 2016, Vol. 36
2016, Vol. 36文章信息
- 高梅香, 刘冬, 张雪萍, 吴东辉
- GAO Meixiang, LIU Dong, ZHANG Xueping, WU Donghui
- 三江平原农田地表和地下土壤螨类丰富度与环境因子的空间关联性
- Spatial relationships between the abundance of aboveground and belowground soil mite communities, and environmental factors in a farmland on the Sanjiang Plain, China
- 生态学报, 2016, 36(6): 1782-1792
- Acta Ecologica Sinica, 2016, 36(6): 1782-1792
- http://dx.doi.org/10.5846/stxb201408271707
-
文章历史
- 收稿日期:2014-08-27
- 网络出版日期:2015-07-29






2. 黑龙江省高校地理环境与遥感监测重点实验室, 哈尔滨师范大学, 哈尔滨 150025;
3. 湿地生态与环境重点实验室, 中国科学院东北地理与农业生态研究所, 长春 130102
2. Key Laboratory of Remote Sensing Monitoring of Geographic Environment, College of Heilongjiang Province, Harbin Normal University, Harbin 150025, China;
3. Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China
空间格局与过程是土壤动物群落生态学研究的重要内容,也是揭示生态系统生物多样性维持机制的重要前提。很多环境条件可能对土壤动物空间分布格局产生影响,如植被类型、土壤结构、土壤含水量、土壤化学性质、土壤温湿度等[1, 2],如果不同物种对环境条件响应不同[3],那么这些物种就可能表现出不同的空间分布格局。因此,土壤动物与环境因子空间关联性研究,是揭示土壤动物空间格局调控机制的基础。然而,土壤有机体的空间分布在多种空间尺度上具复杂性和结构性特征[4, 5]。不同空间尺度上的调控因子和调控机制可能是不同的,因此在不同的空间尺度上土壤动物的空间格局可能表现不同。基于多尺度途径来分析土壤动物的空间格局[6]及其与环境因子的空间作用关系,可以为深入揭示土壤动物空间格局调控机制提供依据。地统计学中的空间分析方法,为揭示土壤动物空间格局及这种空间格局与环境因子的空间作用关系提供了有效的途径[7]。
土壤螨类是地下生态系统重要的生物组分,这类土壤动物体型较小、活动能力较弱[8, 9],广泛的活动于地表层(如农田地表和森林凋落物层)和土壤层,这种生活习性使得土壤螨类成为研究地表和地下格局与过程关系的理想实验对象。本研究在三江平原农田生态系统,分别以地表和地下螨类为研究对象,基于地统计空间分析的方法研究螨类空间分布格局,并揭示这种空间格局与环境因子空间格局的空间关联性,为地表-地下空间格局过程及生物多样性维持机制的研究奠定基础。
1 研究地区与研究方法 1.1 研究区概况三江平原位于黑龙江省东部,包括完达山以北的松花江、黑龙江和乌苏里江冲积形成的低平原和完达山以南乌苏里江支流与兴凯湖形成的冲-湖积平原。是中国最大的淡水沼泽湿地集中分布区,现在已经成为重要的粮食生产基地。试验在中国科学院三江平原沼泽湿地生态实验站内进行(133°31′ E,47°35′ N),研究区属温带大陆性季风气候区,年平均气温1.9 ℃,年均降水量约600 mm,降水集中在7—9月,地貌类型为三江平原沼泽发育最为普遍的碟形洼地;土壤类型包括草甸沼泽土。腐殖质沼泽土。泥炭沼泽土。潜育白浆土和草甸白浆土。
1.2 样地设置与野外调查方法实验样地设置在中国科学院三江平原沼泽湿地生态站内,选择由沼泽湿地开垦的具30a以上开发历史的旱地,该样地土壤类型为白浆土,实验当年种植作物为大豆。在该农田的中心区域设置一个50 m×50 m的样地,以5 m为间隔,将该样地等间距划分为100个5 m×5 m的小样方。以每个小样方的左下角网格线交叉点为中心,以15 cm为半径,用内径为7 cm的土钻随机采集4个10 cm深的土柱作为一个空间采样点调查地下土壤螨群落。然后在相同的空间范围内随机设置3个陷阱,内置醋和糖(诱捕)以及酒精(防腐)来获得地表土壤螨群落,将陷阱置于野外3d。同时在土壤动物样品的右侧使用土钻采集1个10cm深的土柱,带回室内风干处理待测土壤理化性质;同时在每个小样方内随机选择10株健康的大豆,从近地面量至株顶,取株高的平均值作为该小样方的大豆株高。野外调查于2011年8月份和10月份进行,地下土壤螨群落采用Tullgren干漏斗法进行分离,分离结束后显微镜下鉴定种类并计数。
1.3 数据处理分析 1.3.1 螨类物种鉴定和土壤含水量、pH值的测定土壤螨类鉴定到种[10, 11, 12, 13],成螨和若螨分开计数,仅将成螨个体用于后续分析[14, 15]。电位测定法获得土壤pH值,烘干法测定土壤含水量[16, 17],仅8月份有大豆株高数据。
1.3.2 半方差函数及理论模型拟合半方差函数(Semivariogram)是地统计学进行空间分析和揭示空间分异规律的基本数学工具,反映了不同距离的观测值之间的变化,所谓半方差函数就是两点间差值的方差的一半[18],即:
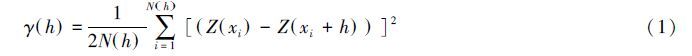
