途径
蛔甙信号传导途径解释了其信息素功能的实现方式。线虫通过对外界化学信号的接收、传递,进而改变自身的行为和发育,并且通常利用化学感受和温度感受来控制扩散周期的形成。目前最重要的任务就是阐明线虫的感觉系统是如何感知并集成复杂的蛔甙的组合,进而引起特定发育和行为变化的。
性信息及聚集信息行为的实现是通过神经传递通路实现的。秀丽隐杆线虫神经系统能够传导及感应蛔甙信息素来指导其交配和聚集行为[22, 26, 27, 28]。其中,化学感受神经元用来识别种内信息物质。翅状神经元ADF(actin depolymerizing factor)和ASI(acid-sensing ion)控制线虫侵染虫态形成并进入扩散周期。成对的感觉神经元控制从扩散周期恢复发育状态,转入繁殖周期[42]。这些神经元主要存在于头感器、尾感器以及内/外唇和中食道球与后食道球之间[43]。这些神经元都有树突状结构,一直延伸到头部开口处。
多尔滞育现象也是通过神经传递系统信号传递实现的。De Bono等[44]发现了npr-1(neuropeptide receptor)的无效突变(null mutation)在蛔甙化感中的功能。Npr-1基因作用的初始位点是RMG(relative maturity group)中间神经元(interneuron)[28]。RMG形成的缺口处连接着负责ascr#3特异性雄性吸引的ASK(amphid single-ciliated sensory neuron type K)感觉神经元[22]。ASK神经元中钙含量减少,而后形成ascr#2,ascr#3和ascr#5三者的混合物,同时ASK产生蛔甙的信号能够传递到其他细胞[28]。RMG的高活性能促进群体聚集行为,还能增强ASK对蛔甙的反应,而NPR-1的高活性能降低这种反应[28]。因此在特化虫株中,蛔甙通过调控聚集行为来增强之前建立的多尔滞育和交配功能[36]。
蛔甙的化学感受生理生化过程包括以下两个步骤[42]:①化学信号分子与化感器分泌蛋白相接触并结合[3],这些分泌蛋白的特殊性质能够增强线虫嗅觉感受效果;②通过特殊转换途径,外界化学分子信息转换为神经信号,被线虫化感器神经元识别:神经元和化学信息分子之间相互作用,刺激GTP(guanosine triphosphate)联接的蛋白(G蛋白)活性,通过G蛋白信号途径转换为神经信号[4, 45, 46]。G蛋白偶联受体家族中一些特异化感受体基因用来响应蛔甙[47, 48]。Zwaal等明确了两个G蛋白的α亚基,GPA-2和GPA-3参与信号转导[49]。McGrath等[48]分析并演示了在多尔形成过程中,G蛋白偶联化感受体作为蛔甙受体被基因srg(serpentine receptor class g)编码,而且在实验室有利条件下终止多尔形成。
蛔甙在信号传递过程中受相关基因的调控,当线虫遇到化学信号刺激时,G蛋白偶联化感受体与配体化学分子结合,启动鸟苷酸环化酶途径,导致神经元细胞内cGMP(cyclic guanosine monophosphate)浓度升高,激活cGMP-门控离子通道(cGMP-gated ion channel),进而使细胞去极化产生钠离子和钙离子内流。经过一系列细胞核转录事件,产生Insulin-like和TGF-β(transforming growth factor-beta)通路的信号分子。这些信号分子在内分泌细胞表面与各自受体结合,分别启动相应通路信号转导。最终两条信号通路共同调控一种细胞色素P450(cytochrome P450)基因daf-9的表达;此基因负责核受体DAF-12(the nuclear hormone receptor)的配体DA(dafachronic acid)生物合成的最后一步。DA与靶组织DAF-12结合后,促进繁殖发育,抑制dauer形成;相反,当DA含量下降时,空载核受体DAF-12则促进多尔形成,抑制繁殖发育[50]。Insulin/IGF-1(insulin-like growth factor I)和TGF-β信号通路在动物寄生性线虫中有少量研究,结果表明,Insulin信号通路在寄生性线虫中作用是保守的,而TGF-β信号通路可能起不同于秀丽隐杆线虫的作用[51]。
可形成蛔甙受体的辅基库有待深入研究。可与蛔甙匹配的受体库应考虑到单个神经元表达多种化学感受器,从线虫的化学感受基因种类、功能等方面进行挖掘[52]。
5 蛔甙信息调节模式蛔甙的信号传导原理有了一定的研究,但信息调节过程除了内部基因、生理调控、生化代谢等影响,也受环境因子的限制。环境对秀丽隐杆线虫的蛔甙的表达、蛔甙释放机制(例如组成型分泌、调节分泌、排便、以及其他可能的机制)、蛔甙家族信号分子的协同机制并未获知。研究仅发现协同作用的10个或更多蛔甙分泌自几个少数的细胞,当达到一定浓度才能实现调节,对于揭密蛔甙的上述多重影响任重道远,明确蛔甙信息调节仍要深入研究。
现研究发现雌、雄虫蛔甙信息素水平有差异。Edison[36]认为蛔甙信息素能够增加雄虫出现的频率,而且效果会随着本地整体信息素水平的升高而升高。图 4阐明了秀丽隐杆线虫生态循环中雌雄虫受蛔甙影响的几个关键点。雌雄同体发展到即将交配阶段时会释放少量不同的蛔甙,在相对统一的多尔信息素水平下,为雄虫寻找多产的雌雄同体提供了方向性[36]。同时,蛔甙通过组成型表达来监测种群密度,也能根据环境或发育因素进行调节。这为环境条件恶化时秀丽隐杆线虫的雌雄同体与雄性的异型杂交提供了一种机制。当信息素增加,食物减少时,产生的异型杂交后代将成为多尔幼虫。该多尔状态会导致雄虫的增加[53],且允许这些后代存活数月,并传播到一个食物充足的新微环境中,从而完成循环。图 4说明雌雄同体个体最初传播到新的食物环境时,与雄性的异交显著[53]。这使后代的数量可以从300增加到1000,大大增加了秀丽隐杆线虫存活机率。

|
| 图4 C.elegans 蛔甙信息素运行的一般模式 Fig.4 General model for C. elegans ascaroside pheromone function |
从圆圈底部楔形开始,代表高食物含量和低量信息素的微环境,沿顺时针方向表示食物水平依次降低,信息素水平依次增加。圆形表面的图案表示食物,颜色深浅表示信息素[36]
