 2015, Vol. 35
2015, Vol. 35文章信息
- 徐万里, 唐光木, 葛春辉, 王西和, 刘骅
- XU Wanli, TANG Guangmu, GE Chunhui, WANG Xihe, LIU Hua
- 长期施肥对新疆灰漠土土壤微生物群落结构与功能多样性的影响
- Effects of long-term fertilization on diversities of soil microbial community structure and function in grey desert soil of Xinjiang
- 生态学报, 2015, 35(2): 468-477
- Acta Ecologica Sinica, 2015, 35(2): 468-477
- http://dx.doi.org/10.5846/stxb201303310567
-
文章历史
- 收稿日期:2013-03-31
- 网络出版日期:2014-03-25






施肥是农业生产的主要措施之一,长期施肥将明显改变作物产量[1]、土壤理化性质[2]和生物活性[3, 4],对土壤质量和可持续利用具有深远影响。微生物是土壤生物中最重要的组成部分,在土壤营养元素循环、土壤肥力形成和发展、生态环境改善、植物生长和土传病害防治等许多方面起着极其重要的作用。研究表明,施肥使土壤微生物群落结构发生显著变化[5, 6],长期平衡施肥使土壤微生物量碳氮和微生物功能活性增强[7, 8],有机肥与化肥长期配施条件下氨氧化细菌的多样性高于化肥处理[9];长期施用有机肥的土壤反硝化细菌多样性高于不施肥和施用化肥的土壤[10];长期施用氮钾肥和秸秆还田对稻田甲烷氧化菌多样性有重要的影响,其中氮肥甲烷氧化菌的数量有抑制作用[11];长期施用有机肥和秸秆还田有利于黄土高原土壤有机碳的积累,从而增加了丛枝菌根的多样性[12]。在研究方法上,由于土壤中微生物数量庞大,种类繁多,且存在大量无法培养的微生物类群,采用传统的培养方法,无法全面了解土壤微生物信息,因此近年来采用磷脂脂肪酸法、碳源代谢利用(Biolog法)和分子生物学等的方法来研究土壤微生物多样性已成为热点[13, 14, 15, 16, 17, 18, 8]。
灰漠土是典型的干旱荒漠地带土壤,是新疆主要的农业耕作土壤之一。但目前新疆灰漠土上长期施肥对土壤微生物群落和功能多样性的研究很少[19],采用Biolog法来揭示长期施肥对灰漠土土壤微生物碳源利用的影响还未见报道。为此,本文采用传统微生物培养和Biolog测定相结合的方法,研究长期定位施肥对灰漠土土壤微生物学特性的影响,为揭示新疆灰漠土质量演变规律、土壤肥力保持、土壤微生态系统健康发展和作物产量提高等方面提供科学的理论依据。
1 材料与方法 1.1 供试土壤供试土壤来自新疆乌鲁木齐市国家灰漠土肥力与肥料效益重点野外科学观测试验站(N:43°95′26″,E:87°46′45″)。该试验站位于新疆乌鲁木齐市以北22 km的新疆农业科学院安宁区综合试验场内。国家灰漠土肥力与肥料效益重点野外科学观测试验站海拔高度680—920 m,年均气温5—7℃,年降水量180—250 mm,年蒸发量1600—2200 mm,年均日照时数2594 h,无霜期156 d,属干旱半干旱荒漠气候。试验土壤为第四纪黄土状沉积物,属灰漠土土类中度熟化灰漠土,在北疆农耕区具有典型性和代表性。试验地的土壤基本养分含量见表 1。
| 处理 Treatments | 有机质 OM/(g/kg) | 全氮 Total N/(g/kg) | 全磷 Total P/(g/kg) | 有效氮 Avail. N/(mg/kg) | 有效磷 Avail. P/(mg/kg) | 有效钾 Avail. K/(mg/kg) |
| CK0:不耕作施肥abandonment; CK:耕作不施肥cultivated without fertilization; N:单施氮肥only nitrogen fertilizer; NK:施氮钾肥nitrogen and potassium fertilizer; NP:施氮磷肥nitrogen and phosphate fertilizer; PK:施磷钾肥phosphate and potassium fertilizer; NPK:施氮磷钾肥nitrogen,phosphate and potassium fertilizer; NPKM1:施氮磷钾+常量有机肥NPK and organic materials; NPKM2:施氮磷钾+增量有机肥NPK and double organic materials; NPKS:施氮磷钾+秸秆还田 NPK and straw | ||||||
| CK0 | 14.57 | 0.86 | 0.85 | 66.81 | 12.19 | 370.76 |
| CK | 13.21 | 0.62 | 0.57 | 11.46 | 1.12 | 190.99 |
| N | 13.36 | 0.57 | 0.60 | 42.00 | 1.29 | 176.34 |
| NK | 15.47 | 0.54 | 0.68 | 45.82 | 2.52 | 294.90 |
