 2016, Vol. 36
2016, Vol. 36文章信息
- 王梓, 韩晓增, 张志明, 郝翔翔
- WANG Zi, HAN Xiaozeng, ZHANG Zhiming, HAO Xiangxiang.
- 中国东北黑土土壤剖面微生物群落碳源代谢特征
- Metabolic characteristics of the microbial community on the basis of carbon source in black soil profiles in northeastern China
- 生态学报[J]. 2016, 36(23): 7740-7748
- Acta Ecologica Sinica[J]. 2016, 36(23): 7740-7748
- http://dx.doi.org/10.5846/stxb201509281979
-
文章历史
- 收稿日期: 2015-09-28
- 修订日期: 2016-04-29





2. 中国科学院东北地理与农业生态研究所海伦农田生态系统国家野外科学观测研究站, 哈尔滨 150081
2. National Observation Station of Hailun Agroecology System, Northeast-Institute of Geography and Agroecology, Chinese Academy of Sciences, Harbin 150081, China
土壤生物被认为是地球关键元素生物, 是地球化学循环过程的引擎[1], 而土壤微生物由于数量巨大, 种类丰富成为其中非常重要的组成部分[2], 对土壤生态功能的发挥有极为重要的作用[3-5]。土壤是在地球表层母质或母岩上发育起来的自然体, 土体物质垂直分布的断面称为土壤剖面, 土壤剖面中的物质变化过程, 是植被根系-微生物-土壤非生命物质三者相互作用过程[6-7]。植物根系的分布会改变不同深度土层中微生物生存的物理环境, 同时根系代谢产物和死亡部分又能给微生物提供生命活动的能量和物质。剖面中微生物群落的活动控制C、N、P等重要元素的生物地球化学循环, 致使某些物质在土壤剖面中存在位置的再分配。所以随着时间的变化, 根系-微生物-土壤这个体系在相互作用中不断地向着某一个方向发育。
中国东北黑土是在冲积沉积物上发育的土壤, 土体深厚、土壤粘重, 形成界限分明的表层、亚表层、过渡层和母质层。剖面中不同层次间微生物群落结构和活性是不同的, 一些研究也证明了这一点, 在新疆阜康地区农田中表层土壤中放线菌门和变形菌门是群落中主要的类群, 二者相对丰度接近50%, 但是在亚表层中放线菌门相对丰度逐渐减少, 变形菌门相对丰度逐渐增加[8]。2012年研究人员对美国科罗拉多州戈登峡谷流域剖面微生物群落进行研究也表明表层土壤微生物群落组成与亚表层土壤微生物群落组成差异非常大[9]。这些研究结果表明土壤剖面中不同层次微生物群落组成差异很大, 从而推论剖面不同深度土层中微生物群落碳源代谢能力也会存在较大的差异, 然而截止到目前为止, 对黑土不同利用方式剖面不同深度土层中微生物碳源代谢特征的研究, 国内外还鲜有报道。
Biolog作为研究土壤微生物群落功能多样性的一种简单、快速方法[10-14], 已经对不同土壤类型[15]、不同植物根系活动下[16-17]的根际土壤微生物多样性和碳源代谢特征做了很多研究。本研究应用Biolog方法研究不同利用方式下黑土剖面不同深度土层中微生物群落碳源代谢特征的变化, 区分不同深度土层中微生物群落的主要代谢碳源类型和利用强度, 更深刻的理解微生物群落变化与土壤剖面物质转化的关系, 加深对土壤形成的认识, 具有重要的生态环境意义。
1 材料方法 1.1 研究区概况试验区位于中国科学院海伦农田生态系统国家野外科学观测研究站(47°26′N, 126°38′E), 地处黑龙江省中部, 属温带大陆性季风气候, 平均海拔234 m, 年均气温1.5℃, ≥10 ℃的有效积温2400-2500℃, 无霜期120-130 d, 作物生长期约120 d。全年降水量500-600 mm, 80%以上集中在5-9月[18]。
1.2 试验设计试验区位于500 m×150 m的区域内, 地势平坦, 土壤质地均一。土壤类型为第四纪黄土状亚黏土发育而成的中厚层黑土。于1985年设置以下3种土地利用方式:1)草地(GL), 1985年退耕休闲, 植被类型为草原化草甸植被, 主要植被有苔草(Carex)、问荆(Equisetum arvense L.)和狗尾草(Setaria viridis)等; 2)农田(Cropland,CL), 采用传统常规耕作方式, 不施肥, 种植制度为小麦-玉米-大豆轮作, 一年一熟制, 本研究当季种植作物为大豆(Soybean), 秋季收获后将作物地上部全部移除; 3)林地(Forestland,FL), 于1985年在耕地上直接种植红皮云杉(Picea koraiensis), 种植密度为2 m×2 m, 株高约4.5 m。
1.3 样品采集2013年6月每种利用方式挖取3个土壤剖面, 剖面深度为200 cm, 在表层采集0-10 cm和10-20 cm, 之后以每20 cm为1层, 共采集11层土壤样品。土壤样品装入封口袋内, 放入冰盒运回实验室, 4℃冰箱中保存, 以备分析。
