 2015, Vol. 35
2015, Vol. 35文章信息
- 刘宏伟, 刘文丹, 王微, 柴捷, 陶建平
- LIU Hongwei, LIU Wendan, WANG Wei, CHAI Jie, TAO Jianping
- 重庆石灰岩地区主要木本植物叶片性状及养分再吸收特征
- Leaf traits and nutrient resorption of major woody species in the karst limestone area of Chongqing
- 生态学报, 2015, 35(12): 4071-4080
- Acta Ecologica Sinica, 2015, 35(12): 4071-4080
- http://dx.doi.org/10.5846/stxb201310262584
-
文章历史
- 收稿日期:2013-10-26
- 网络出版日期:2014-08-28






2. 西南大学生命科学学院, 重庆 400715
2. School of Life Science, Southwest University, Chongqing 400715, China
养分再吸收(Nutrient resorption)是指养分从衰老叶片中转移并被运输到植物其他组织供其重复利用的过程。养分再吸收不仅影响植物体内养分的有效性和养分循环过程,而且也是生物地球化学过程的一个关键步骤[1, 2, 3] 。这个过程控制了养分从衰老植物组织中的移动和回收,以及这些养分向多年生储存器官的运输[1, 4] 。养分再吸收是植物对养分贫瘠环境的一种适应机制[5, 6, 7],也是植物保存养分、增强竞争力、提高养分利用效率和生产力的重要策略[8, 9, 10],研究陆地多年生植物对不同土壤养分有效性的适应,特别是植物对贫瘠环境的适应性,需要考虑从植物的养分再吸收入手。
早在20世纪20年代,科学家就已经注意到在叶片衰老过程中就存在着养分再吸收的现象,到目前仍有大量养分再吸收的研究报道[3, 11]。由于N和P是限制陆地生态系统植物生长的重要营养元素[12, 13],因此关于它们利用效率的研究引起了国内外学者的高度关注。但是,这些研究大都集中在对不同物种或同种物种不同年龄阶段的叶片养分再吸收效率的比较上。国内对养分再吸收效率的研究始于20世纪90年代末,除一些学者介绍国外相关研究进展外[14, 15],研究工作主要集中在樟子松(Pinus sylvestris var. mongolica)、冬麦(Ophiopogon japonica)、马尾松(Pinus massoniana Lamb)、文冠果(Xanthoceras sorbifolia Bunge)、短枝木麻黄(Casuarina equisetifolia)和兴安落叶松(Larix gmelinii)等少数树种上[8, 9, 10, 16, 17, 18],而且都着重于对植物叶片养分再吸收效率现象的研究,而对石灰岩喀斯特地区不同功能群植物养分再吸收效率、叶片功能性状以及它们之间的关系研究较少。
我国西南喀斯特地区面积达50多万km2,是全球喀斯特集中分布区面积最大、岩溶发育最强烈、景观类型复杂、生态系统极为脆弱的典型地区[19] 。喀斯特石灰岩地区营养条件贫瘠,生长在这种立地条件下的植物必有其有效的适应机制,而不同生活型植物对该立地条件具有不同的适应特征,落叶植物具有较粗的导管、较高的木质部比导率和边材比导率,常绿植物的水力导度较低、抵抗空穴化能力较强、水分运输安全性较高、但长期水分利用效率较低[20, 21, 22]。本文试图从养分再吸收的角度对两类植物进行探讨,选取了喀斯特石灰岩地区15种常绿和14种落叶木本植物作为研究对象,比较了两类植物叶片衰老前后叶性状以及养分再吸收效率的差异,旨在了解石灰岩地区常绿和落叶两类不同生活型植物养分保存机制,以为当地植被恢复提供科学依据。
1 材料与方法 1.1 试验地概况试验地设在重庆市中梁山海石公园(106°18′E,29°39′N)处于长江中上游防护绿化带,属于亚热带湿润季风气候。夏季高温多雨,冬季温暖雾多,相对湿度大,降雨集中,年均气温为16.8 ℃,年均降雨量为1000—1300 mm。土壤为山地黄壤和山地黄棕壤,土层浅薄,保水保肥性差,土体不连续,岩石裸露率高,土壤富钙而偏碱性,且总量少[23, 24] 。植被主要以火棘(Pyracantha fortuneana)、南天竺(Nandina domestica)、铁仔(Myrsine Africana L.)、湖北羊蹄甲(Bauhinia purpurea L.)等为优势种的次生灌丛[25]。
