 2015, Vol. 35
2015, Vol. 35文章信息
- 骆争荣, 陈德良, 杨辉, 苏立蕾, 丁炳扬
- LUO Zhengrong, CHEN Deliang, YANG Hui, SU Lilei, DING Bingyang
- 百山祖常绿阔叶林厚叶红淡比的萌蘖繁殖特性
- Sprouting propagation characteristics of Cleyera pachyphylla in the Baishanzu subtropical evergreen forest
- 生态学报, 2015, 35(16): 5385-5392
- Acta Ecologica Sinica, 2015, 35(16): 5385-5392
- http://dx.doi.org/10.5846/stxb201402140254
-
文章历史
- 收稿日期: 2014-02-14
- 网络出版日期: 2014-10-08






2. 浙江凤阳山-百山祖国家级自然保护区百山祖管理处, 庆元 323800;
3. 浙江农林大学林业与生物技术学院, 临安 311300;
4. 温州大学生命与环境科学学院, 温州 325035
2. Management Office of Baishanzu, Fengyangshan-Baishanzu National Nature Reserve, Qingyuan 323800, China;
3. School of Forestry and Bio-technology, Zhejiang A &F University, Lin'an 311300, China;
4. School of Life and Environmental Science, Wenzhou Univeristy, Wenzhou 325035, China
克隆植物主要是指在自然生境条件下,能够通过营养生长过程,形成多个在遗传上一致、在形态和生理上独立或者潜在独立的个体的一类植物[1],在植物界普遍存在[2, 3]。由于具有较强的适应恶劣环境能力和较强的生存竞争能力,克隆植物在许多生态系统中处于优势地位,在群落更新和恢复生态过程以及物种多样性的维持中起着重要的作用[4, 5, 6]。
自Salzman1985年在Science上发表克隆植物生境选择的开创性研究以来,国内外学者已经从形态、生理、生态特征等多方面对克隆植物进行了较深入的研究,提出了不少观点和假说[3, 4, 7, 8, 9, 10]。就针对的生态系统来说,先前的研究大多来自降雨量较少的草地生态系统或灌丛[11, 12, 13, 14, 15],关于森林生态系统的克隆植物信息较少[16, 17, 18, 19]。就选择的研究类群来说,国内外大多数学者的研究以草本克隆植物为对象,较少涉及灌木,而涉及乔木的研究则更加少见[11, 14, 18]。由于乔木物种比草本植物需要更长的生长时间才能到达性成熟,其在克隆繁殖特性的上可能会与克隆草本植物存在较大的差异,许多理论仍有待在克隆性乔木中得到验证。因此,直接在自然森林生态系统研究木本植物的克隆生态特性十分重要[20]。
虽然很多研究表明克隆植物的会降低群落的物种多样性[2, 4, 13],但由于克隆繁殖在一定程度上扩展了物种的更新生态位,它们在干扰后森林群落的恢复中起着十分重要的作用[18, 19]。物种间构件特征的差异是维持森林群落物种多样性的重要原因[21]。克隆繁殖可以提高藤本植物的多度和基面积[17],其和干扰共同决定了热带森林藤本植物的多样性[19]。许多森林优势物种都属于萌蘖型克隆乔木。这些优势克隆乔木在森林中的具体表现仍需进一步研究。
近年来国内外越来越多的森林动态监测样地为在森林中更好地研究克隆性乔木提供了良好的平台[22, 23]。2003年建立的百山祖5 hm2森林动态样地的研究发现该样地多种植物存在明显的克隆繁殖现象,其中包括几种群落优势种[24]。厚叶红淡比(Cleyera pachyphylla)是该群落中单个母株产生的萌蘖数差异最大,总萌蘖数最多的一个优势种。因此,对该森林中以厚叶红淡比为代表的克隆性乔木进行深入的调查和研究,有利于增加对此类亚热带常绿阔叶林群落动态和物种多样性维持机制的理解和认识。本研究以厚叶红淡比为对象,探究其在自然状态下的克隆构型、克隆繁殖效益以及生境对其克隆可塑性的影响。