| NP | 15.11 | 0.69 | 0.76 | 49.64 | 9.67 | 115.79 |
| PK | 14.71 | 0.60 | 0.81 | 26.73 | 19.02 | 179.12 |
| NPK | 15.26 | 0.81 | 0.85 | 34.37 | 13.44 | 494.81 |
| NPKM1 | 27.29 | 1.66 | 0.84 | 91.64 | 37.00 | 641.28 |
| NPKM2 | 38.50 | 1.39 | 1.11 | 133.65 | 97.64 | 1021.29 |
| NPKS | 13.34 | 0.39 | 0.64 | 45.82 | 8.51 | 179.12 |
本试验从长期肥料肥效试验(20a)中选择以下10个处理:(1)不耕作,不施肥(撂荒,CK0);(2)耕作,不施肥(CK);(3)单施氮肥(N);(4)施氮钾肥(NK);(5)施氮磷肥(NP);(6)施磷钾肥(PK);(7)施氮磷钾肥(NPK);(8)施氮磷钾+常量有机肥(NPKM1);(9)施氮磷钾+增量有机肥(NPKM2),即氮、磷、钾和有机肥施用量均是常量处理的1.5倍;(10)施氮磷钾+秸秆还田(NPKS)。每年施N 242 kg/hm2、P2O5 138 kg/hm2、K2O 60 kg/hm2,施肥时60%的化学氮肥及100%化学磷、钾肥和有机肥作基肥,在播种前均匀撒施地表,深翻后播种,剩余40%的化学氮肥作追肥。所有施氮处理的施氮总量相同;有机肥为羊粪,常量有机肥施用量为30000 kg/hm2,每年秋季一次施入翻地。秸秆还田为当季作物全部秸秆粉碎还田。试验种植方式为玉米-冬小麦-棉花轮作。
1.3 样品采集及测定方法 1.3.1 样品采集2010年9月采集以上10组处理土样。采用5点法随机采集耕层0—20 cm土壤样品,每个土样3次重复。将土壤样品混合均匀后4℃保存,进行微生物计数和Biolog碳源利用测定。
1.3.2 土壤微生物数量测定[20]采用稀释平板涂抹培养计数法分析。细菌采用牛肉膏蛋白胨琼脂培养基,放线菌采用改良的高氏一号培养基(每300 mL培养基中加3%重铬酸钾1 mL),真菌采用PDA培养基(每100 mL培养基加1%链霉素溶液0.3 mL)。微生物生理菌群数量,采用稀释液体培养法(MPN法)计数。好氧性自生固氮菌采用阿须贝无氮培养基,纤维素分解菌采用赫奇逊氏培养基,氨化细菌用蛋白胨液体培养基,亚硝化细菌用铵盐培养液(由于土壤中很少有亚硝酸盐的长期累积,所以测定亚硝化细菌的数量就可以反映硝化细菌的数量[21])。
1.3.3 土壤微生物群落碳源代谢利用测定采用 Biolog ECO微平板法进行[20]。称取10 g鲜土加入l00 mL灭菌的生理盐水(0.85%)中,摇匀,静止片刻,然后将土壤样品稀释至10-3。取150 μL菌悬液接种到生态板的每一个孔中,25℃恒温培养,每隔24 h分别在590 nm波长下读数,连续培养7 d。取120 h的平均光密度值进行主成分分析。
1.3.4 数据处理与分析微平板孔中溶液吸光值平均颜色变化率(AWCD)用于描述土壤微生物代谢活性,计算公式如下:
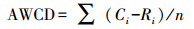
式中,Ci为每个有培养基孔的吸光值,Ri为对照孔的吸光值,n为培养基孔数,Biolog Eco板n值为31。采用Simpson、Shannon-Wiener和McIntosh 3个指数来表征土壤微生物群落功能多样性。其中Simpson指数(D)用于评估某些最常见种的优势度,Shannon-Wiener指数(H)用于评估物种的丰富度,McIntosh指数(U)用于评估群落物种均匀度:
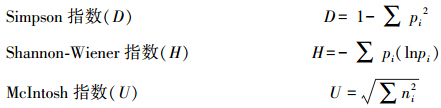
式中,pi为第i孔的相对吸光值与所有整个微平板的相对吸光值总和的比值(Ci-Ri)/∑(Ci-Ri);ni为第i孔的相对吸光值(Ci-Ri)。
数据采用DPS v9.50版软件进行平均吸光值(AWCD)、多样性指数、主成分分析(PCA)和相关的方差分析等工作。
2 结果与分析 2.1 长期施肥对灰漠土土壤微生物群落结构多样性的影响微生物是土壤生态系统重要组成部分,其变化能敏感地反映土壤质量和肥力的变化[22]。从表 2可以看出,与CK处理相比,CK0处理细菌数量有所增加,不同化肥处理(PK除外)、NPKM1和NPKM2处理可增加土壤细菌的数量,不同化肥处理增幅在8.14%—135.70%之间,NPKM1和NPKM2分别增加了162.20%和173.75%;PK和NPKS处理,土壤中细菌数量则分别减少了15.22%和14.17%。从表中还可以看出,CK0处理土壤放线菌数量最高,耕作施肥减少了放线菌数量;不同施肥处理间,长期施化肥(除PK和N处理外)处理相比CK处理增加了土壤中放线菌数量,其增幅在15.30%—44.78%之间; NPKM1和NPKM2处理放线菌数量分别增加了34.39%和39.37%; NPKS处理土壤放线菌数量则减少了21.27%。真菌数量CK0处理相对较高,耕作和施肥后(除NPKM2外)则大幅降低;与CK相比,不同化肥处理(除NK处理外)真菌数量均有一定幅度的降低,其降幅在10.00%—38.33%之间; NPKM1和NPKM2处理土壤真菌明显增加,分别增加了63.33%和488.33%,但NPKS处理的土壤真菌数量却明显降低,减少了63.33%。