计算土壤动物群落丰富度及环境因子的变异函数值,依据决定系数最大、残差平方和较小的原则筛选拟合理论模型,应用模型参数[18]定量描述土壤动物丰富度及环境因子属性值的空间异质性特征。其中块金值(C0)代表了由实验误差和小于最小取样尺度(本实验为5m)引起的随机变异。块金值与基台值之比(C0/(C0+C))反映土壤动物及环境因子的空间依赖性,可表明属性值空间相关性的程度,也表示随机部分引起的空间异质性占总变异的比例。当块金值与基台值之比(C0/(C0+C)) < 25%时空间相关性强,其空间变异主要由结构性因素引起;当(C0/(C0+C))介于25%—75%之间时空间相关性中等,其空间变异由随机性因素和结构性因素共同决定;当(C0/(C0+C))>75%时空间相关性弱[19, 20],表明随机部分引起的空间变异起主要作用[21]。
1.3.3 普通克里格空间插值利用土壤动物丰富度、环境因子属性值和半方差函数的结构性,使用普通克里格插值(Ordinary Kriging,OK)对未采样点区域化变量(x0)进行最优无偏估值[18],对样地土壤螨类丰富度及环境因子属性值进行空间分布模拟,其实质是一个实行局部估计的加权平均值:

交叉方差函数(Crossvariogram)用于分析土壤动物空间格局与环境因子空间格局的空间关联性[22, 23]:
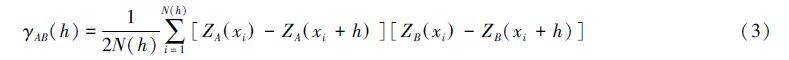
半方差函数和交叉方差函数在GS+9.0软件平台实现;普通克里格空间插值和简单Mantel检验分别在Arcgis 10.0和R软件(3.1.2)的“vegan"软件包实现。
2 结果 2.1 土壤螨群落空间异质性 2.1.1 土壤螨群落丰富度空间异质性特征8月份地表、地下和10月份地表、地下螨群落丰富度分别可以用指数、球状和线性、指数模型进行拟合。10月份地表螨群落丰富度的模型拟合度较低,表现为纯块金效应(图 1)。8月份地表、地下和10月份地下螨群落丰富度的(C0/(C0+C))分别为10.17、4.23和13.45%,表明螨群落丰富度具有强烈的空间自相关性,其空间分异主要由结构性因素决定,普通克里格插值表明这些螨群落丰富度空间斑块特征明显(图 2)。

|
| 图1 土壤螨群落丰富度半方差函数图 Fig.1 Semivariograms of soil mite communities |

|
| 图2 土壤螨群落丰富度空间分布格局 Fig.2 Spatial distribution patterns of soil mite communities |
8 月份地表螨群落中点肋甲螨属(Punctoribates sp.)、绥螨科(Sejidae sp.)和中气门亚目未定种2(Mesostigmata unidentified sp.2)的丰富度可用指数模型拟合,其(C0/(C0+C))均小于25%,表明其空间变异主要由结构性因素引起;坎盲走螨属(Kampinodromus sp.)和中气门亚目未定种1(Mesostigmata unidentified sp.1)的丰富度可用球状模型拟合,其(C0/(C0+C))分别小于25%和介于25%—75%之间,说明其空间变异分别由结构性因素及结构性因素和随机性因素共同决定。其他螨类物种丰富度的理论模型为线性模型,表现为纯块金效应。8月份地下螨群落中上洛甲螨属(Epilohmannia sp.)的丰富度可用高斯模型拟合,(C0/(C0+C))介于25%—75%之间,说明其空间变异由结构性因素和随机性因素共同决定。其余螨类丰富度可用球状模型拟合,(C0/(C0+C))均小于25%,表明空间变异主要由结构性因素引起(表 1)。
| 群落Community | 物种Species | 模型Model | 块金值Nuggetvariance (C0) | 基台值Structural variance sill(C0+C) | 块金值/ 基台值Proportion [C0/(C0+ C)]/% | 变程Range | 残差Residual sum of squares (RSS) | 决定系数Coefficient of determination (R2) |
| 地表螨群落Aboveground | 点肋甲螨属Punctoribates sp. | Exponential | 67.00 | 810.00 | 8.27 | 5.88 | 3265.00 | 0.19 |
| mite community | 小盾蛛甲螨属Suctobelbella sp. | Linear | 375.46 | 375.46 | - | 26.48 | 3395.00 | 0.77 |
| 盖甲螨属Tectocepheus sp. | Linear | 94.82 | 94.82 | - | 26.48 | 306.00 | 0.29 | |
| 网棱甲螨属Areozetes sp. | Linear | 80.37 | 80.37 | - | 26.48 | 65.10 | 0.08 | |
| 厉螨科Laelapidae sp. | Linear | 104.35 | 104.35 | - | 26.48 | 282.00 | 0.03 | |
| 绥螨科Sejidae sp. | Exponential | 7.70 | 39.71 | 19.39 | 7.02 | 15.60 | 0.21 | |
| 坎盲走螨属Kampinodromus sp. | Spherical | 0.80 | 8.99 | 8.90 | 8.77 | 3.32 | 0.23 | |
| 中气门亚目未定种1 Mesostigmata unidentified sp. 1 | Spherical | 3.10 | 6.20 | 49.99 | 16.00 | 0.72 | 0.66 | |