1 研究区域概况研究地点位于我国浙江省省庆元县境内的百山祖,地处凤阳山-百山祖国家级自然保护区。该自然保护区地处中亚热带季风气候,降水充沛,光温互补,相对湿度大,海拔较高,平均温度较低。根据百山祖保护站气象观测点(海拔1100 m)1982至2001年的观测资料,该地年均气温12.8 ℃,年降水量为2,341.8 mm,相对湿度84%,10 ℃ 以上的年积温为3653.3 ℃,极端高温32 ℃,极端低温-13 ℃,无霜期187 d。保护区地处我国东南沿海的闽浙丘陵中山区,以侵蚀地貌为主,山地坡度大多在30°以上。基岩为侏罗纪火成岩,海拔800 m以下为红壤,800 m以上以黄壤为主。由于人为干扰相对较少,区内主峰——百山祖北坡中山地带现存发育较好的常绿阔叶林。
2 调查和研究方法 2.1 野外调查2002—2003年在百山祖主峰北坡的常绿阔叶林内生境相对均一,地势平缓处构建一个250 m×200 m(水平投影)的矩形样地(约119°11′46″E,27°45′35″ N,海拔约1400—1600 m)。整个样地的构建基于CTFS(Center for Tropical Forest Science)样地建设方案开展[25]。以20 m为单位在样地内划分网格(最后一列宽度为10 m),测量相邻网格点之间的高差。2003年进行首次群落调查,对样地内所有乔木和高度2 m以上或dbh(离地表面1.3 m处的直径)>1 cm的灌木和木质藤本编号,进行每木调查。测定内容包括:植物种名,dbh/基径(地面直径),坐标位置,树高,冠幅和生长状况,并对其标记挂牌。对于母株产生的萌蘖以母株为单位进行编号,并测量记录每个萌蘖的dbh (dbh<1 cm的则测量基径)、高度和生长状态。2008年夏和2013年夏,分别对样地进行了两次复查。
根据2008年复查的数据,整个样地厚叶红淡比母株总数为2383株。为进一步了解厚叶红淡比克隆繁殖特性,随机抽取至少有1个根蘖的298株(占总数的12.5%)厚叶红淡比母株作为研究对象。2011年8月详细测量并记录298株厚叶红淡比母株所拥有的萌蘖数、各萌蘖距母株的距离,萌蘖的dbh(dbh>1 cm者)或者基径(不足1.3 m或dbh<1 cm者),及萌蘖所在方位(以母株为原点,将母株所在坡面划分4个象限,自上坡左侧顺时针标记为Ⅰ、Ⅱ、Ⅲ、Ⅳ)。
2.2 研究对象本研究的对象厚叶红淡比为山茶科(Theaceae)红淡比属小乔木,高2.5—4 m,具有较强的从树干基部产生萌蘖的能力。据统计,在百山祖常绿阔叶林平均每棵厚叶红淡比母株连带2.48个存活的克隆分株,一半以上的母株(62.9%)都带有克隆分株,使其成为百山祖常绿阔叶林中克隆分株最多的木本植物[24]。部分较强壮的克隆分株倒伏贴地生长,可在离母株一定距离处产生不定根,吸收土壤养分(类似于根状茎植物)。这些克隆分株继续分株,部分厚叶红淡比基株能在森林中形成庞大无性系群体。
2.3 数据统计和分析 2.3.1 厚叶红淡比萌蘖的数量和距离分级统计各母株所含萌蘖数,进行量级划分,每隔3株归为一个级别,如1株、2株和3株归为第一级,依此类推,从少到多为1—9级。
统计各萌蘖与母株间的距离分布,进行量级划分,间隔10 cm为1级,如与母株相距0—10 cm为第一级,80 cm以下依此类推,从近到远为1—8级,大于80 cm的归为一级(第9级)。
2.3.2 萌蘖与生境的关系根据母株的空间位置确定其所在的10 m×10 m的小样方,小样方的生境类型划分采用王伟等人的研究结果,分为沟谷、陡坡、缓坡、山脊[26]。采用方差分析比较单棵母株的萌蘖数以及母株与其每个萌蘖的距离在这 4 种生境中的差异。为满足方差分析的要求,分析之前先对萌蘖数和距离进行对数转换。
为探求萌蘖分株是否存在方向性,采用成对t检验,比较了母株所在点上坡(Ⅰ、Ⅱ象限)与下坡(Ⅲ、Ⅳ象限)萌蘖数量的差异以及母株所在点左侧(Ⅱ、Ⅲ象限)与右侧(Ⅰ、Ⅳ象限)之间的差异。
2.3.3 萌蘖数与母株大小的关系采用spearman秩相关系数分析萌蘖数量与母株dbh之间的相关关系。
2.3.4 克隆群体样本和整个种群的径级结构比较分析克隆群体样本(298棵母株及其萌蘖)和样地内整个厚叶红淡比(有克隆分株的仅以dbh最大的母株作为该基株的代表,以2003年调查数据为准)种群的径级结构。径级划分为8级:①基径<0.5 cm,②基径≥0.5 cm且dbh<1 cm,③1 cm≤dbh<4 cm,④4 cm≤dbh<7 cm,⑤7 cm≤dbh<10 cm,⑥10 cm≤dbh<13 cm,⑦13 cm≤dbh<16 cm,⑧dbh≥16 cm。采用卡方检验比较克隆群体样本和整个种群径级结构的差异。