|
处理
Treatments | 细菌
Bacteria / (×106 cfu/g) | 放线菌
Actinomycete/ (×105 cfu/g) | 真菌
Fungi / (×104 cfu/g) | 自生固氮菌
Azotobacter/ (×106 cfu/g) | 氨化细菌
Ammoniation/ (×106 cfu/g) | 亚硝化细菌
Nitrifier / (×102 cfu/g) | 纤维素分解菌
Cytophaga / (×103 cfu/g) |
| 每组同列的不同字母表示差异显著性水平; 小写字母为 5%显著水平; CK0: 不耕作施肥; CK: 耕作不施肥; N: 单施氮肥; NK: 施氮钾肥; NP: 施氮磷肥; PK: 施磷钾肥; NPK: 施氮磷钾肥; NPKM1: 施氮磷钾+常量有机肥; NPKM2: 施氮磷钾+增量有机肥; NPKS: 施氮磷钾+秸秆还田 | |||||||
| CK0 | 4.75±0.40d | 11.05±0.76a | 3.08±0.12b | 0.32±0.06e | 7.65±0.49b | 0.42±0.03g | 16.07±0.15b |
| CK | 3.81±0.62de | 5.36±0.36d | 0.60±0.21cde | 1.40±0.91c | 1.07±0.10f | 0.80±0.03f | 3.52±0.05d |
| N | 4.12±0.20de | 3.94±0.55e | 0.42±0.03de | 0.07±0.02f | 1.72±0.07ef | 2.34±0.08b | 1.55±0.08e |
| NK | 6.98±0.30c | 6.18±0.38cd | 0.80±0.15cd | 1.52±0.07c | 7.39±0.42b | 0.71±0.05f | 27.07±1.03a |
| NP | 8.98±0.39b | 7.76±0.53b | 0.37±0.05e | 0.26±0.07ef | 7.31±0.17b | 0.88±0.04f | 1.31±0.06e |
| PK | 3.23±0.56e | 5.32±0.92d | 0.54±0.37de | 2.77±0.75a | 2.11±0.12e | 0.82±0.07f | 1.24±0.08e |
| NPK | 6.41±0.80c | 7.43±0.07b | 0.54±0.29de | 0.76±0.16d | 5.30±0.41c | 1.93±0.08c | 10.00±0.10c |
| NPKM1 | 9.99±0.21ab | 7.20±0.62bc | 0.98±0.30c | 1.96±0.54b | 6.98±0.53b | 4.30±0.10a | 15.58±0.36b |
| NPKM2 | 10.43±0.65a | 7.47±0.61b | 3.53±0.89a | 1.55±0.05c | 11.92±0.32a | 1.64±0.06d | 3.03±0.08d |
| NPKS | 3.27±0.74e | 4.22±0.55e | 0.22±0.08e | 0.08±0.01f | 3.19±0.06d | 1.36±0.02e | 1.64±0.05e |
细菌生理菌群受土壤特性和环境影响而在土壤中呈现不同的数量分布,各生理菌群的分布在一定程度上反映出土壤养分的利用状况和健康程度[23]。从研究结果(表 2)可以看出长期定位不同施肥处理造成土壤环境的不同,可引起土壤中生理菌群数量发生显著变化。CK处理自生固氮菌和亚硝化细菌数量高于CK0处理,氨化细菌和纤维素分解菌数量则低于CK0处理;长期施用化肥的土壤中,与CK相比,NPK处理氨化细菌、亚硝化细菌和纤维素分解菌的数量较好,分别增加了395.33%、141.25%和184.09%;NPKM1和NPKM2处理可提高土壤中与氮素转化有关的生理菌群数量,自生固氮菌分别增加了40%和10.71%,氨化细菌分别增加了552.34%和1014.02%,亚硝化细菌分别增加了437.50%和105.00%,但与碳素转化相关的纤维素分解菌数量,NPKM1增加了342.61%,而NPKM2处理纤维素分解菌数量反而降低了13.92%;NPKS处理氨化细菌和亚硝化细菌数量增加了198.13%和70.00%,自生固氮菌和纤维素分解菌数量降低了94.29%和53.41%。
2.2 长期施肥对灰漠土土壤微生物群落功能多样性的影响 2.2.1 平均颜色变化率平均颜色变化率(AWCD)是表征土壤微生物群落对底物碳源利用强度的指标,AWCD值越大,表明对碳源利用强度越强,微生物代谢活性越高[24]。从图 1可以看出,不同施肥处理土壤的AWCD值随培养时间的延长而提高。在培养的0—24 h内不同处理土壤AWCD值变化不明显,24—72 h内则表现出快速增长的趋势,微生物活性进入对数增长期;在培养的72 h后,在培养时间内,AWCD值随培养时间的延长增长趋于缓慢。从培养处于稳定期的96 h来看,AWCD值在0.69—1.14之间,AWCD值表现为NK> NPKM1、NPKM2>N>CK>NPK>NPKS>PK>CK0>NP,NK处理最高,活性最强,NP处理最低。