| 中气门亚目未定种2 Mesostigmata unidentified sp. 2 | Exponential | 5.50 | 45.02 | 12.22 | 6.87 | 9.71 | 0.39 | |
| 前气门亚目未定种Prostigmata unidentified sp. | Linear | 9.57 | 9.57 | - | 26.48 | 14.30 | 0.19 | |
| 地下螨群落Belowground | 点肋甲螨属Punctoribates sp. | Spherical | 0.01 | 21.39 | 4.68 | 7.75 | 14.40 | 0.12 |
| mite community | 小盾蛛甲螨属Suctobelbella sp. | Spherical | 0.10 | 37.63 | 0.27 | 5.98 | 22.20 | < 0.001 |
| 盖甲螨属Tectocepheus sp. | Spherical | 0.10 | 69.75 | 0.14 | 8.33 | 34.10 | 0.57 | |
| 网棱甲螨属Areozetes sp. | Spherical | 3.7 | 43.37 | 8.53 | 9.47 | 8.96 | 0.82 | |
| 华端三甲螨Acrotritiasinensis | Spherical | 0.04 | 0.29 | 12.65 | 5.98 | < 0.001 | < 0.001 | |
| 上洛甲螨属Epilohmannia sp. | Gaussian | 0.07 | 0.19 | 36.14 | 5.74 | < 0.001 | 0.61 | |
| 厉螨科Laelapidae sp. | Spherical | 0.11 | 20.42 | 0.54 | 10.03 | 3.96 | 0.79 | |
| 绥螨科Sejidae sp. | Spherical | 0.01 | 0.32 | 3.12 | 5.98 | 0.21 | < 0.001 | |
| 坎盲走螨属Kampinodromus sp. | Spherical | 0.001 | 0.45 | 22.42 | 6.77 | < 0.001 | 0.20 | |
| 中气门亚目未定种1 Mesostigmata unidentified sp. 1 | Spherical | 0.31 | 5.05 | 6.14 | 7.58 | 0.27 | 0.21 |
10月份所有地表螨类物种丰富度的理论模型均为线性模型。10月份地下螨群落中的点肋甲螨属(Punctoribates sp.)、网棱甲螨属(Areozetes sp.)和厉螨科(Laelapidae sp.)的理论模型均为线性模型,其余螨类物种丰富度均适合指数模型进行拟合。小盾蛛甲螨属(Suctobelbella sp.)的(C0/(C0+C))为38.53%,其空间变异由随机性因素和结构性因素共同决定。盖甲螨属(Tectocepheus sp.)、华端三甲螨(Acrotritiasinensis)、绥螨科(Sejidae sp.)和中气门亚目未定种1 (Mesostigmata unidentified sp. 1)的(C0/(C0+C))均小于25%,表明由结构性因素引起的变异占较大的比重(表 2)。
| 群落Community | 物种Species | 模型Model | 块金值Nuggetvariance (C0) | 基台值Structural variance sill(C0+C) | 块金值/基台值Proportion [C0/(C0+ C)]/% | 变程Range | 残差Residual sum of squares (RSS) | 决定系数Coefficient of determination (R2) |
| 地表螨群落 | M1 | Linear | 0 | 0 | - | 71.00 | 2229.00 | 0.41 |
| Aboveground | M2 | Linear | 0 | 0 | - | 71.00 | 1516.00 | 0.49 |
| mite community | M4 | Linear | 0 | 0 | - | 71.00 | 1077.00 | 0.65 |
| M7 | Linear | 0 | 0 | - | 71.00 | 2731.00 | 0.94 | |
| M10 | Linear | 0 | 0 | - | 71.00 | 146548.00 | <0.001 | |
| 地下螨群落 | M1 | Linear | 0 | 0 | - | 71.00 | 293.00 | 0.57 |
| Belowground | M2 | Exponential | 4.82 | 12.51 | 38.53 | 36.51 | 1.09 | 0.82 |
| mite community | M3 | Exponential | 2.48 | 16.78 | 14.78 | 12.57 | 1.77 | 0.81 |
| M4 | Linear | 5.31 | 5.31 | - | 26.48 | 1.71 | 0.17 | |
| M5 | Exponential | 0 | 0.01 | 0.18 | 10.05 | <0.001 | 0.55 | |
| M7 | Linear | 4.67 | 4.67 | - | 26.48 | 0.51 | 0.01 | |
| M8 | Exponential | 0.13 | 0.84 | 14.86 | 13.50 | <0.001 | 0.86 | |
| M10 | Exponential | 0.02 | 0.50 | 3.62 | 8.52 | < 0.001 | 0.59 | |