2.3.5 萌蘖数量对母株存活率的影响为了解基株内部的生理整合对母株生存的影响,分析了母株所带萌蘖数与母株在2011年8月至2013年8月间的存活率的关系。由于母株存活与否(存活y=1,死亡y=0)为二元变量,这里采用标准逻辑斯蒂回归模型进行分析,形式如下:
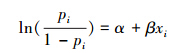
所有分析均在R统计软件中完成。
3 结果 3.1 厚叶红淡比萌蘖的数量和母株—萌蘖距离分布调查的298株厚叶红淡比基株所萌发的萌蘖数存在较大差异。在萌蘖最多的一个基株上发生了27株根蘖。母株的数量随着其萌蘖数量的增加而逐渐减少,母株所带萌蘖数量的分布结构呈明显的倒J型(图 1)。总体上每母株带1—3棵根蘖的比例最高,到4—6和7—9棵急速下降,之后缓慢下降。

|
| 图 1 厚叶红淡比根蘖数量和母株—萌蘖距离的分布结构 Fig.1 Distribution of the number of sprouting per genet and the distance between origin ramet and sprouting of Cleyera pachyphylla |
厚叶红淡比萌蘖大多生长于母株周围(图 1)。多数萌蘖离母株较近,萌蘖在距母株30 cm以内的占所有萌蘖的82.79%,少数萌蘖距离母株较远,距离母株最远的一株萌蘖达到163 cm。
3.2 萌蘖与生境及母株大小的关系在沟谷、陡坡、缓坡、山脊4类生境中,缓坡中的厚叶红淡比平均萌蘖数最低(3.89),萌蘖距母株平均距离为17.21 cm;陡坡中的平均萌蘖数最高(5.92),萌蘖距母株平均距离为15.73 cm。沟谷与山脊中的厚叶红淡比平均萌蘖数分别为4.73和4.62,萌蘖距母株平均距离较高,分别为20.16 cm和20.44 cm(表 1)。然而,方差分析表明单棵厚叶红淡比的萌蘖数量(F=2.11,P= 0.0988)和母株与其所有萌蘖的平均距离(F=1.78,P=0.1502)在这4 种生境中的差异不显著。
|
生境类别 Habitat | 总母株数 No. of genet | 总萌蘖数 No. of sprouting | 单位萌蘖数(±sd)* No. of sprouting per genet | 萌蘖与母株距离(±sd)/cm Average distance between origin and sprouting |
| *x±sd表示均值±标准差 | ||||
| 沟谷 Gully | 26 | 123 | 4.73±4.00 | 20.16±13.89 |
| 陡坡Steep slope | 12 | 71 | 5.92±3.58 | 15.73±7.47 |
| 缓坡Mild slope | 163 | 596 | 3.89±3.57 | 17.21±10.31 |
| 山脊 Ridge | 97 | 448 | 4.62±4.85 | 20.44±8.93 |
针对不同方位萌蘖数量的统计表明,单棵母株上坡的平均萌蘖数为1.87,下坡平均萌蘖数为2.09上下坡比例为0.89;而母株左侧平均萌蘖数为2.14,右侧平均萌蘖数为1.83,左右坡比例为1.16。成对t检验的结果显示上下坡的萌蘖数差异不显著(P =0.1187),而左右坡的萌蘖数差异显著(P=0.0379)。
对母株dbh与其萌蘖数量的相关性分析表明,萌蘖数量与母株的大小呈显著正相关(ρ=0.45,P<0.001),母株萌蘖数随着径级的增大呈现出增大的趋势。
3.3 克隆群体样本和整个种群的径级结构厚叶红淡比dbh小于1 cm的萌蘖数量丰富。就dbh≥1 cm的厚叶红淡比萌蘖而言,萌蘖数量所占的比例随着胸径的增大基本呈减少趋势。这一趋势与样地内整个厚叶红淡比实生树种群的径级分布的变化趋势基本一致(图 2)。但是,克隆群体样本和整个种群的径级分布存在显著差异(χ2=575.85,df=7,P< 0.001)。进一步分析两个径级分布的差异,主要表现在萌蘖形成的幼苗和小幼树(dbh<1 cm)的占比相对于实生的幼苗和幼树的占比要大得多,而dbh在1 cm以上的小树和成年树占比关系正好相反。在克隆群体样本中,萌蘖数量从第②径级到第③径级快速下降,而实生树数量第②径级到第③径级略有上升。

|
| 图 2 厚叶红淡比萌蘖和实生树的径级分布结构 Fig.2 Size distribution of clonal trees and seedling trees of Cleyera pachyphylla “{}”内外数字分别表示基径和胸径值,“[”表示包含本数,而“)”表示不包含本数 Values with and without“{}”indicate diameter on the ground and dbh (diameter at 1.3 m breast height),respectively; “[” indicates the size is included,while “)” indicates the size is not included |