|
| 图 1 长期施肥土壤平均颜色变化率 Fig. 1 AWCD of the Long-term fertilization soil CK0:不耕作施肥abandonment; CK:耕作不施肥cultivated without fertilization; N:单施氮肥only nitrogen fertilizer; NK:施氮钾肥nitrogen and potassium fertilizer; NP:施氮磷肥nitrogen and phosphate fertilizer; PK:施磷钾肥phosphate and potassium fertilizer; NPK:施氮磷钾肥nitrogen,phosphate and potassium fertilizer; NPKM1:施氮磷钾+常量有机肥NPK and organic materials; NPKM2:施氮磷钾+增量有机肥NPK and double organic materials; NPKS:施氮磷钾+秸秆还田 NPK and straw |
根据不同处理碳源利用情况,综合考虑其变化趋势,选取光密度趋于稳定,且不同处理之间有较好分形的96 h的AWCD值进行土壤微生物群落代谢多样性的分析(表 3)。从表中可以看出,Simpson优势度指数除撂荒地较高外,其余处理差异均不明显,Shannon丰富度指数各处理间有一定的差异。其中NK、NPKM1和NPKM2处理的Shannon指数高于CK0;其余处理均低于CK0,但CK、N和NPK处理与CK0差异不显著; NP、PK和NPKS处理Shannon指数较低。从表中还可以看出,McIntosh均匀度指数表现为CK0>NP>NK、NPKM1、NPKM2、NPK、NPKS、PK>N>CK,CK0处理与其他处理间差异显著(P<0.05)。
| 处理 Treatments | 平均颜色变化率 AWCD | 优势度指数 Simpson(D) | 丰富度指数 Shannon(H) | 均匀度指数 McIntosh(U) |
| 每组同列的不同字母表示差异显著性水平; 小写字母为5%显著水平; CK0:不耕作施肥;CK:耕作不施肥;N:单施氮肥;NK:施氮钾肥;NP:施氮磷肥;PK:施磷钾肥;NPK:施氮磷钾肥;NPKM1:施氮磷钾+常量有机肥;NPKM2:施氮磷钾+增量有机肥;NPKS:施氮磷钾+秸秆还田 | ||||
| CK0 | 0.69±0.23de | 0.99±0.02a | 3.02±0.04ab | 0.98±0.05a |
| CK | 0.99±0.05ab | 0.97±0.00b | 2.99±0.05ab | 0.92±0.00b |
| N | 1.01±0.09ab | 0.97±0.00b | 2.98±0.02ab | 0.92±0.00b |
| NK | 1.14±0.06a | 0.98±0.00b | 3.08±0.09a | 0.93±0.01b |
| NP | 0.52±0.05e | 0.98±0.01b | 2.73±0.14d | 0.95±0.03ab |
| PK | 0.73±0.28cde | 0.97±0.00b | 2.84±0.13cd | 0.93±0.02b |
| NPK | 0.94±0.18abc | 0.98±0.00b | 3.00±0.07ab | 0.93±0.01b |
| NPKM1 | 1.06±0.06a | 0.98±0.00b | 3.07±0.04a | 0.93±0.01b |
| NPKM2 | 1.06±0.01a | 0.98±0.00b | 3.09±0.04a | 0.93±0.01b |
| NPKS | 0.80±0.15bcd | 0.97±0.01b | 2.91±0.04bc | 0.93±0.02b |
利用培养96h的AWCD值,对长期施肥土壤微生物利用单一碳源特性进行主成分分析,同时提取特征值大于1的主成分的特征根及方差贡献率。本研究共提取出7个主成分,累计贡献率达95.82%。其中第一主成分(PC1)的特征根为13.09,方差贡献率为42.24%;第二主成分(PC2)的特征根为4.56,方差贡献率为14.69%;第三主成分(PC3)的特征根为3.82,方差贡献率为12.34%。其余4—7主成分贡献率均小于10%。
选取方差贡献率最高的前两个主成分PC1和PC2进行微生物群落功能多样性分析。主成分分析表明(图 2),不同施肥处理碳源利用在PC轴上差异显著,在PC1轴上,CK、N、NK、NPK、NPKM1和NPKM2处理分布在正方向上,CK0、NP、PK和NPKS处理分布在负方向上;在PC2轴上,CK0、CK、N、NK和NP处理分布在正方向上,NPKM1、NPKM2、NPK、NPKS和PK处理分布在负方向上。

|
| 图 2 长期施肥土壤微生物群落主成分分析 Fig. 2 Principal component analysis of soil microbial community CK0:不耕作施肥;CK:耕作不施肥;N:单施氮肥;NK:施氮钾肥;NP:施氮磷肥;PK:施磷钾肥;NPK:施氮磷钾肥;NPKM1:施氮磷钾+常量有机肥;NPKM2:施氮磷钾+增量有机肥;NPKS:施氮磷钾+秸秆还田 |