| M1: 点肋甲螨属 (Punctoribates sp.);M2: 小盾蛛甲螨属(Suctobelbella sp.);M3: 盖甲螨属(Tectocepheus sp.);M4: 网棱甲螨属(Areozetes sp.);M5: 华端三甲螨(Acrotritiasinensis);M7: 厉螨科(Laelapidae sp.);M8: 绥螨科(Sejidae sp.);M10: 中气门亚目未定种1 (Mesostigmata unidentified sp.1). | ||||||||
8月份土壤含水量、土壤pH值和大豆株高分别可用球状、高斯和球状模型拟合;10月份土壤含水量和土壤pH值分别可用指数和球状模型拟合;这些环境因子的(C0/(C0+C))均小于25%,表明其空间变异主要由结构性因素引起(表 3)。普通克里格插值进一步表明这些环境因子在8月份和10月份均具明显的空间斑块性特征。
| 月份Month | 因子Factor | 模型Model | 块金值Nugget variance (C0) | 基台值Structural variance sill(C0+C) | 块金值/ 基台值Proportion [C0/(C0+ C)%] | 变程Range | 残差Residual sum of squares (RSS) | 决定系数Coefficient of determination (R2) |
| 8 | Soil water content | Spherical | 2.33 | 12.65 | 7.30 | 81.60 | 4.00 | 0.04 |
| Soil pH | Gaussian | 0.0042 | 0.04 | 5.26 | 88.60 | <0.001 | 0.03 | |
| Average height of soybean | Spherical | 12.99 | 35.59 | 22.88 | 63.50 | 0.05 | 1.00 | |
| 10 | Soil water content | Exponential | 0.01 | 4.12 | 0.81 | 9.48 | 0.58 | 0.25 |
| Soil pH | Spherical | 0 | 0.03 | 1.55 | 6.98 | <0.001 | 0.08 |
8月份地表螨群落丰富度与土壤含水量和土壤pH值分别在5.98—26.48 m为负的空间相关,与大豆株高在5.98—21.13 m为正的空间相关,大于21.13 m之后为负的空间相关。8月份地下螨群落丰富度与土壤含水量在5.98—26.48 m具正的空间相关;和土壤pH值、大豆株高在5.98—26.48 m均为负的空间相关(图 3)。10月份地表螨群落丰富度与土壤含水量在5.98—26.48 m为空间负相关,与土壤pH值在5.98—26.48 m为空间正相关。10月份地下螨群落丰富度与土壤含水量和土壤pH值均为多尺度的空间负相关。

|
| 图3 8月份地表和地下土壤螨群落丰富度与土壤含水量、pH值和大豆株高的空间关联性 Fig.3 The spatial relationships between aboveground and belowground soil mite abundances and soil water content,soil pH and soybean height in August |
8月份土壤含水量和地表点肋甲螨属(Punctoribates sp.)、盖甲螨属(Tectocepheus sp.)、网棱甲螨属(Areozetes sp.)、绥螨科(Sejidae sp.)、坎盲走螨属(Kampinodromus sp.)和中气门亚目未定种2(Mesostigmata unidentified sp.2)在5.98—26.48 m均表现为空间负相关;和中气门亚目未定种1(Mesostigmata unidentified sp.1)在5.98—26.48m表现为空间正相关,和小盾蛛甲螨属(Suctobelbella sp.)在11.51 m为空间负相关.其余尺度为空间正相关;和厉螨科(Laelapidae sp.)在5.98—11.51 m为空间负相关,大于11.51m之后为空间正相关。土壤pH值和点肋甲螨属(Punctoribates sp.)、小盾蛛甲螨属(Suctobelbella sp.)、盖甲螨属(Tectocepheus sp.)、厉螨科(Laelapidae sp.)、绥螨科(Sejidae sp.)和中气门亚目未定种2(Mesostigmata unidentified sp.2)在5.98—26.48 m均为空间负相关;和坎盲走螨属(Kampinodromus sp.)、前气门亚目未定种(Prostigmata unidentified sp.)在5.98—26.48 m为空间正相关;和网棱甲螨属(Areozetes sp.)在21.13 m为空间负相关,在其他尺度为空间正相关;和中气门亚目未定种1(Mesostigmata unidentified sp.1)在5.98 m、21.13 m为空间负相关,在其他尺度为空间正相关。大豆株高和网棱甲螨属(Areozetes sp.)、中气门亚目未定种2(Mesostigmata unidentified sp.2),前气门亚目未定种(Prostigmata unidentified sp.)在5.98—26.48 m为空间负相关;和小盾蛛甲螨属(Suctobelbella sp.)、厉螨科(Laelapidae sp.)、绥螨科(Sejidae sp.)、坎盲走螨属(Kampinodromus sp.)、中气门亚目未定种1(Mesostigmata unidentified sp.1)在5.98—26.48 m为空间正相关;和点肋甲螨属(Punctoribates sp.)在26.48 m为空间负相关,在其他尺度均为空间正相关;和盖甲螨属(Tectocepheus sp.)在5.98 m为空间正相关,大于5.98m之后为空间负相关(表 4)。8月份地下和10月份地表、地下螨类物种丰富度和环境因子均在多种尺度上表现为复杂的正的或负的空间关联性。