1.4 试验方法土壤有机碳与全氮采用元素分析仪(EA3000, Euro Vector, Italy)测定; 碱解氮采用碱解扩散法测定; 土壤速效钾采用乙酸铵提取法-火焰光度计测定; 土壤pH采用电位法(水土比2.5:1)。
土壤可培养细菌、放线菌和真菌计数采用平板菌落计数法测定。细菌采用牛肉膏蛋白胨琼脂培养基(pH 7.0-7.2), 放线菌采用改良高氏1号培养基(pH 7.2-7.4), 真菌采用马丁(Mantin)培养基[19-21]。
Biolog EcoPlate接种和培养:称取相当于5 g烘干土重的鲜土加入到装有45 mL灭菌生理盐水(0.85%)的三角瓶中, 超声1 min, 然后在振荡器上振荡40 min混匀。土壤溶液稀释至10-3浓度梯度, 向ECO板中每孔加入150 μL土壤稀释液, 28℃保湿避光培养216 h。每隔24 h用Biolog自动读数仪测量590 nm (颜色和浊度)和750 nm (浊度)波长下的数值。
1.5 数据分析按照Classen[20]方法计算:
(1)平均颜色变化率(AWCD)
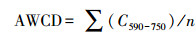
|
(2) Shannon指数H(评估物种丰富度)
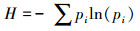
|
式中, C是反应孔吸光值, n是碳源数。pi为第i孔相对吸光度值与整个平板相对吸光度值总和的比率。
选择培养第6天的数据进行后续分析。
试验结果的方差分析及主成分分析采用SPSS 19.0进行。
2 结果 2.1 理化性质虽然地表植被不同, 但是SOC、TN和EN都随着深度增加而减少, 在0-10 cm土层中草地含量高于林地和农田。SOC、TN和EN在0-60 cm土层变化最大, 在60-200 cm土层变化减小。3个剖面表层pH都低于底层。
2.2 可培养微生物数量变化剖面不同层次可培养微生物数量变化结果见图 2。结果显示, 无论是真菌、细菌还是放线菌都是表层数量最多, 3种利用方式中60 cm以下土层的可培养微生物数量几乎相同。但是在0-20 cm土壤中无论真菌、细菌还是放线菌数量均为草地>农田>林地。

|
| 图 1 3种利用方式下剖面的土壤属性 Fig. 1 Soil properties of soil profile under three land use CL:农田Crop Land; GL:草地Grass Land; FL:林地Forest Land |

|
| 图 2 剖面中可培养微生物数量 Fig. 2 The number of cultivable microorganisms in three profiles |
从剖面不同层次平均颜色变化率(AWCD)结果(图 3)可以看到, 微生物群落的活性(AWCD高低)随剖面深度增加而减小。农田0-20 cm土层的AWCD值显著高于其他层次, 由于农田0-20 cm土壤每年都会翻耕, 并且作物根系主要集中在0-20 cm层次中, 所以在0-10cm和10-20cm土层中AWCD值相近。由于农田剖面中有犁底层产生, 这决定了耕层以下微生物群落的AWCD值较低。草地剖面中植被种类多根系密集, 所以0-10 cm和10-20 cm层表现出较高的AWCD值。恢复草地处理中由于大量杂草根系进入较深的土层中, 所以40-60 cm层中AWCD值仍然较高, 显著高于农田和林地相应层次。人工林地0-40 cm之间的3个土层的AWCD值比较接近, 但是显著高于深层次土层的AWCD值。

|
| 图 3 平均颜色变化率变化 Fig. 3 Average Well Color Development (AWCD) of microbial community in three profiles |
Shannon指数是应用最广泛的多样性指数, 是评价群落物种多样性的指数。3个处理中, 表层多样性指数显著高于底层(P < 0.05), 碳源利用数量结果也是如此(P < 0.05)(表 1)。在0-40 cm土层中草地剖面中的Shannon指数和碳源利用数量都是最高的, 说明草地植被会增加微生物群落的多样性。但是3个剖面在底层(180-200 cm)无论是多样性指数还是碳源利用数量是没有显著差异(表 1), 所以可以推测虽然不同类型的植被在同一块土地生长了近30年, 但是不同类型的植被并没有对底层的微生物群落碳源代谢多样性产生影响。
| 深度 Depth/cm | 香浓多样性指数 Shannon diversity index (H) | 碳源利用数 Number of carbon source utilized | ||||
| CL | GL | FL | CL | GL | FL | |
| 0-10 | 2.83±0.11Ba | 3.11±0.04Aa | 2.88±0.04Ba | 21.33±2.89Ba | 26.33±1.53Aa | 23.00±2ABa |
| 10-20 | 2.85±0.08Ba | 3.11±0.07Aa | 2.78±0.18Ba | 22.00±1Ba | 27.00±1.73Aa | 19.00±3.46Ba |