逻辑斯蒂回归模型分析表明,母株上所萌发的萌蘖数量对母株本身在2011年8月至2013年8月的存活率没有显著影响(偏差D=2.8,P=0.141)。
4 讨论本研究发现厚叶红淡比的萌蘖大多在距母株30cm的范围内。每棵母株拥有的萌蘖数多为1—3棵,但是个体间差异较大,在野外观察到的最大的厚叶红淡比无性系基株含有94个分株。综合来看,就样地内整个厚叶红淡比种群的萌蘖率而言,每棵厚叶红淡比平均拥有2.48个萌蘖[24]。神农架地区米心水青冈(Fagus engleriana)萌枝数量的调查发现其平均萌枝数在不同生境间差异极显著,范围约在1—15之间[27]。与之相比,厚叶红淡比的萌蘖数量处在中下水平。克隆植物构型主要由间隔物的长度、分枝强度和分枝角度等3个因素决定[13]。百山祖常绿阔叶林中的厚叶红淡比分株与母株距离较短,其克隆种群更偏向于密集型构型。这种构型能够较好地利用局部的资源,并提高竞争排斥其他植物种类侵入的能力[10]。这种策略保证了在森林动态演替过程中,厚叶红淡比已经占据空间较难被其他物种入侵和替代。
由于克隆生殖较有性生殖更容易,繁殖成本更低,在环境改善时克隆植物可以通过克隆分株快速占领空间[3, 27]。但在亚热带常绿阔叶林下,植物所接受的光是有限的,而水分和养分相对较充足。在这种环境条件下,密集型克隆乔木的无性繁殖可能是低效的。本研究结果表明,虽然厚叶红淡比母株产生了大量幼苗(第①径级)和小幼树(第②径级),但是进入第③径级的幼树相对较少。大多数萌蘖幼苗和小幼树在林下处于被抑制状态,只有在出现林窗时其生长才能被释放[27]。在漫长的抑制状态中,萌蘖幼苗需要面对多种死亡风险,其中之一就是密度制约[22, 25]。虽然克隆后代由于母体的供养更容易安全度过幼龄期,但是克隆生长所带来的弊端也很大。由于克隆后代之间的遗传同一性和生理连接使得疾病更易传播[28];克隆后代扩散范围有限,往往集中在母株周围,造成激烈的同胞竞争[1]。这就使克隆植物幼苗/幼树期所经历的密度制约效应比非克隆植物更加严重[29]。先前的研究表明在幼苗期和幼苗-幼树转换期厚叶红淡比的个体存活和空间格局受到明显的密度制约效应的影响[30]。在这种效应的影响下,在幼苗和幼树期的厚叶红淡比萌蘖比实生苗的死亡率要高得多。在这种不利因素的影响下,森林中的具有强克隆繁殖能力乔木树种的实际克隆繁殖效率并不高。例如,青冈(Cyclobalanopsis glauca)也具有极强的萌蘖能力,但是由于受到包括密度制约在内的不利因素的影响,我国华东地区青冈每个基因型含有的克隆分株仅为1.508[31]。这说明,虽然像青冈这样的克隆性乔木能产生大量萌蘖,但是真正成功的克隆繁殖(形成独立新个体)并不多。通过克隆繁殖,青冈仅能使个体(基因型)增殖1.508倍。 这一机制能够较好的解释常绿阔叶林中,很多克隆植物都是群落的优势种,但是没有一个克隆植物能形成独霸生存空间的现象。这有利于森林物种多样性的维持。
厚叶红淡比克隆形态可塑性与其本身的遗传特性有关,同时与环境条件也有密切的关系。适应假说认为克隆植物的形态可塑性是与生境的资源状况相适应的,并预测来自资源水平高且资源斑块性稳定生境的克隆植物比来自资源水平低且资源斑块性不稳定生境的具有更高的形态可塑性[32]。山地森林是异质性较大的系统,因为地形的变化可以引起一系列的水分和土壤养分的异质性。克隆植物常以形态上的可塑性来整合这些呈异质性分布的资源。贺金生等在神农架地区对米心水青冈的克隆繁殖的调查发现不同样地之间米心水青冈的萌蘖强度差异显著[27]。本研究分析了单棵厚叶红淡比的萌蘖数量在依地形差异划分的各类生境中的变化,结果表明不同生境中厚叶红淡比的萌蘖数量差异不显著。由于本样地在建设时就有目的地选择了地形比较均一的森林斑块,地形差异较小,水分和养分的异质性也较小,因此厚叶红淡比的萌蘖数量没有在不同地形生境中体现出差异。然而,从厚叶红淡比萌蘖数量的空间分布上看,即使在这样一个地形和土壤相对均质的环境中,厚叶红淡比萌蘖强度依然表现出了较大的可塑性。在林下环境,光照的异质性对木本植物的克隆强度可塑性具有更为重要的影响。贺金生等人对神农架米心水青冈萌枝过程的研究表明干扰后林窗的形成期是米心水青冈萌枝产生的高峰期[27]。本研究结果也显示,位于母株左侧(东侧)的萌蘖数量显著大于右侧(西侧)的。