| 物种 Species | |||||||||||||||
| 土壤含水量Soil water content | 土壤pH值Soil pH | 大豆株高Average height of soybean | |||||||||||||
| 5.98m | 11.51m | 16.43m | 21.13m | 26.48m | 5.98m | 11.51m | 16.43m | 21.13m | 26.48m | 5.98m | 11.51m | 16.43m | 21.13m | 26.48m | |
| M1 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 正 | 负 |
| M2 | 正 | 负 | 正 | 正 | 正 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 正 | 正 |
| M3 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 正 | 负 | 负 | 负 | 负 |
| M4 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 负 | 正 | 负 | 负 | 负 | 负 | 负 |
| M7 | 负 | 负 | 正 | 正 | 正 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 正 | 正 |
| M8 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 正 | 正 |
| M9 | 负 | 负 | 负 | 负 | 负 | 正 | 正 | 正 | 正 | 正 | 正 | 正 | 正 | 正 | 正 |
| M10 | 正 | 正 | 正 | 正 | 正 | 负 | 正 | 正 | 负 | 正 | 正 | 正 | 正 | 正 | 正 |
| M11 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 | 负 |
| M13 | 负 | 正 | 负 | 正 | 负 | 正 | 正 | 正 | 正 | 正 | 负 | 负 | 负 | 负 | 负 |
| M1: 点肋甲螨属 Punctoribates sp.; M2: 小盾蛛甲螨属Suctobelbella sp.; M3: 盖甲螨属Tectocepheus sp.; M4: 网棱甲螨属Areozetes sp.; M7: 厉螨科Laelapidae sp.; M8: 绥螨科Sejidae sp.; M9: 坎盲走螨属Kampinodromus sp.; M10: 中气门亚目未定种1 Mesostigmata unidentified sp.1; M11: 中气门亚目未定种2 Mesostigmata unidentified sp.2; M13: 前气门亚目未定种Prostigmata unidentified sp.; 正: 该种土壤螨类物种丰富度和环境因子在该种尺度为空间正相关;负: 该种土壤螨类物种丰富度和环境因子在该种尺度为空间负相关 | |||||||||||||||
简单Mantel检验结果表明,仅8月份地表中气门亚目未定种1(Mesostigmata unidentified sp.1)和大豆株高具有显著的正的空间关联性(r=0.29,P=0.04),其他螨类物种丰富度和环境因子的空间关联性均不显著。
3 讨论土壤动物的水平分布是复杂的,且在不同的空间尺度具结构性特征[24, 25]。地下生态系统生态学过程在不同的空间尺度可能会表现出不同的空间格局特征,空间分析过程中多尺度研究是揭示这种结构性特征的重要方法和手段[26]。在土壤动物群落空间异质性研究过程中引入地统计学方法,可以定量得出土壤动物多样性的变异程度和变化范围,合理的描述土壤动物多样性在空间分布上的随机性和结构性特征,表征多样性在各种尺度上的变异规律,为地下生态系统格局与过程及生物多样性维持机制研究奠定基础。
半方差函数分析结果表明8月份地表、地下和10月份地下土壤螨群落及这些群落中大部分螨类物种具有显著的空间自相关性[27],其空间格局分别可以用指数、球状和高斯模型进行拟合,并在5—40 m空间尺度内形成集群[27],表现出一种明显的空间结构。相关研究也表明土壤动物在热点区内(hot spots)高度集群,并且在几十米范围内形成空间结构[28]。地表和地下螨类的这种集群性格局是土壤有机体最常见的一种空间结构[7, 29, 30]。块金值和基台值的比值(C0/(C0+C))表明,8月份地下和10月份地下土壤螨群落及这些群落大部分螨类物种及8月份地表近一半螨类物种的空间变异,是结构性因素或结构性因素和随机性因素共同调控的结果,说明在揭示螨群落物种共存机制的过程中,一些确定性过程和非确定性过程均不容忽视。基于生态位理论的环境筛选和种间竞争(确定性过程)及基于中性理论的扩散限制(非确定性过程),被认为是调控这种空间异质性格局的重要机制[14, 31, 32, 33]。相关研究发现,基于生物间相互作用的种间竞争对该农田土壤螨群落空间格局的调控作用并不明显[34],说明环境筛选和扩散限制可能在该小尺度空间起主要调控作用[35],相关研究还有待于进一步深入开展。而10月份地表螨群落、8月份地表近一半螨类物种和10月份地表所有螨类物种的丰富度均表现为明显的纯块金效应,即存在由采样误差、微尺度空间变异等因素带来的块金效应,该结果强调小于5 m的空间过程不容忽视,为了揭示10月份地表螨群落及这些螨类物种的空间异质性格局,需要在现有的空间尺度内,缩小空间粒度并加大采样密度进行研究。