1.2 物种选择与取样方法研究样地位于重庆市海石公园园内,地貌特征为相对平坦的石灰岩山地,在立地条件相似的混交林中设置一块大小为500m×500 m的样地,在样地中分别选择15个常绿木本植物种和14个落叶木本植物种进行叶片性状的测定和养分再吸收测定(表 1)。对于选定的每种植物,分别在样地内标记5株生长良好、大小一致、没有遮阴的个体作为取样植株。
| 生活型Life form | 编号Number | 物种Species | 科Family |
| 常绿Evergreen | 1 | 湖北羊蹄甲Bauhinia purpurea | 豆科Leguminosae |
| 2 | 常春油麻藤Mucuna sempervirens | 豆科Leguminosae | |
| 3 | 香花崖豆藤Millettia dielsiana | 豆科Leguminosae | |
| 4 | 金樱子Rosa laevigata | 蔷薇科Rosaceae | |
| 5 | 火棘Pyracantha fortuneana | 蔷薇科Rosaceae | |
| 6 | 大叶樟Cinnamomum septentrionale | 樟科Lauraceae | |
| 7 | 天竺桂Cinnamomum pedunculatum | 樟科Lauraceae | |
| 8 | 小蜡Ligustrum sinense | 木犀科Oleaceae | |
| 9 | 女贞Ligustrum lucidum | 木犀科Oleaceae | |
| 10 | 南天竺Nandina domestica | 小檗科Berberidaceae | |
| 11 | 铁仔Myrsine africana | 紫金牛科Myrsinaceae | |
| 12 | 榕叶冬青Lex ficoidea | 冬青科Aquifoliaceae | |
| 13 | 乌柿Diospyros cathayensis | 柿科Ebenaceae | |
| 14 | 金山荚蒾Viburnum Chinshanense | 忍冬科Caprifoliaceae | |
| 15 | 山杜英Elaeocarpus sylvestris | 杜英科Elaeocarpaceae | |
| 落叶Deciduous | 16 | 算盘子Glochidion puberum | 大戟科Euphorbiaceae |
| 17 | 乌桕Sapium sebiferum | 大戟科Euphorbiaceae | |
| 18 | 石岩枫Mallotus repandus | 大戟科Euphorbiaceae | |
| 19 | 华南云实Caesalpinia crista | 豆科Leguminosae | |
| 20 | 云实Caesalpinia decapetala | 豆科Leguminosae | |
| 21 | 山合欢Albizia kalkora | 豆科Leguminosae | |
| 22 | 牧荆Vitex negundo | 马鞭草科Verbenaceae | |
| 23 | 盐肤木Rhus chinensis | 漆树科Anacardiaceae | |
| 24 | 悬钩子Rubus corchorifolius | 蔷薇科Rosaceae | |
| 25 | 朴树Celtis sinensis | 榆科Ulmaceae | |
| 26 | 黄葛树Ficus virens | 桑科Moraceae | |
| 27 | 柿子Diospyros | 柿科Ebenaceae | |
| 28 | 勾儿茶Berchemia racemosa | 鼠李Rhamnaceae | |
| 29 | 异叶八角枫Rhodotypos scandens | 八角枫科Alangiaceae |
于2012年7月下旬至8月中旬采集植物成熟叶片,考虑到在不同高度树木的叶片养分含量会有差异,本研究选取树冠中部的叶片,以代表整个树冠层的叶片。取样时,每株植物选择完全展开、没有病虫害且向阳一侧的健康成熟叶片(绿色)进行采集。采集叶片数目因具体物种和叶片大小而定,对于叶片较大的植物,每株个体采集3—5片完全展开的健康样品用于叶干物质含量、比叶面积及叶片厚度的测定,另外采集3—5片完全展开的健康样品用于养分含量的测定,对于小叶植物,每个个体采集8—10个叶片。用剪刀剪下叶片,然后选择与采集叶片对生或邻近的叶片,挂牌标记并与已采集的健康叶片一一对应,作为衰老叶片的采集对象。将采集的健康成熟叶片置于两片湿润的滤纸之间,放入自封袋内,然后储藏在黑暗的冰袋内(底部有冰袋,内部温度<5 ℃),带回实验室待测[26]。