6 线虫蛔甙对其他物种行为的影响寄生线虫真菌中有一种称为捕食线虫真菌(nematode-trapping fungi),是一种生活在土壤中的线虫专性寄生天敌[54]。捕食线虫真菌在进化中形成了专用于寄生的捕食器。2012年发现,线虫分泌在土壤中的蛔甙诱导捕食真菌产生捕食器[55]。此现象是以微生物天敌基于线虫信息传递信号使寄生与被寄生产生协同进化的一个例证,证明了三营养级水平的信息物质也参与进化进程。
7 讨论近年来蛔甙的结构与鉴定、组成和功能、生物合成与代谢调控、化学感受信号途径以及信息调节模式都有了深入快速的研究,为线虫化学生态学的学科发展提供了借鉴。今后需要继续鉴定和深入研究蛔甙对寄生线虫的滞育发育诱导机制。同时,在分子生物学方面,可以继续深入完善蛔甙的化感通路,包括新的化感受体、化感器、神经元及其功能等。而蛔甙在线虫及其与捕食真菌协同进化中的作用更显示出蛔甙在多物种互作和进化方面研究的巨大潜力。
在应用方面,蛔甙是寄生线虫种群发展的重要调节器,了解寄生线虫的化学信号能够帮助人们挖掘控制这些物种的工具。识别蛔甙的物种特异性可能使该信息素能够干扰并控制寄生线虫的繁殖和生存,对于研发控制动植物寄生线虫危害和传播新型药剂提供了新思路。有研究表明,昆虫寄生线虫嗜菌异小杆线虫利用小分子信息素来控制感染势态的发展。该信息素与秀丽隐杆线虫的多尔幼虫信息素蛔甙在结构上类似,但存在种间高度特异性[56]。蛔甙对昆虫寄生线虫滞育态的维持功能有利于提高昆虫寄生线虫在农业害虫防控中的应用,即解决昆虫寄生线虫的长途运输和季节性进入繁殖周期的问题。例如将蛔甙维持昆虫寄生线虫的多尔状态进行长途运输和长期保存,在需要发挥作用的时候进行释放,并使其进入繁殖周期。而植物寄生线虫多尔形成和雌雄交配行为对于它们的种群密度和扩散传播等危害程度影响也很大[55],急需绿色、安全的信息素防控技术,对蛔甙在其中起的关键调控作用进行研究将给予新型技术研发的希望。因此,围绕蛔甙进行寄生线虫种群调控的意义很大。
| [1] | Poinar G O Jr. The natural History of Nematodes. Englewood Cliffs: Prentice-Hall, 1983. |
| [2] | Huettel R N. Chemical communicators in nematodes. Journal of Nematology, 1986, 18: 3-8. |
| [3] | Perry R N. Chemoreception in plant parasitic nematodes. Annual Review of Phytopathology, 1996, 34(1): 181-199. |
| [4] | O'halloran D M, Burnell A M. An investigation of chemotaxis in the insect parasitic nematode Heterorhabditis bacteriophora. Parasitology, 2003, 127(4): 375-385. |
| [5] | Bargmann C I. Chemosensation in C. elegans//The C. elegans Research Community. WormBook, 2006: 1-29. |
| [6] | Jeong P Y, Jung M, Yim Y H, Kim H, Park M, Hong E, Lee W, Kim Y H, Kim K, Paik Y K. Chemical structure and biological activity of the Caenorhabditis elegans dauer-inducing pheromone. Nature, 2005, 433(7025): 541-545. |
| [7] | von Reuss S H, Bose N, Srinivasan J, Yim J J, Judkins J C, Sternberg P W, Schroeder F C. Comparative Metabolomics Reveals Biogenesis of Ascarosides, a Modular Library of Small-Molecule Signals in C. elegans. Journal of the American Chemical Society, 2012, 134(3): 1817-1824. |
| [8] | Ludewig A H, Schroeder F C. Ascaroside signaling in C. elegans. WormBook: the online review of C. elegans biology, 2013: 1. |
| [9] | Choe A, von Reuss S H, Kogan D, Gasser R B, Platzer E G, Schroeder F C, Sternberg P W. Ascaroside signaling is widely conserved among nematodes. Current Biology, 2012, 22(9): 772-780. |
| [10] | Hu P J. Dauer. WormBook, 2007: 1-19. http://www.wormbook.org/chapters/www_dauer/dauer.html. |
| [11] | Cassada R C, Russell R L. The dauerlarva, a post-embryonic developmental variant of the nematode Caenorhabditis elegans. Developmental Biology, 1975, 46(2): 326-342. |
| [12] | Klass M, Hirsh D. Non-ageing developmental variant of Caenorhabditis elegans. Nature, 1976, 260(5551): 523-525. |
| [13] | Golden J W, Riddle D L. A pheromone influences larval development in the nematode Caenorhabditis elegans. Science, 1982, 218(4572): 578-580. |