和土壤螨类生存密切相关的环境因子也表现为明显的空间结构性特征。8月份土壤含水量、土壤pH值和大豆株高分别在约85、90和65 m的空间尺度内形成集群格局,10月份土壤含水量和土壤pH值分别在10和7 m的空间尺度内集群。对于相同环境因子来说,8月份形成的集群斑块明显大于10月份形成的集群斑块。但是这些环境因子的空间异质性,与生存其中的地表和地下螨群落及螨类物种空间异质性并不完全一致。块金值和基台值的比值(C0/(C0+C))表明这些环境因子的空间异质性均表现为明显的结构性特征,土壤含水量。土壤pH值和大豆株高的空间分异主要受到结构性因素的调控。总体看来,无论是土壤螨群落、螨类物种还是土壤含水量、土壤pH值、大豆株高,其空间异质性在8月份和10月份表现规律均不相同,均存在着一定的时间动态性差异。
交叉方差函数分析的结果表明,8月份地表、地下和10月份地表、地下土壤螨群落及不同螨类在多种尺度上和环境因子表现出复杂的正的或负的空间关联性。但Mantel检验仅发现8月份地表中气门亚目未定种1(Mesostigmata unidentified sp.1)和大豆株高存在明显的正的空间关联性。表明土壤螨群落和其他螨类物种与环境因子存在伪的(spurious)空间关联性,这种空间关联性可能由简单的空间格局形成,并不是真正的空间相互关联性(not a true correlation)[7]。非生物因子常常被认为是影响土壤动物分布的重要因子,如土壤水分含量被认为是湿地生境影响螨类空间分布格局的非常重要的因子[36]。而本研究未发现土壤螨类丰富度和土壤含水量、土壤pH值和大豆株高的这种显著的空间关联性,Hasegawa[37]也未发现土壤螨群落丰富度和总有机质(Total organic matter)的明显关联,Gutiérrez-López M等[7]基于地统计分析也没有发现土壤螨群落及不同螨类物种丰富度和环境因子的显著空间关联性。当然,我们的研究结果不能简单的否定环境因子与土壤螨类的空间关联性,这种结果恰恰说明在基于多尺度条件考察土壤螨类与环境因子空间关联性时,存在着更大的复杂性和不确定性。另外,本研究所测量的环境因子变量较少(土壤含水量、土壤pH值和大豆株高),这三种环境因子难以代表全部环境因子与土壤螨群落间复杂的空间作用关系,需要在将来考虑更多重要的环境因子、在更综合的层次上进行分析探讨。
4 结论本研究表明三江平原农田生态系统,夏季地表。地下和秋季地下土壤螨群落及这些群落内大部分螨类物种存在明显的空间异质性格局,表现为一定的时间动态变化特征,且结构性因素或结构性因素和随机性因素对这种空间分异起主要调控作用,但土壤螨群落及大部分螨类物种丰富度和环境因子并未表现出明显的多尺度空间关联性。本研究仅能表明50m×50m尺度下的地表和地下土壤螨类空间格局及这种格局与环境因子的空间关联性,其他空间尺度的相关研究有待于进一步开展。
致谢: 感谢常亮、张兵、宋理洪、沙迪、张丽梅在野外调查和样品分离过程中的帮助,感谢李荣对写作的帮助。
| [1] | Sousa J P,da Gama M M,Ferreira CS. Effects of replacing oak-woods by eucalyptus on edaphic Collembola communities:does the size and type of plantation matter.ActaEntomologicaIbérica e Macaronesica,2003,1: 1-10. |
| [2] | Coulson S J,Hodkinson I D,Webb N R. Microscale distribution patterns in high Arctic soil microarthropod communities: the influence of plant species within the vegetation mosaic. Ecography,2003,26(6): 801-809. |
| [3] | Jiménez J J,Rossi J P. Spatial dissociation between two endogeic earthworms in the Colombian "Llanos". European Journal of Soil Biology,2006,42(S1):S218-S224. |
| [4] | Rossi J P. Short-range structures in earthworm spatial distribution: The 7th international symposium on earthworm ecology · Cardiff · Wales · 2002. Pedobiologia,2003,47(5/6): 582-587. |
| [5] | Rossi J P. The spatiotemporal pattern of a tropical earthworm species assemblage and its relationship with soil structure: The 7th international symposium on earthworm ecology · Cardiff · Wales · 2002. Pedobiologia,2003,47(5/6): 497-503. |