由于研究样地位于百山祖主峰北坡,朝向略偏东,因此东侧树冠往往能够获得更好的光照。已有的研究表明,克隆植物的生理整合常常具有一定的格局和范围,位于同一侧的植物构件常具有直接的维管联系,而相对侧的构件间维管系统相对独立[33]。因此东侧的树冠合成的有机物将更多地往东侧根部输送,以支持更多地萌蘖。光资源的异质性可能是厚叶红淡比萌蘖强度表现出差异的重要原因。另外,本研究的结果显示母株萌蘖数量与母株的大小呈显著正相关。这表明对于个体较小的母株,母株本身从环境中竞争获得光、肥和水的能力有限,同化的营养基本用来促进自身迅速的生长,少有剩余的能量和物质用以无性繁殖,导致较小的厚叶红淡比母株萌蘖率低。而当母株生长到一定阶段时,生长状态良好的厚叶红淡比母株能不断产生新的且具有潜在的独立生长能力的萌蘖来巩固自身的生存空间。母株大小的差异是其萌蘖强度可塑性的重要原因。
由于克隆分株的生长需要母株在物质和能量上提供支持,对资源缺乏的分株的资源供给,可能会降低资源供应分株的适合度[34],而过多的萌蘖可能会降低母株的存活率。然而,本研究发现在常绿阔叶林中,较多的萌蘖不会降低厚叶红淡比母株短期的存活率。这说明,厚叶红淡比可能具有自我调节萌蘖数量的能力,单棵母株萌蘖数量的增加与之可提供的能量相适应,不会以提高母株本身的死亡风险为代价。萌蘖数量较多的往往是个体较大、树龄较长的厚叶红淡比个体。这些个体对资源的获取能力更强,抗风险能力更强。在这种情况下,增加萌蘖数量并不会提高母株的死亡风险。经过长期的进化,厚叶红淡比对萌蘖数量具备了较好自我控制能力。
5 结论通过自我调节控制萌蘖数量,厚叶红淡比萌蘖数量的增加不以提高母株本身的死亡风险为代价。同时,利用密集型的克隆构型,厚叶红淡比可以充分利用局部资源,使其成为百山祖常绿阔叶林群落的优势种之一。但是由于存在密度制约效应等负面作用的影响,厚叶红淡比的克隆繁殖成功率较低,使其不能独霸生存空间,有利于森林物种多样性的维持。因此,克隆性乔木的优势和弊端共同决定了其在森林中的适合度,也决定了其在森林中的地位。
本研究没有在不同生境中检测到厚叶红淡比显著的形态可塑性,原因在于本研究主要根据地形差异进行生境分类,分析仍然较粗。本研究间接显示厚叶红淡比萌蘖对光资源的异质性具有反应,存在一定的向光性。因此,今后需要详细测定厚叶红淡比生长环境的土壤养分和光照水平,进一步探求其在异质性生境中的生理整合和形态可塑性。萌蘖繁殖的向光性也说明:与热带森林中的藤本植物类似,适度干扰(林窗)可能有利于提高亚热带森林中克隆性乔木的多度和多样性。
致谢:感谢王伟、蒋清玲、潘霞、顾莎莎等同学和百山祖保护站护林员吴天明、傅明珠和吴义松在野外作业时的帮助。| [1] | de Kroon H, Van Groenendael J. The Ecology and Evolution of Clonal Plants. The Netherlands: Backhuys Publishers, 1997. |
| [2] | 宋明华, 董鸣. 群落中克隆植物的重要性. 生态学报, 2002, 22(11): 1960-1967. |
| [3] | 王洪义, 王正文, 李凌浩, 陈雅君, 任丽昀. 不同生境中克隆植物的繁殖倾向. 生态学杂志, 2005, 24(6): 670-676. |
| [4] | Callaghan T V, Carlsson B A, Jonsdottir I S, Svensson B M, Jonasson S. Clonal plants and environmental change: introduction to the proceedings and summary. Oikos, 1992, 63(3): 341-347. |
| [5] | Prach K, Pysek P. Clonal plants-what is their role in succession. Folia Geobotanica et Phytotaxonomica, 1994, 29(2): 307-320. |
| [6] | Oborny B, Bartha S. Clonality in plant communities-an overview. Abstracta Botanuca, 1995, 19: 115-127. |
| [7] | Salzman A G. Habitat selection in a clonal plant. Science, 1985, 228(4699): 603-604. |