2012年9月底开始采集植物衰老叶片,以后每周采集两次,直到2013年3月底样片全部采集完,取样时,当标记的叶片出现明显的衰老特征(个别除外,大部分变为黄色或红色)时进行采集,或者用手轻轻一震被标记叶片所在的枝条,叶片脱落,即可判断此时叶片已经衰老,收集落叶,将叶片置于两片湿润的滤纸之间,放入自封袋内,然后储藏在黑暗的冰包内(底部有冰袋,内部温度<5 ℃),带回实验室待测。
1.3 叶性状因子的测定测定叶干物质含量(LDMC)、比叶面积(SLA)、叶片厚度(LT)3个结构性状和叶氮含量(LNC)、叶磷含量(LPC)和叶氮磷比(N ∶ P)3个养分性状。
1.3.1 叶片结构性状的测定将叶片放入水中,在5 ℃的黑暗环境中储藏12 h,取出后迅速用吸水纸吸去叶片表面的水分,在万分之一的电子天平上称重(饱和鲜重),然后将叶片用扫描仪扫描,利用Delta-T叶面积仪(Cambridge,UK)测定其面积(LA),用精度为0.02 mm的游标卡尺测量叶片同侧上、中、下3个厚度(避开主叶脉),取平均值即为单叶LT,最后将叶片于105 ℃下杀青15 min,75 ℃下烘干48 h至恒重。LDMC)和SLA的计算公式为:LDMC=叶片干重(g)/叶片饱和鲜重(kg),SLA=叶片面积(m2)/叶片干重(kg)[27]。
1.3.2 叶片养分性状的测定叶片全氮含量采用CHNS-O元素分析仪(Vario EL cube,Germany)测定;叶片全磷含量采用酸溶-钼锑抗比色法测定。LNC(mg/g)=叶片全氮(mg)/叶片干重(g);LPC=叶片全磷(mg)/叶片干重(g)[26, 28] 。
1.4 叶片养分再吸收效率和再吸收程度的计算养分再吸收大小通常用养分再吸收效率(NRE)和养分再吸收程度(NRP)来表示,前者是指叶片在衰老过程中再吸收养分量与叶片最高养分库含量之比,或成熟叶和衰老叶之间养分减少的百分率,而后者则直接用衰老叶中的养分含量来表示。本文分别基于单位质量叶片养分含量(mg/g,RE1)和单位面积叶片养分含量(μg/cm2,RE2)对计算养分再吸收效率,其中,计算公式如下:
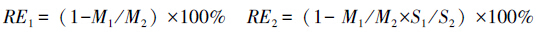
式中,M1和M2分别代表衰老叶片和成熟叶片元素的质量浓度(mg/g);S1和S2分别代表衰老叶片和成熟叶片单位叶面积的质量(μg/cm2)。
养分元素再吸收程度(RP)用衰老叶片中养分元素含量来表示,衰老叶片中的养分含量元素含量越低表示再吸收程度越高[29] 。
1.5 数据统计分析采用独立样本T检验(Independent T-Test)比较常绿和落叶两种不同生活型植物成熟和衰老叶片两个时期各叶性因子、养分含量及养分再吸收效率之间的差异,采用Pearson相关系数检验法分析N、P再吸收效率与其他叶性因子之间的相关性。数据统计结果表示为平均值±标准误(Mean±SE),所有分析过程均在SPSS for Windows(Version19.0)软件下进行。
2 结果与分析 2.1 叶片衰老前后常绿和落叶植物叶片结构特征之间差异所有植物叶片在衰老过程中LDMC表现出下降的趋势,但SLA和LT变化不明显(图 1)。其中,在成熟叶中,常绿植物LDMC和LT均要显著高于落叶植物,而SLA则显著低于落叶植物(P<0.05),常绿和落叶植物成熟叶平均LDMC和LT分别为(419.97±13.91) g/kg、(0.18±0.01) mm和(366.85±16.57) g/kg、(0.14±0.01) mm,SLA分别是(11.64±1.08) m2/kg和(16.09±1.41) m2/kg;在衰老叶中,常绿植物SLA显著低于落叶植物,LT显著高于落叶植物(P<0.05),而LDMC在两类植物中无显著差异(P>0.05),常绿和落叶植物衰老叶平均SLA和LT分别为(12.05±0.09) m2/kg、(0.18±0.02) mm和(16.68±1.37) m2/kg、(0.14±0.01) mm,LDMC则分别为(368.45±15.29) g/kg和(333.23±19.96) g/kg。

|
| 图1 叶片衰老前后常绿和落叶植物叶性因子比较 Fig.1 Foliar characteristics before and after senescing in evergreen and deciduous species 不同字母的数值间差异显著(P<0.05) |