| [6] | Birkhofer K,Henschel J R,Scheu S. Spatial-pattern analysis in a territorial spider: evidence for multi-scaleeffects. Ecography,2006,29(5): 641-648. |
| [7] | Gutiérrez-López M,Jesús J B,Trigo D,Fernández R,Novo M,Díaz-Cosín D J. Relationships among spatial distribution of soil microarthropods,earthworm species and soil properties. Pedobiologia,2010,53(6): 381-389. |
| [8] | Lindo Z,Winchester N N. Spatial and environmental factors contributing to patterns in arboreal and terrestrial oribatid mite diversity across spatial scales. Oecologia,2009,160(4): 817-825. |
| [9] | Ojala R,Huhta V. Dispersal of microarthropods in forest soil. Pedobiologia,2001,45(5): 443-450. |
| [10] | Krantz G W,Walter D E. AManual of Acarology. 3rded.Lubbock,Texas: Texas Tech University Press,2009. |
| [11] | Walter D E,Proctor H C. Mites in Soil:an Interactive Key to Mites and Other Soil Microarthropods.Australia: CSIRO Publishing,2001. |
| [12] | 尹文英,胡圣豪,沈韫芬,宁应之,孙希达,吴纪华,诸葛燕,张云美,王敏,陈建英,徐成钢,梁彦龄,王洪铸,杨潼,陈德牛,张国庆,宋大祥,陈军,梁来荣,胡成业,王慧芙,张崇州,匡溥人,陈国孝,赵立军,谢荣栋,张骏,刘宪伟,韩美贞,毕道英,肖宁年,杨大荣. 中国土壤动物检索图鉴. 北京: 科学出版社,1998: 163-242. |
| [13] | Balogh J,Balogh P. The OribatidMites Genera of the World (vol.1,2).Budapest,Hungary: The Hungarian National Museum Press,1992. |
| [14] | Ingimarsdóttir M,Caruso T,Ripa J,Magnúsdóttir Ó B,Migliorini M,Hedlund K. Primary assembly of soil communities: disentangling the effect of dispersal and local environment. Oecologia,2012,170(3): 745-754. |
| [15] | Minor M A. Spatial patterns and local diversity in soil oribatid mites (Acari: Oribatida) in three pine plantation forests. European Journal of Soil Biology,2011,47(2): 122-128. |
| [16] | 劳家柽. 土壤农化分析手册. 北京: 农业出版社,1988. |
| [17] | Pansu M,Gautheyrou J. Handbook of Soil Analysis: Mineralogical,Organic and Inorganic Methods. Berlin,Heidelberg: Springer,2006.. |
| [18] | 王政权. 地统计学及在生态学中的应用. 北京: 科学出版社,1999. |
| [19] | Kravchenko A N. Influence of spatial structure on accuracy of interpolation methods. Soil Science Society of America Journal,2003,67(5): 1564-1571. |
| [20] | CambardellaC A,Moorman T B,Parkin T B,Karlen D L,Novak J M,Turco R F,Konopka A E. Field-scale variability of soil properties in central Iowa soils. Soil Science Society of America Journal,1994,58(5): 1501-1511. |
| [21] | 郭旭东,傅伯杰,马克明,陈利顶,杨福林. 基于 GIS 和地统计学的土壤养分空间变异特征研究——以河北省遵化市为例. 应用生态学报,2000,11(4): 557-563. |
| [22] | Rossi J P,Lavelle P,Tondoh J E. Statistical tool for soil biology X. Geostatistical analysis. European Journal of Soil Biology,1995,31(4): 173-181. |