初始载荷因子反映主成分与碳源利用的相关系数,载荷因子越高,表示该碳源对主成分的影响越大[25]。从表 4中可以看出,对第一主成分(PC1)贡献较大的碳源有18种,主要包括糖类(5种)、氨基酸类(3种)、羧酸类(5种)、聚合物类(1种)、胺类(2种)和其他类(2种);对第二主成分(PC2)贡献较大的碳源有5种,主要包括氨基酸类(2种)、羧酸类(2种)和胺类(1种)。糖类、氨基酸类、羧酸类和胺类是微生物利用的主要碳源。
从图 2和表 4综合分析可以看出,说明长期不同施肥处理的土壤微生物碳源利用能力存在差异,且不同处理在主成分轴上的差异,是和聚集在该轴上微生物利用碳底物的利用能力相关联的。在PC1上,CK、N、NK、NPK、NPKM1和NPKM2处理对糖类(N-乙酰基-D-葡萄胺、D-纤维二糖、D-木糖、D-甘露醇和β-甲基D-葡萄糖苷)、氨基酸(L-苯基丙氨酸、L-苏氨酸和L-丝氨酸)、羧酸类(衣康酸、2-羟基苯甲酸、D-苹果酸、y-羟基丁酸和D-半乳糖内酯)和胺类(苯乙基胺和腐胺)等碳源的利用能力高于CK0、NP、PK和NPKS处理。在PC2上,CK0、CK、N、NK和NP处理对L-天冬酰胺酸、L-精氨酸、4-羟基苯甲酸和苯乙基胺的利用能力较强,而NPKM1、NPKM2、NPK、NPKS和PK处理对D-苹果酸的利用能力较强。因此施肥对灰漠土中微生物群落在碳源的利用能力的影响是导致其活性差异的主要原因。
| 碳源类型Carbon source | 第一主成分PC1 | 第二主成分PC2 | |
| 糖类Carbohydrates | N-乙酰基-D-葡萄胺N-Acetyl-D-Glucosamine | 0.93 | -0.01 |
| D-纤维二糖D-Cellobiose | 0.91 | 0.07 | |
| D-木糖D-Xylose | 0.87 | -0.24 | |
| D-甘露醇D-Mannitol | 0.87 | -0.18 | |
| β-甲基D-葡萄糖苷β-Methyl-D-Glucoside | 0.81 | -0.01 | |
| 氨基酸Amino acids | L-苯基丙氨酸L-Phenylalanine | 0.73 | -0.01 |
| L-天冬酰胺酸L-Asparagine | 0.19 | 0.86 | |
| L-精氨酸L-Arginine | 0.48 | 0.65 | |
| L-苏氨酸L-Threonine | 0.61 | 0.45 | |
| L-丝氨酸L-Serine | 0.73 | 0.19 | |
| 羧酸类Carboxylic acids | 衣康酸Itaconic Acid | 0.88 | -0.08 |
| 2-羟基苯甲酸2-Hydroxy Benzoic Acid | 0.68 | 0.12 | |
| 4-羟基苯甲酸4-Hydroxy Benzoic Acid | 0.42 | 0.67 | |
| D-苹果酸D-Malic Acid | 0.65 | -0.60 | |
| y-羟基丁酸 y-Hydroxybutyric Acid | 0.76 | -0.22 | |
| D-半乳糖内酯D-Galactonic Acid y-Lactone | 0.82 | 0.07 | |
| 聚合物类Polymers | 吐温80 Tween 80 | 0.65 | 0.49 |
| 胺类Amines/amides | 苯乙基胺Phenylethyl-amine | 0.63 | 0.66 |
| 腐胺Putrescine | 0.81 | -0.43 | |
| 其他类Miscellaneous | 葡萄糖-1-磷酸盐 | 0.65 | -0.37 |
| D,L-a-甘油D,L-a-Glycerol | 0.77 | 0.04 |
应用各种碳源光密度值(96 h)作统计变量进行聚类分析,可以清晰直观的反映各施肥处理间的远近关系[26]。本研究采用欧式距离中的最短距离法。由图 3可以看出,含有氮肥的各种处理(除NP处理外)具有较相似的碳源利用模式;其中NPK、NPKM1、NPKM2、N、NPKS和NK处理间距离接近,具有相似的碳源利用,但与CK0和CK处理距离较远,土壤微生物群落的碳源利用差异显著; PK处理与施氮处理在碳源利用上有一些差异,但与CK0和CK处理较为相似;NP处理与其余处理差异均比较显著。聚类分析表明,施氮处理(除NP外)有着较为相似的碳源利用,其中NPKM1、NPKM2、 N和NK处理具有较高的AWCD值。辜运富认为施N能够促进作物的发育,从而得到更多的根系分泌物,进而提高土壤中微生物的数量[27]。

|
| 图 3 长期施肥土壤微生物群落碳源代谢的聚类分析树状图 Fig. 3 Cluster analysis dendrogram of microbial communities′C sources utilization under Long-term fertilization CK0:不耕作施肥;CK:耕作不施肥;N:单施氮肥;NK:施氮钾肥;NP:施氮磷肥;PK:施磷钾肥;NPK:施氮磷钾肥;NPKM1:施氮磷钾+常量有机肥;NPKM2:施氮磷钾+增量有机肥;NPKS:施氮磷钾+秸秆还田 |