实验结果显示(图 2),基于单位质量叶片计算的养分含量,落叶植物成熟叶中N、P含量(N为(19.49±1.16) mg/g,P为(1.02±0.10) mg/g)略高于常绿植物(N为(18.15±1.15) mg/g,P为(0.90±0.07) mg/g),但无显著差异(P>0.05),氮磷比在两者之中差异不大(常绿植物为(21.38±1.51),落叶植物为(20.63±1.63));落叶植物衰老叶中LNC((14.79±1.34) mg/g)显著高于常绿植物LNC((11.01±1.19) mg/g)(P<0.05),与成熟叶相比,衰老叶中氮磷比表现出相反的趋势,落叶植物叶片N ∶ P(24.07±2.03)高于常绿植物叶片N ∶ P(23.55±2.42)。

|
| 图2 叶片衰老前后常绿和落叶植物养分含量比较 (基于单位质量叶片养分含量) Fig.2 Nutrient contents in mature and senesced foliar of evergreen and deciduous species (nutrient content based on per mass) |
根据RE2计算的N、P含量与RE1相比,则表现出相反的趋势(图 3),常绿植物成熟和衰老叶片LNC和LPC均要高于落叶植物,但无显著差异(P>0.05),其中,常绿植物成熟和衰老叶片中LNC和LPC分别为(171.37±15.94) μg/cm2、(102.01±12.31) μg/cm2和(8.23±1.23) μg/cm2、(5.04±0.77) μg/cm2,落叶植物的则为(134.90±12.41) μg/cm2、(95.53±9.29) μg/cm2和(7.10±0.94) μg/cm2、(4.61±0.76) μg/cm2;而用两种方法计算的叶片N ∶ P在两类植物中则表现出相似的规律,均表现为常绿植物成熟叶片N ∶ P高于落叶植物,衰老叶片N ∶ P则低于落叶植物。
2.3 基于不同方法计算的养分再吸收效率和养分再吸收程度基于不同方法计算的养分再吸收效率表现出相似的规律(表 2),常绿植物N、P元素养分再吸收效率均高于落叶植物,根据RE1计算的养分再吸收效率,常绿和落叶植物N、P再吸收效率分别为39.42%、43.79%和24.08%、33.59%,而根据RE2计算的养分再吸收效率,常绿和落叶植物N、P再吸收效率分别为39.32%、45.19%和29.59%、38.45%;对于养分再吸收程度,实验结果表明(表 3),常绿植物衰老叶片中N含量(11.01 mg/g)显著低于落叶植物(14.79 mg/g,P<0.05),P含量(0.51 mg/g)虽然也低于落叶植物(0.68 mg/g),但无显著差异(P>0.05),因此可见,常绿和落叶植物N、P再吸收程度是不同的。
| 方法 Method | 元素 Element | 生活型 Life form | |
| 常绿植物 Evergreen species | 落叶植物 Evergreen species | ||
| 同行标有不同字母的数值间差异显著(P<0.05) | |||
| 基于单位质量叶片养分含量Nutrient content based on mass(mg/g) | N | 39.42±5.02a | 24.08±4.44b |
| P | 43.79±4.67a | 33.59±4.86a | |
| 基于单位面积叶片养分含量Nutrient content based on area(μg/cm) | N | 39.32±6.03a | 29.59±5.12a |
| P | 45.19±5.13a | 38.45±5.78a | |
| 方法 Method | 元素 Element | 生活型 Life form | |
| 常绿植物 Evergreen species | 落叶植物 Evergreen species | ||
| 同行标有不同字母的数值间差异显著(P<0.05) | |||
| 基于单位质量叶片养分含量Nutrient content based on mass/(mg/g) | N | 11.01±1.19a | 14.79±1.34b |
| P | 0.51±0.07a | 0.68±0.08a | |

|
| 图3 叶片衰老前后常绿和落叶植物养分含量比较 (基于单位面积叶片养分含量) Fig.3 Nutrient contents in mature and senesced foliar of evergreen and deciduous species (nutrient content based on per mass) |