| [23] | 王正军,李典谟,商晗武,程家安. 地质统计学理论与方法及其在昆虫生态学中的应用. 昆虫知识,2002,39(6): 405-411. |
| [24] | Rossi J P. Clusters in earthworm spatial distribution: The 7th international symposium on earthworm ecology · Cardiff · Wales · 2002. Pedobiologia,2003,47(5/6): 490-496. |
| [25] | Lindo Z N,Winchester N. Scale dependent diversity patterns in arboreal and terrestrial oribatid mite (Acari: Oribatida) communities. Ecography,2008,31(1): 53-60. |
| [26] | Whittaker R J,Willis K J,Field R. Scale and species richness: towards a general,hierarchical theory of species diversity. Journal of Biogeography,2001,28(4): 453-470. |
| [27] | 高梅香,刘冬,吴东辉,张雪萍. 三江平原农田地表和地下土壤螨群落空间自相关性研究. 土壤学报,2014,51(6): 163-171. |
| [28] | Decans T,Rossi J P. Spatio-temporal structure of earthworm community and soil heterogeneity in a tropical pasture. Ecography,2001,24(6): 671-682. |
| [29] | 高梅香,孙新,吴东辉,张雪萍. 三江平原农田土壤跳虫多尺度空间自相关性. 生态学报,2014,34(17): 4980-4990. |
| [30] |
高梅香,何萍,刘冬,郭传伟,张雪萍,李景科. 温带落叶阔叶林土壤螨群落多尺度空间自相关性 |
| [31] | 高梅香,何萍,孙新,张雪萍,吴东辉. 环境筛选、扩散限制和生物间相互作用在温带落叶阔叶林土壤跳虫群落构建中的作用. 科学通报,2014,59(24): 2426-2438. |
| [32] | Caruso T,Taormina M,Migliorini M. Relative role of deterministic and stochastic determinants of soil animal community: a spatially explicit analysis of oribatid mites. Journal of Animal Ecology,2012,81(1): 214-221. |
| [33] | Caruso T,Trokhymets V,Bargagli R,Convey P. Biotic interactions as a structuring force in soil communities: evidence from the micro-arthropods of an Antarctic moss model system. Oecologia,2013,172(2): 495-503. |
| [34] | Lin L,Gao M X,Wu D H,Zhang X P,Wu H T. Co-occurrence patterns of above-ground and below-ground mite communities in farmland of Sanjiang Plain,Northeast China. Chinese Geographical Science,2014,24(3): 339-347. |
| [35] | Gao M X,He P,Liu D,Zhang X P,Wu D H. Relative roles of spatial factors,environmental filtering and biotic interactions in fine-scale structuring of a soil mite community. Soil Biology and Biochemistry,2014,79: 68-77. |
| [36] | Hernándeza P,Fernándeza R,Novoa M,Trigoa D,Cosín D J D. Geostatistical and multivariate analysis of the horizontal distribution of an earthworm community in El Molar (Madrid,Spain).Pedobiologia,2007,51(1): 13-21. |
| [37] | Hasegawa M. The relationship between the organic matter composition of a forest floor and the structure of a soil arthropod community.European Journal of Soil Biology,2001,37(4): 281-284. |