| [14] | Golden J W, Riddle D L. A Caenorhabditis elegans dauer-inducing pheromone and an antagonistic component of the food supply. Journal of Chemical Ecology, 1984, 10(8): 1265-1280. |
| [15] | Butcher R A, Fujita M, Schroeder F C, Clardy J. Small-molecule pheromones that control dauer development in Caenorhabditis elegans. Nature Chemical Biology, 2007, 3(7): 420-422. |
| [16] | Baiga T J, Guo H B, Xing Y L, O'Doherty G A, Dillin A, Austin M B, Noel J P, La Clair J J. Metabolite induction of Caenorhabditis elegans dauer larvae arises via transport in the pharynx. ACS Chemical Biology, 2008, 3(5): 294-304. |
| [17] | Gallo M, Riddle D L. Effects of a Caenorhabditis elegans dauer pheromone ascaroside on physiology and signal transduction pathways. Journal of Chemical Ecology, 2009, 35(2): 272-279. |
| [18] | Golden J W, Riddle D L. The Caenorhabditis elegans dauer larva: Developmental effects of pheromone, food, and temperature. Developmental Biology, 1984, 102(2): 368-378. |
| [19] | Butcher R A, Ragains J R, Li W, Ruvkun G, Clardy J, Mak H Y. Biosynthesis of the Caenorhabditis elegans dauer pheromone. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(6): 1875-1879. |
| [20] | Kaplan F, Srinivasan J, Mahanti P, Ajredini R, Durak O, Nimalendran R, Sternberg P W, Teal P E A, Schroeder F C, Edison A S, Alborn H T. Ascaroside expression in Caenorhabditis elegans is strongly dependent on diet and developmental stage. PLoS ONE, 2011, 6(3): e17804. |
| [21] | Butcher R A, Ragains J R, Kim E, Clardy J. A potent dauer pheromone component in Caenorhabditis elegans that acts synergistically with other components. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(38): 14288-14292. |
| [22] | Srinivasan J, Kaplan F, Ajredini R, Zachariah C, Alborn HT, Teal P E, Malik R U, Edison A S, Sternberg P W, Schroeder F C. A blend of small molecules regulates both mating and development in Caenorhabditis elegans. Nature, 2008, 454(7208): 1115-1118. |
| [23] | Pungaliya C, Srinivasan J, Fox B W, Malik R U, Ludewig A H, Sternberg P W, Schroeder F C. A shortcut to identifying small molecule signals that regulate behavior and development in Caenorhabditis elegans. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(19): 7708-7713. |
| [24] | Butcher R A, Ragains J R, Clardy J. An indole-containing dauer pheromone component with unusual dauer inhibitory activity at higher concentrations. Organic Letters, 2009, 11(14): 3100-3103. |
| [25] | Izrayelit Y, Srinivasan J, Campbell S L, Jo Y, von Reuss S H, Genoff M C, Sternberg P W, Schroeder F C. Targeted metabolomics reveals a male pheromone and sex-specific ascaroside biosynthesis in Caenorhabditis elegans. ACS Chemical Biology, 2012, 7(8): 1321-1325. |