| 20-40 | 2.29±0.14Bbc | 2.81±0.02Ab | 2.85±0.07Aa | 20.67±1.15Ba | 20.00±1Ab | 21.33±0.58Aa |
| 40-60 | 2.35±0.07Bb | 2.63±0.1Ab | 2.37±0.06Bb | 23.00±2.65Aa | 15.33±2.08Bc | 12.67±0.58Bb |
| 80-100 | 2.03±0.15Bbc | 2.36±0.08Ac | 2.50±0.06Aab | 16.33±3.06Ab | 12.00±1Bd | 15.00±1ABb |
| 180-200 | 1.77±0.27Ac | 1.89±0.28Ad | 1.77±0.48Ac | 8.67±2.08Ac | 7.67±2.08Ae | 6.33±3.51Ac |
| CL:农田Crop Land; GL:草地Grass Land; FL:林地Forest Land;平均值±标准偏差, n=3;大写字母代表不同处理相同层次(P < 0.05), 小写字母代表相同处理不同层次(P < 0.05) | ||||||
Eco板中有31个碳源, 分为6类, 由于碳源含有生态有关的化合物, 所以更适合研究土壤微生物的功能多样性[11]。由于机械作业在农田耕层(0-20 cm)下会产生一层坚硬的犁底层, 会阻碍水分、肥料和植物根系进入土壤剖面下方, 从图 4中看到, 农田剖面中6类碳源的吸光值都是0-20 cm土层最高, 之后随深度增加而降低。糖类、氨基酸类、羧酸类和酚酸类在20 cm之后迅速下降, 这4类碳源在Eco板上31个碳源中是大多数。并且糖类、氨基酸类和羧酸类是土壤微生物群落利用最多的碳源, 这些碳源在20 cm土层以下利用程度迅速下降, 说明了人类农业活动对土壤剖面微生物群落的影响是很大的。草地和林地0-40 cm中各类碳源利用变化较大, 但是在40 cm以下碳源利用程度逐渐减小。

|
| 图 4 3个剖面不同土壤层次内微生物群落对6类碳源的利用强度 Fig. 4 Utilization intensity of six kind carbon sources in all layers of three profiles |
微生物群落对碳源利用情况反映了微生物群落组成的变化[21], 对碳源利用结果进行主成分分析。从图 5中看到在3个剖面中微生物群落从表层逐渐过渡到底层, 既反映了微生物群落碳源利用特征的变化, 也反映了微生物群落组成的变化。从0-100 cm土层中表现出植被根系对微生物群落都产生了影响, 导致微生物群落碳源利用特征产生分异, 而在180-200 cm土层中植被根系的影响几乎消失。

|
| 图 5 对3个剖面不同层次土壤微生物群落的主成分分析 Fig. 5 Principal component analysis of soil microbial community in all layers of three profiles |
虽然平板稀释法培养的微生物只是整个群落中的一小部分[22-23], 但是依然可以从某种程度上表现出微生物群落的变化趋势。在研究中可培养微生物数量随深度增加而减少, 与以前的研究相同[24]。前期的研究表明, 植被根系对SOC的影响可以达到100 cm以下[25], 但是对可培养微生物数量的影响只是在0-60 cm中(图 2)。在0-60 cm中草地植被显著提高了可培养微生物的数量, 林地的可培养微生物数量却是减少了, 这充分体现了植被对土壤微生物群落生存环境的影响。而且最近的研究表明, 不同的植被覆盖对微生物群落组成以及碳矿化速率都有非常大的影响[26]。从表层到深层的理化性质数据显示养分含量随逐渐减少, 容重逐渐增加[25], 说明越往深层土壤孔隙越少, 空气越少, 造成水分养分运移减少[27], 环境逐渐变得恶劣, 所以可培养微生物数量随深度减少甚至消失。
不仅可培养微生物总量随着深度变化而变化, 各类群微生物比例也随深度变化而变化(表 2)。农田由于每年翻耕, 所以0-10 cm和10-20 cm中的细菌、放线菌和真菌的数量和比例相近。由于20-40 cm是犁底层, 经过机械压实容重较大, 所以这一层中细菌比例减少, 细菌总量也是少于0-20cm表层土壤。可培养微生物在草地剖面所有土层中都被检测到, 在0-80 cm中细菌和放线菌的比例较大, 在80 cm以下细菌和放线菌比例急剧减少, 二者比例逐渐接近。说明根系作用的层次主要是0-80 cm。林地只在0-40 cm中3种微生物都被检测出, 但细菌与放线菌的比例显著小于同层次的农田和草地, 可能由于木本植被是多年生植被, 根系在土壤中很深, 近地表的根系并不是农作物与杂草那样柔软且具有大量根毛的根系, 所以不能刺激微生物生长; 林地中地表温度也要低于草地和农田; 其次林地地表有一层半分解的枯枝落叶层, 而且枯枝落叶分解主要依靠真菌。这些原因综合起来决定了林地中可培养微生物总量较少, 整个微生物群落中也是真菌比例较大。
| 深度 Depth/cm | 农田 Crop Land | 草地 Grass Land | 林地 Forest Land | 深度 Depth/cm | 农田 Crop Land | 草地 Grass Land | 林地 Forest Land |