根据RE1和RE2计算的养分再吸收效率基本相似,这里仅基于单位质量叶片养分含量计算的养分再吸收效率与各叶性状因子之间建立相关性,常绿和落叶植物N、P再吸收效率与成熟叶片各叶性状因子之间的相关性如表 4所示,结果表明两类植物N、P再吸收效率与成熟叶片中LDMC、SLA、LPC、LNC、LPC及N ∶ P之间均不存在显著相关性(P>0.05);两类植物N、P再吸收效率与衰老叶片各叶性状因子之间的相关性如表 5所示,N、P再吸收效率与LDMC、SLA、LT及N ∶ P之间没有显著相关性(P>0.05),而与衰老叶片中的养分含量存在显著负相关(P<0.05),常绿植物N再吸收效率与衰老叶中P含量存在显著负相关(r=-0.602),落叶植物P再吸收效率与衰老叶中N含量也存在显著负相关(r=-0.776),这表明植物衰老叶中养分含量越低,N、P的再吸收效率越高。
3 讨论在植物对环境长期的适应进化过程中,为了适应不同的立地条件,不同功能群的植物具有各自的养分再吸收效率。Aerts[2]通过对不同种植物的养分再吸收研究认为:养分再吸收效率在不同生活型之间差异不大,常绿植物N的再吸收效率为47%,落叶植物的为54%,P的再吸收效率在这两种生活型之间没有明显差异(分别为51%和50%)。Leonardus等人[3]在全球尺度上对多种陆生植物的养分再吸收效率和叶片中的养分含量研究表明:N、P具有较高的养分再吸收效率(分别为62.1%、64.9%)。本研究与上述结论存在差异,对于生长在喀斯特石灰岩地区的植物,无论是常绿还是落叶植物,N、P再吸收效率都低于上述研究结果,其中,常绿植物N、P再吸收效率为39.42%、43.79%,落叶植物的是24.08%、33.59%,这可能一方面由于不同的研究选取的物种不同导致的,说明养分再吸收存在种间差异,另一方面也可能由于立地条件对再吸收效率具有显著影响,喀斯特石灰岩地区作为一种特殊的立地条件,植物养分再吸收效率表现出了特异性。
| 叶片性状 Leaf traits | 常绿植物Evergreen species | 落叶植物Deciduous species | ||
| N再吸收效率(NRE) Nitrogen resorption efficiency | P再吸收效率(PRE) Phosphorus resorption efficiency | N再吸收效率(NRE) Nitrogen resorption efficiency | P再吸收效率(PRE) Phosphorus resorption efficiency | |
| **P<0.01,*P<0.05;表中LDMC为叶干物质含量table represents leaf dry matter content;SLA为比叶面积represents specific leaf area;LT为叶片厚度represents leaf thickness;LNC为叶氮含量represents leaf nitrogen concentration;LPC为叶磷含量represents leaf phosphorus concentration | ||||
| LDMC | -0.248 | -0.145 | -0.129 | 0.147 |
| SLA | -0.148 | -0.155 | -0.177 | -0.313 |
| LT | 0.235 | 0.234 | 0.084 | 0.003 |
| LNC | -0.019 | -0.048 | -0.016 | -0.227 |
| LPC | 0.074 | 0.237 | 0.143 | -0.078 |
| N∶P | -0.014 | -0.326 | 0.041 | 0.078 |
| 叶片性状 Leaf traits | 常绿植物Evergreen species | 落叶植物Deciduous species | ||
| N再吸收效率 Nitrogen resorption efficiency | P再吸收效率 Phosphorus resorption efficiency | N再吸收效率 Nitrogen resorption efficiency | P再吸收效率 Phosphorus resorption efficiency | |
| **P<0.01,*P<0.05 | ||||
| LDMC | -0.290 | -0.513 | -0.448 | -0.184 |
| SLA | -0.011 | 0.103 | 0.022 | -0.110 |
| LT | 0.212 | 0.386 | 0.028 | -0.052 |
| LNC | -0.796** | -0.288 | -0.719** | -0.776** |