| [8] | 董鸣. 资源异质性环境中的植物克隆生长: 觅食行为. 植物学报, 1996, 38(10): 828-835. |
| [9] | 何成新, 李先琨, 唐润琴, 苏宗明. 克隆植物种群生态学研究透视. 广西植物, 2001, 21(3): 264-272. |
| [10] | 汤俊兵, 肖燕, 安树青. 根茎克隆植物生态学研究进展. 生态学报, 2010, 30(11): 3028-3036. |
| [11] | Yu F H, Dong M, Krusi B. Clonal integration helps Psammochloa villosa survive sand burial in an inland dune. New Phytologist, 2004, 162(3): 697-704. |
| [12] | Yu F H, Wang N, He W M, Chu Y, Dong M. Adaptation of rhizome connections in drylands: increasing tolerance of clones to wind erosion. Annals of Botany, 2008, 102(4): 571-577. |
| [13] | Eilts J A, Mittelbach G G, Reynolds H L, Gross K L. Resource heterogeneity, soil fertility, and species diversity: effects of clonal species on plant communities. The American Naturalist, 2011, 177(5): 574-588. |
| [14] | Xu L, Yu F H, van Drunen E, Schieving F, Dong M, Anten N P. Trampling, defoliation and physiological integration affect growth, morphological and mechanical properties of a root-suckering clonal tree. Annals of Botany, 2013, 109(5): 1001-1008. |
| [15] | Pinno B D, Wilson S D. Nitrogen translocation between clonal mother and daughter trees at a grassland-forest boundary. Plant Ecology, 2014, 215(3): 347-354. |
| [16] | Du J, Wang N, Alpert P, Yu M J, Yu F H, Dong M. Clonal integration increases performance of ramets of the fern Diplopterygium glaucum in an evergreen forest in southeastern China. Flora, 2010, 205(6): 399-403. |
| [17] | Yorke S R, Schnitzer S A, Mascaro J, Letcher S G, Carson W P. Increasing liana abundance and basal area in a tropical forest: the contribution of long-distance clonal colonization. Biotropica, 2013, 45(3): 317-324. |
| [18] | Escandon A B, Paula S, Rojas R, Corcuera L J, Coopman R E. Sprouting extends the regeneration niche in temperate rain forests: the case of the long-lived tree Eucryphia cordifolia. Forest Ecology and Management, 2013, 310: 321-326. |
| [19] | Ledo A, Schnitzer S A. Distribution and clonal reproduction determine liana distribution and maintain liana diversity in a tropical forest. Ecology, 2014. |