在土壤生态系统中,微生物是土壤物质循环和能量流动的主要参与者,是土壤养分的储备库和周转库[21]。影响土壤微生物的因素很多,如土壤和作物类型、耕作制度和施肥措施等。长期施肥由于改变了土壤的理化性质,进而改变了土壤中微生物群落构成,其数量、生理类群和生态功能都会产生一定变化[28, 29]。
已有的研究表明长期施肥条件下细菌、固氮菌数量与有机质、全氮、碱解氮、全磷、速效磷含量呈显著正相关,真菌、放线菌数量与土壤养分因子之间没有明显的相关性[30],本研究表明不同施肥处理对土壤微生物多样性产生了显著的影响,与土壤养分含量的关系较为密切,但长期施肥对土壤的影响较为复杂,长期形成的养分限制因子、养分丰缺程度以及物理环境不同的土壤,在养分含量与微生物之间出现较大差异。本研究显示,土壤细菌和真菌与土壤养分含量关系密切,而放线菌关系较差。由于土壤中不同生理功能微生物生长、繁殖和代谢所需要的环境条件不同,和长期施肥土壤养分的相关性因菌群不同而有所差异。同时,李秀英的研究显示长期施肥土壤中固氮细菌和氨化细菌数量与土壤养分相关性较强[22],则本研究表明仅氨化细菌与有机质、全N、全P、有效N、有效P和有效K的关系密切,亚硝化细菌和全N的关系较为紧密,其余几类菌群与土壤养分的相关程度均较低,Biolog碳源利用AWCD值与土壤养分的相关程度均较差。
影响土壤微生物碳源利用多样性的主要因素可能与碳源种类,植物残体和根系分泌物、土壤物理性状和化学性质等有密切关系[31]。将长期施肥土壤Biolog 数据中AWCD值、微生物多样性指数与可培养微生物类群进行相关性分析后发现,AWCD值与各种土壤微生物类群之间未达到显著相关水平;从趋势上看,AWCD值除与放线菌之间呈负相关,与其余类群呈正相关关系。其原因可能是可培养微生物的培养基使用的碳源较为单一,与Biolog-Eco板上的碳源不一致造成。但多样性指数与微生物类群之间有相关性,其中微生物碳源利用Simpson(D)指数与放线菌、真菌、氨化细菌和纤维素分解菌之间呈显著相关关系,说明微生物种群的优势物种受这些类群的微生物的影响比较大;Shannon(H)指数与纤维素分解菌之间显著正相关性,微生物种群的丰富度与纤维素分解菌关系密切;McIntosh(U)指数与放线菌、氨化细菌呈显著相关,放线菌和氨化细菌决定了微生物种群的均匀性。
研究表明长期不同施肥处理微生物多样性存在一定的差异。撂荒地由于对土壤的扰动少,土壤结构良好,自然回归土壤的有机物质较多,土壤微生物数量高于长期不施肥和部分单施化肥处理,这与李秀英和梁斌的研究结果一致[22, 32];但养分的不均衡性又表现出具特殊功能的生理类群数量与长期施肥相比并没有明显的提高;由于Biolog微平板上的碳源与根系分泌物有关,受地上部植物和根系分泌物种类多样的影响,撂荒地土壤中利用这些碳源的微生物活性较弱,AWCD值较低,但微生物多样性指数较高,但这一结果与时鹏的研究结果不同[25]。这可能是因为长期不施肥和秸秆不还田处理土壤中养分含量低,微生物数量也相对较低,同时耕作改变了土壤环境[33],耕种的作物种类较为单一,根系分泌物有一定的累积,利用这些根系分泌物的微生物增多,Biolog碳源利用活性(AWCD值)高于撂荒地,但多样性指数较低。
长期采用不同的施肥模式,土壤有机质含量、组成会有较大的变化,土壤微生物多样性也呈现较大的差异。本研究表明除个别缺素施肥处理微生物活性低于不施肥处理外,其余处理对土壤微生物多样性有一定程度的提升;由于土壤中某些营养元素的缺失,提升效果没有NPKM和NPK处理显著。秸秆还田处理对土壤微生物多样性的影响还存在一些争议,较多文献报道旱地红壤秸秆还田细菌数量较其余处理低,而放线菌和真菌数量较高[34];化肥有机肥配施使红壤微生物碳源利用率增强而秸秆还田对其有不利影响[35, 36],本研究表明NPKS处理对微生物多样性并没有提升作用,分析原因可能与不同土壤类型和气候条件有关[35],且因为秸秆还田后无机N、P相对缺乏且C / N比高,而养分多是缓效的,微生物分解慢,同时改变了土壤微生物优势种群,引起微生物多样性低下[36]。
由于可培养的微生物数量只占土壤微生物数量的极小一部分,而Biolog生态板使用的碳源多与植株产生的根系分泌物关系密切,具有一定的局限性,如结合先进的分子生物学方法,可以使结果更为全面、合理。
| [1] | 王飞, 林诚, 李清华, 何春梅, 李昱, 林新坚. 长期不同施肥处理对南方黄泥田水稻子粒品质性状与土壤肥力因子的影响. 植物营养与肥料学报, 2011, 17(2): 283-290. |
| [2] | Liang B, Yang X Y, He X H, Zhou J B. Effects of 17-year fertilization on soil microbial biomass C and N and soluble organic C and N in loessial soil during maize growth. Biology and Fertility of Soils, 2011, 47(2): 121-128. |