| LPC | -0.330 | -0.786** | -0.602* | -0.828** |
| N∶P | -0.451 | 0.501 | 0.196 | 0.384 |
根据Killingbeck等人[29]在分析了89个常绿和落叶木本多年生植物老叶中的N、P含量后提出的方法,直接用衰老叶片中的养分含量来表示养分再吸收程度,叶片衰老过程中N、P含量分别降到7 mg/g和0.5 mg/g以下时,植物养分再吸收程度最大,此时认为叶片对养分是完全再吸收的;当N、P含量分别达到10 mg/g和0.8 mg/g以上时,植物具有较低的再吸收程度,衰老叶片中养分含量越低,表示植物再吸收程度越高,反之越低,常绿植物再吸收P的程度显著高于落叶植物(在老叶中P含量分别为0.45 mg/g和0.67 mg/g)。本文研究发现,常绿植物(n=15)和落叶植物(n=14)衰老叶中N和P的含量分别为11.0、0.548 mg/g和14.9、0.702 mg/g,表明对于生长在喀斯特石灰岩地区的植物N和P具有较高的再吸收程度,常绿植物再吸收N和P程度高于落叶植物,这与Killingbeck研究的结果相似。这表明常绿植物对养分元素具有较高的再吸收水平。
在大多数陆地生态系统中,N和P的有效性限制了植物的生长,可依据植物成熟叶中N、P含量及N ∶ P比作为植物营养限制的标准[7],成熟叶中N ∶ P>16,且P含量<1.0 mg/g,受P限制;N ∶ P<14,同时N含量<20.0 mg/g,受N限制;介于二者之间表示N和P共同限制。本研究中喀斯特地区常绿和落叶植物N ∶ P在成熟叶和衰老叶中均不存在显著差异,且N ∶ P均高于16,同时P含量大都低于1.0 mg/g,说明喀斯特石灰岩地区植物生长主要受到P素供应的限制。
已有的研究表明:养分再吸收效率与成熟叶片中养分含量之间没有必然的联系[30, 31] 。Aerts[2]也认为成熟叶养分含量与养分再吸收效率之间没有显著相关性,但养分再吸收效率与衰老叶中养分含量之间存在显著负相关,这与本文的研究结果相同,N、P再吸收效率与衰老叶中N、P含量呈显著负相关,也就是说,衰老叶中养分含量越低,养分再吸收效率越高,即养分再吸收程度越高,其养分再吸收效率就越高,这说明植物是通过降低衰老叶中养分含量来提高养分再吸收效率的。Kobe等人[11]的研究结果也表明,无论是种间还是种内,叶片中养分含量的升高会导致再吸收效率的下降,相反,叶片中养分含量的下降会引起再吸收效率的升高。本研究还发现,养分再吸收效率与叶片结构性状(LDMC、SLA、LT)相关性不显著(P>0.05),但目前有关这一方面的研究还比较匮乏,具体机理有待进一步研究。
喀斯特山地土层浅薄,土壤不连续,岩石裸露率高,土壤富钙而偏碱性,土壤肥沃但总量少,水分和养分易于流失。正是由于这种原因,每年都有大量的养分从生态系统中流失,在养分不足的石灰岩山地养分不断流失而又得不到补充的情况下,系统的养分供应迟早会成为限制植物生长的主要因素,生长在这种生境的植物养分再吸收效率较低,表明植物N、P养分保存能力降低,植物在对养分保存上则表现出一种衰退趋势,这就决定喀斯特生境的植物具有低生产力和低生物量的特征。同时,养分再吸收还会影响叶片枯落物中的C ∶ P,从而影响叶片枯落物的分解速率及土壤中有效N的移动,最终影响生态系统的养分循环和稳定性。
4 结论喀斯特石灰岩地区有很多的特有植物,它们对钙质土的适应机理有可能为开发喀斯特植物资源和筛选喀斯特适生植物提供理论基础,本研究对石灰岩地区常绿和落叶两种生活型植物叶片养分含量、养分再吸收状况做了对比,其中基于单位质量叶片计算的养分含量,常绿植物成熟叶中N含量和P含量高于落叶植物,衰老叶中N含量常绿植物显著低于落叶植物;基于单位面积叶片计算的养分含量则表现出相反的趋势,常绿植物成熟和衰老叶中N含量和P含量均低于落叶植物,但所有植物叶片养分含量总体上表现出较低的特征,基于不同方法计算的N ∶ P及其再吸收效率在两类植物中无显著差异,且N ∶ P均大于16,可见该地区植物生长主要受P素制约。N、P再吸收效率与叶性因子之间关系的研究结果显示,养分再吸收效率与叶干物质含量、比叶面积、叶片厚度及成熟叶片中的养分含量无显著关系,而与衰老叶中N、P含量呈显著负相关,这说明衰老叶中养分含量越低,养分再吸收效率越高,可见石灰岩地区无论是常绿植物还是落叶植物,它们是通过降低衰老叶片中的养分含量,即提高养分再吸收程度和养分再吸收效率来适应贫瘠立地的。
