 2015, Vol. 35
2015, Vol. 35文章信息
- 谈冰畅, 朱小龙, 宁晓雨, 谷娇, 安苗, 李宽意
- TAN Bingchang, ZHU Xiaolong, NING Xiaoyu, GU Jiao, AN Miao, LI Kuanyi
- 纹沼螺对苦草生长的影响
- Effects of Parafossarulus striatulus on the Growth of Vallisneria spiralis
- 生态学报, 2015, 35(10): 3199-3203
- Acta Ecologica Sinica, 2015, 35(10): 3199-3203
- http://dx.doi.org/10.5846/stxb201405100941
-
文章历史
- 收稿日期:2014-05-10
- 网络出版日期:2014-08-13






2. 中国科学院南京地理与湖泊研究所, 南京 210008;
3. 江南大学, 无锡 214122
2. Nanjing Institute of Geography & Limnology, Chinese Academy of Sciences, Nanjing 210008, China;
3. Jiangnan University, Wuxi 214122, China
在水生态系统中,沉水植物对于维持水体清水态起着极其重要的作用[1],随着水体富营养化的加剧,许多湖泊中的沉水植物均不同程度地减少甚至消亡。沉水植被消亡的主要机理之一是由于植物表面附着藻类过度繁殖生长[2],形成了与沉水植物在光照、二氧化碳等资源方面的竞争[3]。螺类是水生态系统中常见的大型软体动物,主要以附着生物、植物碎屑等为食,通过刮食去除植物表面的附着生物,减弱后者对沉水植物的抑制作用,从而有利于沉水植物的生长[4]。
关于淡水螺类与沉水植物之间的生态关系,国内外研究较多的螺类主要有铜锈环棱螺(Bellamya aeruginosa)[5]、椭圆萝卜螺(Radix swinhoei)[6, 7, 8, 9]和静水椎实螺(Lymnaea stagnalis)[10, 11]。已有研究表明,不同密度的铜锈环棱螺均明显促进了受附着生物胁迫的苦草的生长[5]。野外调查发现,与环棱螺相比,纹沼螺(Parafossarulus striatulus)个小、体轻,更易于附着于水草上,初步观察可知纹沼螺与水草之间关系更密切,两者是否也存在“螺-草”互利现象?针对此问题,国内外尚未见报道。
纹沼螺通常是浅水草型湖泊中的优势种类[12],属前腮亚纲(Prosobranchia),中腹足目(Mesogastropoda),豆螺科(Bithyniidae),沼螺属(Parafossarulus)。本文选用纹沼螺为目标螺,设置不同螺密度,研究它们对太湖常见沉水植物——苦草(Vallisneria spiralis)生长的影响,进而探讨螺类与沉水植物之间的生态关系。
1 材料与方法室外受控实验于2013年9月15日至10月15日在苏州太湖基地进行。实验设一组无螺对照及两组有螺处理(低密度为150 个/m2;高密度为450 个/m2),每个处理组设4个重复。实验所用容器为12个高强度的聚乙烯塑料桶(桶高70 cm,上、下底直径分别为48 cm和40 cm)。为了控制水温,实验桶放置在水深50 cm的室外水泥池中。实验所用的苦草与纹沼螺从太湖采集。实验所用沉积物来自东太湖,经60目过筛混匀后使用,沉积物中总氮(TN)、总磷(TP)和有机质(OM)含量分别为(3.24±0.07)mg/g、(0.57±0.20)mg/g和(76.20±1.49)mg/g。实验用水为500目过滤的东太湖湖水,其初始水质指标TN、TP和叶绿素a(Chl a)浓度分别为(0.74±0.08)mg/L、(32.40±7.43)μg/L和(15.42±1.09)μg/L。
实验设置沉积物厚度为10 cm,水深40 cm。等水体变清后选取8株苦草移栽至桶中,各桶苦草初始总湿重基本一致(表 1),苦草株高约15—17 cm,叶片5—7片,颜色亮绿。然后根据实验设计在桶中放入规格相近、不同密度的纹沼螺(表 1)。为了采集附着生物,在各桶内放置4块硬塑料板(8 cm×10 cm),塑料板与实验桶材质一致,塑料板下方用棉绳系一螺帽,利用水的浮力使塑料板保持垂直状态,与苦草叶片方向一致。实验期间,每天观察桶内纹沼螺与苦草的生长情况,为保证螺密度的稳定,将新生螺移出水体,实验中随时补充水以保证水量不变。实验期间桶内水温变化范围为29.5—36℃。
| 螺密度 Snail density/(个/m2) | 螺总湿重 Snail total wet weight/g | 苦草总湿重 Plant total wet weight/g |
| 0 | 0 | 19.35±0.90 |
| 150 | 3.31±0.23 | 19.23±0.81 |
| 450 | 12.06±0.47 | 19.60±0.58 |
每隔7d采用YSI水质仪测定水温、pH等指标,采集水样与附着生物样,分析水体叶绿素a、总氮和总磷含量,测定单位面积附着生物干重,其方法依据《湖泊生态调查观测与分析》[13]。整个实验期间共采集水样与附着生物样品4次。实验结束后,测定苦草湿重、叶片数和株高等生长指标,计算苦草的相对生长率,其计算公式如下:
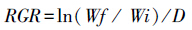