| 0-10 | 462:22:1 | 652:82:1 | 185:14:1 | 10-20 | 466:22:1 | 321:44:1 | 191:11:1 |
| 20-40 | 315:22:1 | 1014:81:1 | 180:8:1 | 40-60 | 429:57:1 | 905:29:1 | - |
| 60-80 | 571:14:1 | 653:19:1 | - | 80-100 | - | 4:2:1 | - |
| 100-120 | - | 3:2:1 | - | 120-140 | - | 4:3:1 | - |
| 14-160 | - | 2:2:1 | - | 160-180 | - | 0.8:2:1 | - |
| 180-200 | - | 2:2:1 | - |
利用方式是影响土壤剖面土壤属性最主要的人为因素之一, 植被的转变导致进入土壤中凋落物种类和数量的变化, 造成凋落物分解后进入土壤剖面中的物质量和种类不同[28]。Biolog可研究土壤微生物群落对碳源利用能力的变化规律, 但无法对土壤微生物进行分类研究[29], 所以在本文研究有一定局限性。但综合几种传统方法, 仍能探索微生物群落在土壤剖面中利用碳源的规律。微生物群落AWCD, 多样性指数和碳源利用数量都随着深度增加而减少, 其中多样性指数与SOC和TN呈极显著正相关关系(表 3), 说明剖面中微生物群落多样性随养分含量减少而减少。值得注意的是, 草地与林地的多样性指数与pH呈极显著负相关, 但是农田的多样性指数却没有显著相关, 这一点也与前期有机碳在剖面分布结果相同[25], 都证明了人类生产活动对土壤属性会产生显著影响。
| 处理Treatments | 有机碳SOC | 全氮TN | 碱解氮EN | PH |
| CL | 0.910** | 0.851** | 0.873** | -0.349** |
| GL | 0.779** | 0.726** | 0.771** | -0.743** |
| FL | 0.922** | 0.878** | 0.892** | -0.886** |
| **P < 0.01 | ||||
黑土农田剖面中由于每年的翻耕和机械压实产生了犁底层[30], 阻挡了根系的生长, 所以每种碳源利用程度都在0-20 cm较高, 在20 cm以下逐渐下降。地上植被转变后经过植被自然生长, 犁底层逐渐消失, 根系活动的土层加深导致剖面中不同层次微生物群落对6类碳源的利用发生变化(图 4)。在0-10 cm土层中林地对羧酸类碳源的利用显著低于草地和农田。而且林地剖面中微生物群落对碳源的利用强度在40 cm以下土层才逐渐下降; 而草地剖面中微生物群落在除糖类和胺类碳源利用强度变化与农田相似外, 氨基酸类、羧酸类和酚酸类碳源利用强度在40 cm以下土层逐渐减弱, 而聚合物类碳源利用强度在60 cm以下在逐渐减弱。从主成分分析结果中可以看到, 根系已经影响到100 cm以下的土壤微生物群落, 但是并没有影响到180-200 cm土层的微生物群落。
本研究应用的Biolog技术由于其操作简便广泛应用于微生物群落功能多样性的研究中。但是本身也有一定的局限性, 其中所含的碳源具有一定的人为倾向, 并没有包含土壤中所有种类的化合物, 而且只能检测到生长较快的微生物类群, 这样有可能低估土壤微生物群落的实际代谢水平[31]。因此, 在今后的研究中尽量结合多种研究方法, 得到更全面的更客观的研究结果。
4 结论对黑土不同利用方式下土壤剖面中微生物群落碳源利用特征研究表明, 从农田植被转变为恢复草地增加了可培养微生物数量, 转变为人工林地后减少了可培养微生物数量。植被根系对可培养微生物数量的影响在0-60 cm之间。恢复草地和人工林地除了改变表层微生物群落活性和碳源利用强度外, 还改变了100 cm以下的微生物群落碳源代谢特征, 增加了剖面深层中的微生物活性及各类碳源的利用强度, 促进了剖面中养分与物质的循环。
| [1] | 贺纪正, 陆雅海, 傅博杰. 土壤生物学前沿. 北京: 科学出版社, 201529–29. |
| [2] | 黄昌勇. 土壤学. 北京: 中国农业出版社, 200051–51. |
| [3] | Nannipieri P, Ascher J, Ceccherini M T, Landi L, Pietramellara G, Renella G. Microbial diversity and soil functions. European Journal of Soil Science , 2003, 54 (4) : 655–670. DOI:10.1046/j.1351-0754.2003.0556.x |
| [4] | Van Der Heijden M G A, Bardgett R D, Van Straalen N M. The unseen majority:soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecology Letters , 2008, 11 (3) : 296–310. DOI:10.1111/ele.2008.11.issue-3 |
| [5] | 林先贵, 胡君利. 土壤微生物多样性的科学内涵及其生态服务功能. 土壤学报 , 2008, 45 (5) : 892–900. |