| [20] | 董鸣, 于飞海, 安树青, 何维明, 梁士楚. 植物克隆性的生态学意义. 植物生态学报, 2007, 31(4): 549-551. |
| [21] | Wildova R, Goldberg D E, Herben T. The Contrasting roles of growth traits and architectural traits in diversity maintenance in clonal plant communities. The American Naturalist, 2012, 180(6): 693-697. |
| [22] | Condit R. Research in large, long-term tropical forest plots. Trends in Ecology and Evolution, 1995, 10(1): 18-22. |
| [23] | 马克平. 监测是评估生物多样性保护进展的有效途径. 生物多样性, 2011, 19(2): 125-126. |
| [24] | 陈娇, 王伟, 叶珍林, 陈晓慧, 蒋清玲, 丁炳扬. 百山祖常绿阔叶林木本植物的克隆繁殖特性. 浙江大学学报: 农业与生命科学版, 2012, 38(6): 684-692. |
| [25] | Condit R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots. Berlin: Springer-Verlag, 1998: 50-56. |
| [26] | 王伟, 骆争荣, 周荣飞, 许大明, 哀建国, 丁炳扬. 百山祖常绿阔叶林木本植物的生境相关性分析. 生物多样性, 2011, 19(2): 134-142. |
| [27] | 贺金生, 陈伟烈, 刘峰. 神农架地区米心水青冈萌枝过程的研究. 植物生态学报, 1998, 22(5): 385-391. |
| [28] | Pitelka L F, Ashmun J W. Physiology and integration of ramets in clonal plants//Jackson J B C, Buss L W. Cook R E. Population Biology and Evolution of Clonal Organisms. New Haven: Yale University Press, 1985: 399-436. |
| [29] | Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalist, 1970, 104(940): 501-528. |
| [30] | Luo Z R, Mi X C, Chen X R, Ye Z L, Ding B Y. Density dependence is not very prevalent in a heterogeneous subtropical forest. Oikos, 2012, 121(8): 1239-1250. |
| [31] | 陈小勇, 宋勇昌. 青冈种群克隆多样性及其与环境因子的关系. 植物生态学报, 1997, 21(4): 342-348. |
| [32] | Dong M. Foraging Through Morphological Responses in Clonal Herbs [D]. Utrecht: Utrecht University, 1994. |
| [33] | Price E A C, Hutchings M J. Studies of growth in the clonal herb Glechoma hederacea. Ⅱ. The effect s of selective defoliation. Journal of Ecology, 1992, 80(1): 39-47. |
| [34] | Pitelka L F, Hansen S B, Ashmun J W. Population biology of Clintonia borealis. I. Ramet and patch dynamics. Journal of Ecology, 1985, 73(1): 169-183. |