式中,Wi和Wf分别为植物实验前、后的湿重,D为天数。
应用办公软件Microsoft Excel 2010对数据进行计算处理,统计软件SPSS 21.0中One-Way ANOVA方法对数据差异进行分析,作图软件GraphPad Prism 5对数据绘制柱状图与线图。
2 结果与分析 2.1 苦草生长指标纹沼螺对苦草生长有明显促进作用(P<0.05),随着螺密度的升高,苦草的相对生长率、叶片数、株高均逐步提高(图 1)。具体而言,不同密度螺处理下的苦草相对生长率差异显著(P=0.019),低密度组和高密度组的苦草相对生长率分别为(48.35±1.89)mg g-1 d-1和(51.67±0.97)mg g-1 d-1,均高于无螺组((40.29±2.46)mg g-1 d-1),升幅分别为20%(P=0.023)和28%(P=0.009);苦草叶片数同样有显著差异(P=0.038),与无螺组的苦草叶片数(101±3)相比,高密度组(116±3)增加了15%(P=0.019)。苦草株高的差异也极其显著(P=0.001),低密度组和高密度组的苦草株高,分别为(25.0±1.4)cm和(26.3±1.5)cm,均显著高于无螺组((20.8±1.5)cm),升幅分别为20%(P=0.003)和27%(P=0.001)。由此可见,纹沼螺的存在促进了苦草生长。

|
| 图1 苦草的相对生长率、叶片数和株高 (平均值±标准差) Fig.1 The relative growth rate,number of leaves and plant height of V. spiralis (Mean ± SD) |
纹沼螺明显降低了单位面积附着生物的干重(图 2)。无螺组的附着生物干重远高于有螺处理组(P<0.01)。随着纹沼螺密度的升高,附着生物干重显著降低,高密度组的附着生物干重最低(P<0.01)。

|
| 图2 附着生物干重的变化 (平均值±标准差) Fig.2 Changes of periphyton dry mass (Mean ± SD) |
不同纹沼螺密度处理的水体营养盐浓度和叶绿素a含量差异显著(图 3)。低密度组的总氮、总磷和叶绿素浓度均高于无螺组和高密度组(P<0.05),其变化范围分别为0.83—1.12 mg/L、33.69—70.45 μg/L和6.31—29.50 μg/L。高密度组的总氮、总磷和叶绿素浓度最低(P<0.01),变化范围分别为0.71—0.87 mg/L、22.73—31.11 μg/L和3.13—7.63 μg/L。

|
| 图3 水体总氮、总磷与叶绿素a变化 (平均值±标准差) Fig.3 Changes of the concentration of total nitrogen,total phosphorus and Chl a in water (Mean ± SD) |
沉水植物的生长受到光照、营养盐和水生动物牧食等诸多因素的综合作用。Qin等认为浮游植物的大量增加会导致水体的透明度下降,从而使沉水植物的光合作用受到抑制,这可能是沉水植物消失的原因之一[14]。就本实验而言,虽然不同处理之间浮游植物生物量(叶绿素含量)差别显著,但就绝对值而言较低,实验期间确实也观察到水体清澈见底,因此浮游植物的遮光作用应该不是限制苦草生长的主要因素。沉积物与水体营养盐的含量也决定沉水植物生长的优劣,Xie等发现沉积物是有根水生植物获取营养的主要来源[15],如根系发达的苦草其生长主要由沉积物营养水平决定,叶片获取水体中营养的作用较弱[16]。本实验使用的是同一种沉积物,虽然各处理组水体氮磷浓度有显著差别,但绝对值较低,因此水体营养盐水平应该也不是影响苦草生长的主要因素。Liboriussen等研究表明沉水植物表面的附着藻类与植物存在光照、营养盐等竞争,水体中的营养盐优先被附着藻类吸收,从而使附着藻类大量增加,附着生物的过度覆盖会对沉水植物产生不利影响[17]。本实验结果显示随着纹沼螺密度升高,对附着生物的牧食作用增强,有效地控制了附着生物量,实验中附着生物的干重显著降低,而苦草的叶片数和株高明显提高、生长加快。这与相关研究结果一致,即附着藻类对沉水植物的光竞争是沉水植被消亡的关键因子[18],螺类通过牧食附着藻类促进了沉水植物生长[2]。
在水生态系统中,底栖动物通过新陈代谢活动及生物扰动影响水体营养盐浓度[19]。Underwood发现螺类通过新陈代谢释放营养物质[20],促进局部水体溶解态氮磷含量增加[21]。同时,螺类的牧食活动也引起了少量沉积物的再悬浮[22],这也可能增加了水体营养盐含量。本实验中低密度螺类处理组水体营养盐含量高于对照组,可能与这两个因素有关。但奇怪的是,高密度组水体TN和TP均低于其余两组。这可能有两方面的原因,一是随着螺密度的增加,对附着生物的牧食压力加大,附着生物层变薄,有利于这部分附着藻类吸收营养盐,使实验水体中营养盐浓度降低;二是附着生物大量减少,与植物叶片争夺营养盐的竞争减弱,有利于植物叶片吸收营养,虽然苦草主要从沉积物中吸收营养,但叶片也有吸收营养盐的辅助作用[16]。因此,高密度组水体TN和TP均低于其余两组,可能是附着藻类与苦草叶片的营养盐吸收超过螺生物扰动与新陈代谢营养盐释放的结果。当然这有待进一步实验验证。