| [6] | 孙波, 廖红, 苏彦华, 许卫锋, 蒋瑀霁. 土壤-根系-微生物系统中影响氮磷利用的以下关键协同机制的研究进展. 土壤 , 2015, 47 (2) : 210–219. |
| [7] | Gyaneshwar P, Kumar G N, Parekh L J, Poole P S. Role of soil microorganisms in improving P nutrition of plants. Plant and Soil , 2002, 245 (1) : 83–93. DOI:10.1023/A:1020663916259 |
| [8] | Li C H, Yan K, Tang L S, Jia Z J, Li Y. Change in deep soil microbial communities due to long-term fertilization. Soil Biology and Biochemistry , 2014, 75 : 264–272. DOI:10.1016/j.soilbio.2014.04.023 |
| [9] | Eilers K G, Debenport S, Anderson S, Fierer N. Digging deeper to find unique microbial communities:the strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biology and Biochemistry , 2012, 50 : 58–65. DOI:10.1016/j.soilbio.2012.03.011 |
| [10] | Preston-Mafham J, Boddy L, Randerson P F. Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles-a critique. FEMS Microbiology Ecology , 2002, 42 (1) : 1–4. |
| [11] | 郑华, 欧阳志云, 方治国, 赵同谦. BIOLOG在土壤微生物群落功能多样性研究中的应用. 土壤学报 , 2004, 41 (3) : 456–461. |
| [12] | Choi K H, Dobbs F C. Comparison of two kinds of Biolog microplates (GN and ECO) in their ability to distinguish among aquatic microbial communities. Journal of Microbiological Methods , 1999, 36 (3) : 203–213. DOI:10.1016/S0167-7012(99)00034-2 |
| [13] | Garland J L. Analysis and interpretation of community-level physiological profiles in microbial ecology. FEMS Microbiology Ecology , 1997, 24 (4) : 289–300. DOI:10.1111/j.1574-6941.1997.tb00446.x |
| [14] | Garland J L, Mills A L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied and Environmental Microbiology , 1991, 57 (8) : 2351–2359. |
| [15] | Sun B, Wang X Y, Wang F, Jiang Y J, Zhang X X. Assessing the relative effects of geographic location and soil type on microbial communities associated with straw decomposition. Applied and Environmental Microbiology , 2013, 79 (11) : 3327–3335. DOI:10.1128/AEM.00083-13 |
| [16] | Yin R, Deng H, Wang H L, Zhang B. Vegetation type affects soil enzyme activities and microbial functional diversity following re-vegetation of a severely eroded red soil in sub-tropical China. Catena , 2014, 115 : 96–103. DOI:10.1016/j.catena.2013.11.015 |
| [17] | 刘明, 李忠佩, 张桃林. 不同利用方式下红壤微生物生物量及代谢功能多样性的变化. 土壤 , 2009, 41 (5) : 744–748. |