| [3] | 马宁宁, 李天来, 武春成, 张恩平. 长期施肥对设施菜田土壤酶活性及土壤理化性状的影响. 应用生态学报, 2010, 21(7): 1766-1771. |
| [4] | Ge G F, Li Z J, Fan F L, Chu G X, Hou Z N, Liang Y C. Soil biological activity and their seasonal variations in response to long-term application of organic and inorganic fertilizers. Plant ant Soil, 2010, 326(1/2): 31-44. |
| [5] | Zhong W H, Gu T, Wang W, Zhang B, Lin X G, Huang Q R, Shen W S. The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant ant Soil, 2010, 326(1/2): 511-522. |
| [6] | 张奇春, 王雪芹, 时亚南, 王光火. 不同施肥处理对长期不施肥区稻田土壤微生物生态特性的影响. 植物营养与肥料学报, 2010, 16(1): 118-123. |
| [7] | Hu J L, Lin X G, Wang J H, Dai J, Chen R R, Zhang J B, Wong M H. Microbial functional diversity, metabolic quotient, and invertase activity of a sandy loam soil as affected by long-term application of organic amendment and mineral fertilizer. Journal of Soils and Sediments, 2011, 11(2): 271-280. |
| [8] | 张志明, 许艳丽, 韩晓增, 李晓慧. 连续施肥对农田黑土微生物功能多样性的影响. 生态学杂志, 2012, 31(3): 647-651. |
| [9] | 裴雪霞, 周卫, 梁国庆, 孙静文, 王秀斌, 李双来. 长期施肥对黄棕壤性水稻土氨氧化细菌多样性的影响. 植物营养与肥料学报, 2011, 17(3): 724-730. |
| [10] | 陈哲, 袁红朝, 吴金水, 魏文学. 长期施肥制度对稻田土壤反硝化细菌群落活性和结构的影响. 生态学报, 2009, 29(11): 5923-5929. |
| [11] | Zheng Y, Zhang L M, Zheng Y M, Di H J, He J Z. Abundance and community composition of methanotrophs in a Chinese paddy soil under long-term fertilization practices. Journal of Soils and Sediments, 2008, 8(6): 406-414. |
| [12] | Wu F S, Dong M X, Liu Y J, Ma X J, An L Z, Young J P W, Feng H Y. Effects of long-term fertilization on AM fungal community structure and Glomalin-related soil protein in the Loess Plateau of China. Plant ant Soil, 2011, 342(1/2): 233-247. |
| [13] | Islam R, Chauhan P S, Kim Y, Kim M, Sa T M. Community level functional diversity and enzyme activities in paddy soils under different long-term fertilizer management practices. Biology and Fertility of Soils, 2011, 47(5): 599-604. |
| [14] | Ge Y, Zhang J B, Zhang L M, Yang M, He J Z. Long-term fertilization regimes affect bacterial community structure and diversity of an agricultural soil in northern China. Journal of Soils and Sediments, 2008, 8(1): 43-50. |
| [15] | 毕明丽, 宇万太, 姜子绍, 马强, 张璐, 徐永刚. 利用PLFA方法研究不同土地利用方式对潮棕壤微生物群落结构的影响. 中国农业科学, 2010, 43(9): 1834-1842. |