环棱螺与纹沼螺均为长江中下游水体常见螺类,环棱螺往往是很多湖泊中的优势种,而纹沼螺通常是草型湖泊的优势种。蔡永久等发现环棱螺在太湖全湖均有分布,在草型湖湾东太湖的密度则高达299 个/m2[23]。龚志军等对武汉东湖的调查表明环棱螺密度最高可达157 个/m2[24],而陈其羽对东湖早期的调查发现密度可高达500 个/m2以上[25]。有研究表明在环棱螺密度为160、320和640 个/m2的受控实验中,螺类牧食活动均极显著地促进了苦草生长,证实二者之间存在螺-草互利关系[5],由于其设置的实验螺密度在自然水体中广泛存在,因而环棱螺减轻了湖泊中附着藻类对沉水植物生长的抑制作用,促进了沉水植物的生长,从而保护了沉水植物,进而维持了湖泊的清水态。与环棱螺不同,纹沼螺在自然水体中的密度较低。蔡永久等实地调查洪湖、赤湖和大冶湖的纹沼螺密度分别为64、96和69 个/m2[26]。闫云君等发现湖北扁担塘的纹沼螺种群密度为48 个/m2左右,于5月下旬出现峰值(158 个/m2),但此时大多为刚孵化出的幼体,成螺密度较低[27]。本实验中纹沼螺密度为150 与450 个/m2,在该密度时纹沼螺均显著促进了苦草生长,因而在理论上这两者之间也存在螺-草互利关系。由于本实验设计的纹沼螺密度要高于自然水体,在湖泊中是否存在螺-草互利关系尚需进一步实验验证。但就统计结果的显著程度而言,Li等结果表明低密度环棱螺(160 个/m2)极显著地促进了苦草生长(P<0.001)[5],而本实验中低密度纹沼螺(150 个/m2)对苦草生长的促进作用仅为显著(P<0.05),因而推断自然水体中纹沼螺的牧食活动促进沉水植物生长的作用可能远不如铜锈环棱螺。另外,调查表明环棱螺在湖泊中的分布不受沉水植物制约,但其分布密度往往与沉水植物的分布状况有关,而纹沼螺的分布密度与沉水植物的丰度正相关,完全受沉水植物分布的制约[28]。因此,就螺-草关系而言,在自然生态系统中纹沼螺与植物的关系更为密切,对沉水植物的依赖程度相对环棱螺要强些。
| [1] | Hobbs W O, Hobbs J M R, Lafrancois T, Zimmer K D, Theissen K M, Edlund M B, Michelutti N, Butler M G, Hanson M A, Carlson T J. A 200-year perspective on alternative stable state theory and lake management from a biomanipulated shallow lake. Ecological Applications, 2012, 22(5): 1483-1496. |
| [2] | Li K Y, Liu Z W, Gu B H. Persistence of clear water in a nutrient-impacted region of Lake Taihu: The role of periphyton grazing by snails. Fundamental and Applied Limnology/Archiv für Hydrobiologie, 2008, 173(1): 15-20. |
| [3] | Jones J I, Young J O, Eaton J W, Moss B. The influence of nutrient loading, dissolved inorganic carbon and higher trophic levels on the interaction between submerged plants and periphyton. Journal of Ecology, 2002, 90(1): 12-24. |
| [4] | Jones J I, Sayer C D. Does the fish-invertebrate-periphyton cascade precipitate plant loss in shallow lakes? Ecology, 2003, 84(8): 2155-2167. |
| [5] | 李宽意, 文明章, 杨宏伟, 李艳敏, 刘正文. "螺-草"的互利关系. 生态学报, 2007, 27(12): 5427-5432. |
| [6] | 李宽意, 刘正文, 李传红, 李艳敏, 文明章. 太湖椭圆萝卜螺的食物来源分析. 湖泊科学, 2008, 20(3): 339-343. |
| [7] | 李宽意, 刘正文, 杨宏伟, 宋晓兰, 文明章. 螺类牧食与沉积物类型对苦草生长的影响. 生态学报, 2007, 27(11): 4907-4912. |
| [8] | Li K Y, Liu Z W, Hu Y H, Yang H W. Snail herbivory on submerged macrophytes and nutrient release: implications for macrophyte management. Ecological Engineering, 2009, 35(11): 1664-1667. |
| [9] | 李宽意,刘正文,胡耀辉,王传海.椭圆萝卜螺 Radix swinhoei(H.Adams)对三种沉水植物的牧食选择.生态学报, 2006, 26(10): 3221-3224. |
| [10] | Pinowska A. Effects of snail grazing and nutrient release on growth of the macrophytes Ceratophyllum demersum and Elodea canadensis and the filamentous green alga Cladophora sp.. Hydrobiologia, 2002, 479(1/3): 83-94. |