| [1] | Killingbeck K T. The terminological jungle revisited: making a case for use of the term resorption. Oikos, 1986, 46(2): 263-264. |
| [2] | Aerts R. Nutrient resorption from senescing leaves of perennials: are there general patterns? The Journal of Ecology, 1996, 84(4): 597-608. |
| [3] | Leonardus V, Stefano M, Amilcare P, Novais R F, Jackson R B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecological Monographs, 2012, 82(2): 205-220. |
| [4] | Chapin F S III. The mineral nutrition of wild plants. Annual Review Ecology and Systematics, 1980, 11(1): 233-260. |
| [5] | Aerts R. Nutrient use efficiency in evergreen and deciduous species from heathlands. Oecologia, 1990, 84(3): 391-397. |
| [6] | May J D, Killingbeck K T. Effects of presenting nutrient resorption on plant fitness and foliar nutrient dynamics. Ecology, 1992, 73(5): 1868-1878. |
| [7] | Santa Regina I, Leonardi S, Rapp M. Foliar nutrient dynamics and nutrient-use efficiency in Castanea sativa coppice stand of southern Europe. Forestry, 2001, 74(1): 1-10. |
| [8] | 曾德慧, 晨光生, 陈伏生, 赵琼, 冀小燕. 不同林龄樟子松叶片养分含量及其再吸收效率. 林业科学, 2005,41(5): 21-27. |
| [9] | 李荣华, 汪思龙, 王清奎. 不同林龄马尾松针叶凋落前后养分含量及回收特征. 应用生态学报, 2008, 19(7): 1443-1447. |
| [10] | 叶功富, 张立华, 林益明, 王亨, 周海超, 曾琦. 福建东山短枝木麻黄小枝氮磷含量及其再吸收率季节动态. 生态学报, 2009, 29(12): 6519-6526. |
| [11] | Kobe R K, Lepczyk C A, Meera I. Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology, 2005, 86(10): 2780-2792. |
| [12] | Aerts R, Chapin F S. The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 2000, 30: 1-67. |
| [13] | Venterink H O, Wassen M J, Verkroost A W M, De Ruiter P C. Species richness-productivity patterns differ between N-,P-,and K-limited wetlands. Ecology, 2003, 84(8): 2191-2199. |
| [14] | 苏波, 韩兴国,黄建辉, 渠春梅. 植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略. 生态学报, 2000, 20(2): 335-343. |
| [15] | 邢雪荣, 韩兴国, 陈灵芝. 植物养分利用效率研究综述. 应用生态学报, 2000, 11(5): 785-790. |
| [16] | 陈伏生, 胡小飞, 葛刚. 城市地被植物麦冬叶片氮磷化学计量比和养分再吸收效率. 草叶学报, 2007, 16(4): 47-54. |
| [17] | 阴黎明, 王力华, 刘波. 文冠果叶片养分含量元素的动态变化及再吸收特征. 植物研究, 2009, 29(6): 685-691. |