| [26] | Srinivasan J, von Reuss S H, Bose N, Zaslaver A, Mahanti P, Ho M C, O'Doherty O G, Edison A S, Sternberg P W, Schroeder F C. A modular library of small molecule signals regulates social behaviors in Caenorhabditis elegans. PLoS Biology, 2012, 10(1): e1001237. |
| [27] | Yamada K, Hirotsu T, Matsuki M, Butcher R A, Tomioka M, Ishihara T, Clardy J, Kunitomo H, Iino Y. Olfactory plasticity is regulated by pheromonal signaling in Caenorhabditis elegans. Science, 2010, 329(5999): 1647-1650. |
| [28] | Macosko E Z, Pokala N, Feinberg E H, Chalasani S H, Butcher R A, Clardy J, Bargmann C I. A hub-and-spoke circuit drives pheromone attraction and social behaviour in C. elegans. Nature, 2009, 458(7242): 1171-1175. |
| [29] | Fouquey C, Polonsky J, Lederer E. Chemical structure of ascarylic alcohol isolated fromParascaris equorum. Bulletin De La Société De Chimie Biologique, 1957, 39(1): 101-132. |
| [30] | Jezyk P F, Fairbairn D. Ascarosides and ascaroside esters in Ascaris lumbricoides (Nematoda). Comparative Biochemistry and Physiology, 1967, 23(3): 691-705. |
| [31] | Tarr G E, Fairbairn D. Conversion of ascaroside esters to free ascarosides in fertilized eggs of Ascaris suum (Nematoda). The Journal of Parasitology, 1973, 59(3): 428-433. |
| [32] | Bartley J P, Bennett E A, Darben P A. Structure of the ascarosides from Ascaris suum. Journal of Natural Products, 1996, 59(10): 921-926. |
| [33] | Lapage G. Veterinary Parasitology. Edinburgh: Oliver and Boyd, 1956. |
| [34] | Barrett J. Studies on the induction of permeability inAscaris lumbricoides eggs. Parasitology, 1976, 73(1): 109-121. |
| [35] | Fagerholm H P, Nansen P, Roepstorff A, Frandsen F, Eriksen L. Differentiation of cuticular structures during the growth of the third-stage larva of Ascaris suum (Nematoda, Ascaridoidea) after emerging from the egg. Journal of Parasitology, 2000, 86(3): 421-427. |
| [36] | Edison A S. Caenorhabditis elegans Pheromones Regulate Multiple Complex Behaviors. Current Opinion in Neurobiology, 2009, 19(4): 378-388. |
| [37] | Holterman M, van der Wurff A, van den Elsen S, van Megen H, Bongers T, Holovachov O, Bakker J, Helder J. Phylum-wide analysis of SSU rDNA reveals deep phylogenetic relationships among nematodes and accelerated evolution toward crown clades. Molecular Biology and Evolution, 2006, 23(9): 1792-1800. |
| [38] | Ye W M, Torres-Barragan A, Cardoza Y J. Oscheius carolinensis n. sp. (Nematoda: Rhabditidae), a potential entomopathogenic nematode from vermicompost. Nematology, 2010, 12(1): 121-135. |
| [39] | Wang J, Kim S K. Global analysis of dauer gene expression in Caenorhabditis elegans. Development, 2003, 130(8): 1621-1634. |
| [40] | Berg J M, Tymoczko J L, Stryer L. Biochemistry. 5th ed. New York: WH Freeman and Co., 2002. |