| [16] | 王霞, 陈哲, 袁红朝, 吴敏娜, 魏文学, 吴金水, 秦红灵. 应用16S rDNA克隆文库技术研究长期稻草还田对水稻土细菌多样性的影响. 生态学报, 2010, 30(13): 3865-3874. |
| [17] | 袁红朝, 秦红灵, 刘守龙, 童成立, 魏文学, 吴金水. 长期施肥对红壤性水稻土细菌群落结构和数量的影响. 中国农业科学, 2011, 44(22): 4610-4617. |
| [18] | 李晨华, 贾仲君, 唐立松, 吴宇澄, 李彦. 不同施肥模式对绿洲农田土壤微生物群落丰度与酶活性的影响. 土壤学报, 2012, 49(3): 567-574. |
| [19] | 刘骅, 林英华, 张云舒, 谭新霞, 王西和. 长期施肥对灰漠土生物群落和酶活性的影响. 生态学报, 2008, 28(8): 3898-3904. |
| [20] | 姚槐应, 黄昌勇. 土壤微生物生态学及其实验技术. 北京: 科学出版社, 2006. |
| [21] | 周海霞, 周向军, 许禔森. 不同土壤利用类型对微生物的影响. 安徽农业科学, 2008, 36(15): 6415-6417. |
| [22] | 李秀英, 赵秉强, 李絮花, 李燕婷, 孙瑞莲, 朱鲁生, 徐晶, 王丽霞, 李小平, 张夫道. 不同施肥制度对土壤微生物的影响及其与土壤肥力的关系. 中国农业科学, 2005, 38(8): 1591-1599. |
| [23] | 张雷, 许艳丽, 李春杰, 韩晓增, 魏巍. 不同水肥条件对农田土壤细菌生理菌群的影响. 大豆科学, 2009, 28(2): 251-255, 265-265. |
| [24] | 米亮, 王光华, 金剑, 刘居东, 刘晓冰. 黑土微生物呼吸及群落功能多样性对温度的响应. 应用生态学报, 2010, 21(6): 1485-1491. |
| [25] | 时鹏, 高强, 王淑平, 张妍. 玉米连作及其施肥对土壤微生物群落功能多样性的影响. 生态学报, 2010, 30(22): 6173-6182. |
| [26] | 侯晓杰, 汪景宽, 李世朋. 不同施肥处理与地膜覆盖对土壤微生物群落功能多样性的影响. 生态学报, 2007, 27(2): 655-661. |
| [27] | 辜运富, 云翔, 张小平, 涂仕华, 孙锡发, Lindström K. 不同施肥处理对石灰性紫色土微生物数量及氨氧化细菌群落结构的影响. 中国农业科学, 2008, 41(12): 4119-4126. |
| [28] | Wu M N, Qin H L, Chen Z, Wu J S, Wei W X. Effect of long-term fertilization on bacterial composition in rice paddy soil. Biology and Fertility of Soils, 2011, 47(4): 397-405. |
| [29] | 徐永刚, 宇万太, 马强, 周桦. 长期不同施肥制度对潮棕壤微生物生物量碳、氮及细菌群落结构的影响. 应用生态学报, 2010, 21(8): 2078-2085. |
| [30] | 孙瑞莲, 朱鲁生, 赵秉强, 周启星, 徐晶, 张夫道. 长期施肥对土壤微生物的影响及其在养分调控中的作用. 应用生态学报, 2004, 15(10): 1907-1910. |
| [31] | Kowalchuk G A, Buma D S, de Boer W, Klinkhamer P G L, van Veen J A. Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms. Antonie van Leeuwenhoek, 2002, 81(1/4): 509-520. |
| [32] | 梁斌, 周建斌, 杨学云, 艾娜. 不同培肥措施下种植制度及撂荒对土壤微生物量碳氮的影响. 中国生态农业学报, 2009, 17(2): 209-214. |
| [33] | Saggar S, Yeates G W, Shepherd T G. Cultivation effects on soil biological properties, microfauna and organic matter dynamics in Eutric Gleysol and Gleyic Luvisol soils in New Zealand. Soil and Tillage Research, 2001, 58(1/2): 55-68. |
| [34] | 郑勇, 高勇生, 张丽梅, 何园球, 贺纪正. 长期施肥对旱地红壤微生物和酶活性的影响. 植物营养与肥料学报, 2008, 14(2): 316-321. |
| [35] | 孙凤霞, 张伟华, 徐明岗, 张文菊, 李兆强, 张敬业. 长期施肥对红壤微生物生物量碳氮和微生物碳源利用的影响. 应用生态学报, 2010, 21(11): 2792-2798. |
| [36] | 罗希茜, 郝晓晖, 陈涛, 邓婵娟, 吴金水, 胡荣桂. 长期不同施肥对稻田土壤微生物群落功能多样性的影响. 生态学报, 2009, 29(2): 740-748. |