| [18] | 石洪艾, 尤孟阳, 李禄军, 韩晓增. 长期施用有机物料下黑土氮素有效性及其与作物产量的关系. 生态学杂志 , 2012, 31 (9) : 2283–2288. |
| [19] | 李振高, 骆永明, 滕应. 土壤与环境微生物研究法. 北京: 科学出版社, 200852–52. |
| [20] | Classen A T, Boyle S I, Haskins K E, Overby S T, Hart S C. Community-level physiological profiles of bacteria and fungi:plate type and incubation temperature influences on contrasting soils. FEMS Microbiology Ecology , 2003, 44 (3) : 319–328. DOI:10.1016/S0168-6496(03)00068-0 |
| [21] | 夏雪, 谷洁, 车升国, 高华, 秦清军. 施氮水平对塿土微生物群落和酶活性的影响. 中国农业科学 , 2011, 44 (8) : 1618–1627. |
| [22] | Kirk J L, Beaudette L A, Hart M, Moutoglis P, Klironomos J N, Lee H, Trevors J T. Methods of studying soil microbial diversity. Journal of Microbiological Methods , 2004, 58 (2) : 169–188. DOI:10.1016/j.mimet.2004.04.006 |
| [23] | 宋长青, 吴金水, 陆雅海, 沈其荣, 贺纪正, 黄巧云, 贾仲君, 冷疏影, 朱永官. 中国土壤微生物学研究10年回顾. 地球科学进展 , 2013, 28 (10) : 1087–1105. |
| [24] | Fierer N, Schimel J P, Holden P A. Variations in microbial community composition through two soil depth profiles. Soil Biology and Biochemistry , 2003, 35 (1) : 167–176. DOI:10.1016/S0038-0717(02)00251-1 |
| [25] | 郝翔翔, 韩晓增, 李禄军, 邹文秀, 陆欣春, 乔云发. 土地利用方式对黑土剖面有机碳分布及碳储量的影响. 应用生态学报 , 2015, 26 (4) : 965–972. |
| [26] | Fanin N, Bertrand I. Aboveground litter quality is a better predictor than belowground microbial communities when estimating carbon mineralization along a land-use gradient. Soil Biology and Biochemistry , 2016, 94 : 48–60. DOI:10.1016/j.soilbio.2015.11.007 |
| [27] | 趙洪书. 从容重计算土壤总孔隙度的初步探讨. 土壤学报 , 1964, 12 (2) : 230–234. |
| [28] | Lambin E F, Turner B L, Geist H J, Agbola S B, Angelsen A, Bruce J W, Coomes O T, Dirzo R, Fischer G, Folke C, George P S, Homewood K, Imbernon J, Leemans R, Li X B, Moran E F, Mortimore M, Ramakrishnan P S, Richards J F, Skånes H, Steffen W, Stone G D, Svedin U, Veldkamp T A, Vogel C, Xu J C. The causes of land-use and land-cover change:moving beyond the myths. Global Environmental Change , 2001, 11 (4) : 261–269. DOI:10.1016/S0959-3780(01)00007-3 |
| [29] | 李晨华, 张彩霞, 唐立松, 熊正琴, 王保战, 贾仲君, 李彦. 长期施肥土壤微生物群落的剖面变化及其与土壤性质的关系. 微生物学报 , 2014, 54 (3) : 319–329. |
| [30] | 韩晓增, 邹文秀, 王凤仙, 王凤菊. 黑土肥沃耕层构建效应. 应用生态学报 , 2009, 20 (12) : 2996–3002. |
| [31] | 章家恩, 蔡燕飞, 高爱霞, 朱丽霞. 土壤微生物多样性实验研究方法概述. 土壤 , 2004, 36 (4) : 346–350. |