| [41] | Burnell A M, Houthoofd K, O'Hanlon K, Vanfleteren J R. Alternate metabolism during the dauer stage of the nematode Caenorhabditis elegans. Experimental Gerontology, 2005, 40(11): 850-856. |
| [42] | Troemel E R. Chemosensory signaling in C. elegans. Bioessays, 1999, 21(12): 1011-1020. |
| [43] | Inglis P N, Ou G S, Leroux M R, Scholey J M. The sensory cilia of Caenorhabditis elegans. WormBook, 2007, 1-22. |
| [44] | de Bono M, Bargmann C I. Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans. Cell, 1998, 94(5): 679-689. |
| [45] | Ebrahimi F A W, Chess A. Olfactory G proteins: simple and complex signal transduction. Current Biology, 1998, 8(12): R431-R433. |
| [46] | O'Halloran D M, Fitzpatrick D A, Burnell A M. The chemosensory system of Caenorhabditis elegans and other nematodes//Schoonhoven L M, Van Loon J J A, Dicke M. Chemical Ecology: From Gene to Ecosystem. New York: Springer Press, 2006, 71-88. |
| [47] | Kim K, Sato K, Shibuya M, Zeiger D M, Butcher R A, Ragains J R, Clardy J, Touhara K, Sengupta P. Two chemoreceptors mediate developmental effects of dauer pheromone in C. elegans. Science, 2009, 326(5955): 994-998. |
| [48] | McGrath P T, Xu Y F, Ailion M, Garrison J L, Butcher R A, Bargmann C I. Parallel evolution of domesticated Caenorhabditis species targets pheromone receptor genes. Nature, 2011, 477(7364): 321-325. |
| [49] | Zwaal R R, Mendel J E, Sternberg P W, Plasterk R H A. Two neuronal G proteins are involved in chemosensation of the Caenorhabditis elegans dauer-inducing pheromone. Genetics, 1997, 145(3): 715-727. |
| [50] | Fielenbach N, Antebi A.C. elegans dauer formation and the molecular basis of plasticity. Genes & Development, 2008, 22(16): 2149-2165. |
| [51] | Beall M J, Pearce E J. Transforming growth factor-beta and insulin-like signalling pathways in parasitic helminths. International Journal for Parasitology, 2002, 32(4): 399-404. |
| [52] | Thomas J H, Robertson H M. The Caenorhabditis chemoreceptor gene families. BMC Biology, 2008, 6(1): 42. |
| [53] | Morran L T, Cappy B J, Anderson J L, Phillips P C. Sexual partners for the stressed: facultative outcrossing in the self-fertilizing nematode Caenorhabditis elegans. Evolution, 2009, 63(6): 1473-1482. |
| [54] | Barron G L. The nematode-destroying fungi. Guelph, Canada: Canadian Biological Publications Ltd., 1977. |
| [55] | Hsueh Y P, Mahanti P, Schroeder F C, Sternberg P W. Nematode-trapping fungi eavesdrop on nematode pheromones. Current Biology, 2013, 23(1): 83-86. |
| [56] | Noguez J H, Conner E S, Zhou Y, Ciche T A, Ragains J R, Butcher R A. A novel ascaroside controls the parasitic life cycle of the entomopathogenic nematode Heterorhabditis bacteriophora. ACS Chemical Biology, 2012, 7(6): 961-966. |