| [18] | 赵琼, 刘兴宇, 胡亚林, 曾德慧. 氮添加对兴安落叶松养分分配和再吸收效率的影响. 林业科学, 2010, 46(5): 14-19. |
| [19] | Liu C Q. Biogeochemical Processes and Cycling of Nutrients in the Earth's Surface: Cycling of Nutrients in Soil-plant Systems of Karstic Environments, Southwest China. Beijing: Science Press, 2009. |
| [20] | Chen J W, Zhang Q, Cao K F. Inter-species variation of photosynthetic and xylem hydraulic traits in the deciduous and evergreen Euphorbiaceae tree species from a seasonally tropical forest in southwestern China. Ecological Research, 2009, 24(1): 65-73. |
| [21] | Fan D Y, Jie SL, Liu C C, Zhang X Y, Xu X W, Zhang S R, Xie Z Q. The trade-off between safety and efficiency in hydraulic architecture in 31 woody species in a karst area. Tree Physiology, 2011, 31(8): 865-877. |
| [22] | 郭柯, 刘长成, 董鸣. 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报, 2011, 35(10): 991-999. |
| [23] | 朱守谦. 喀斯特森林生态研究II. 贵阳: 贵州科技出版社, 1997. |
| [24] | 朱守谦. 喀斯特森林生态研究III. 贵阳: 贵州科技出版社, 2003. |
| [25] | 范文武, 陈晓德, 李加海, 张淑琴, 李艳霞. 重庆中梁山海石公园石灰岩山地植物多样性研究. 西南大学学报: 自然科学版, 2009, 31(5): 106-110. |
| [26] | Cornelissen J H C, Lavorel S, Gamier E, Díaz S, Buchmann N, Gurch D E, Reich P B, ter Steege H, Morgan H D, van der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for Standardized and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335-380. |
| [27] | 宝乐, 刘艳红. 东灵山地区不同森林群落叶功能性状比较. 生态学报, 2009, 29(7): 3692-3703. |
| [28] | 方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [29] | Killingbeck K T. Nutrients in senesced leaves:keys to the search for potential resorption and resorption proficiency. Ecology, 1996, 77(6): 1716-1727. |
| [30] | Del Arco J M, Escudero A, Garrido V M. Effects of site characteristics on nitrogen retranslocation from senescing leaves. Ecology, 1991, 72(2): 701-708. |
| [31] | Wright I J, Westoby M. Nutrient concentration, resorption and life span: leaf traits of Austrlian sclerophyll species. Functional Ecology, 2003, 17(1): 10-19. |