| [11] | Elger A, Lemoine D. Determinants of macrophyte palatability to the pond snail Lymnaea stagnalis. Freshwater Biology, 2005, 50(1): 86-95. |
| [12] | 梁彦龄, 刘伙泉. 草型湖泊资源、环境与渔业生态学管理(一). 北京: 科学出版社, 1995. |
| [13] | 黄祥飞. 湖泊生态调查观测与分析. 北京: 中国标准出版社, 1999: 72-79. |
| [14] | Qin B Q, Gao G, Zhu G W, Zhang Y L, Song Y Z, Tang X M, Xu H, Deng J M. Lake eutrophication and its ecosystem response. Chinese Science Bulletin, 2013, 58(9): 961-970. |
| [15] | Xie D, Yu D, You W H, Wang L G. Morphological and physiological responses to sediment nutrients in the submerged macrophyte Myriophyllum spicatum. Wetlands, 2013, 33(6): 1095-1102. |
| [16] | Carignan R. An empirical model to estimate the relative importance of roots in phosphorus uptake by aquatic macrophytes. Canadian Journal of Fisheries and Aquatic Sciences, 1982, 39(2): 243-247. |
| [17] | Liboriussen L, Jeppesen E. Temporal dynamics in epipelic, pelagic and epiphytic algal production in a clear and a turbid shallow lake. Freshwater Biology, 2003, 48(3): 418-431. |
| [18] | Phillips G L, Eminson D, Moss B. A mechanism to account for macrophyte decline in progressively eutrophicated freshwaters. Aquatic Botany, 1978, 4: 103-126. |
| [19] | Karsten R. Tidal Flat Ecology: An Experiment Approach to Species Interaction. New York: Springer-Verlag, 1985. |
| [20] | Underwood G J C. Growth enhancement of the macrophyte Ceratophyllum demersum in the presence of the snail Planorbis planorbis: the effect of grazing and chemical conditioning. Freshwater Biology, 1991, 26(2): 325-334. |
| [21] | Steinman A D. Effects of grazes on freshwater benthic algae//Stevenson R J, Bothwell M L, Lowe R L. Algal Ecology. San Diego: Academic Press, 1996: 341-373. |
| [22] | 魏阳春, 濮培民. 太湖铜锈环棱螺对氮磷的降解作用. 长江流域资源与环境, 1999, 8(1): 89-93. |
| [23] | 蔡永久, 龚志军, 秦伯强. 太湖软体动物现存量及空间分布格局(2006-2007年). 湖泊科学, 2009, 21(5): 713-719. |
| [24] | 龚志军, 李艳玲, 谢平. 武汉东湖铜锈环棱螺的种群动态及次级生产力. 湖泊科学, 2009, 21(3): 401-407. |
| [25] | 陈其羽. 武汉东湖铜锈环棱螺种群变动和生产量的初步观察. 水生生物学报, 1987, 11(2): 117-130. |
| [26] | 蔡永久, 姜加虎, 张路, 陈宇炜, 龚志军. 长江中下游湖泊大型底栖动物群落结构及多样性. 湖泊科学, 2010, 22(6): 811-819. |
| [27] | 阎云君, 梁彦龄, 王洪铸. 扁担塘螺类生产力的研究Ⅱ. 纹沼螺的周年生产量. 水生生物学报, 2001, 25(1): 36-41. |
| [28] | 陈其羽. 湖北省花马湖软体动物的调查报告. 海洋与湖沼, 1979, 10(1): 46-62. |