 2015, Vol. 35
2015, Vol. 35文章信息
- 刘勇, 程家骅
- LIU Yong, CHENG Jiahua
- 东海区底拖网作业三类主要渔获物的时空关系
- Spato-temporal relationships among three groups of fishes caught in bottom trawl survey in the East China Sea
- 生态学报, 2015, 35(10): 3403-3410
- Acta Ecologica Sinica, 2015, 35(10): 3403-3410
- http://dx.doi.org/10.5846/stxb201306201745
-
文章历史
- 收稿日期:2013-06-20
- 网络出版日期:2014-05-30






海洋是地球生命的起源,其中的生物品种极其丰富,至今还有人类未发现、待探索的未知生物。渔船是目前人类开发和利用海洋生物资源的主要方式,其捕捞对象主要是有经济价值的品种,这些品种可分为三大类:鱼类、甲壳类和头足类。这三类生物隶属于动物界的不同分类,鱼类属于有脊索动物,而甲壳类和头足类均属于无脊索动物。三类生物生存空间存在差异、但不完全隔绝,不管从水平方向、还是从垂直方向,彼此都会存在不同程度的重叠,这种空间关系还会随时间变化而变化[1, 2];三类生物存在诸多联系,如同处环境稳定、条件适宜的产卵场和育卵场,或在饵料丰富的索饵场共同进食,或存在捕食与被捕食[3, 4, 5]等联系,还有更多的关系有待进一步研究和发现。
目前国外许多专家提倡对海洋生物资源进行基于生态系统水平的管理[6, 7, 8],国内越来越多的学者也认识到研究海洋生态系统的重要性[9, 10, 11],海洋生态系统研究正在成为国际发展的趋势和国内发展的需求。基于生态系统的管理与研究要求把一定时空内各种生物之间以及生物群落与无机环境之间的相互作用看作一个统一的整体来进行研究,而国内关于海洋资源方面的研究目前还多数是关于单类别群落的研究,如研究得较多的是鱼类群落[12, 13, 14, 15],甲壳类和头足类的研究相对较少[16, 17, 18],有关三类海洋生物联系的综合研究还尚未见报告。本文拟据一年四季度东海区大面积定点调查资料,分析三类海洋生物空间分布关系,以期了解东海区生态系统主要生物之间的时空关系,为以后生态系统进一步研究提供基础资料。
1 材料和方法 1.1 材料来源数据来自2000年东海区渔业资源春(4月)、夏(6月)、秋(9月)、冬(12月)四次大面积定点调查。调查船为双拖渔轮,功率为183.25 kW / 艘,网具为100 目 × 4 m,网囊网目为2.5 cm,平均拖速为2海里/h。调查范围为27°00′—34°00′N,122°00′—127°00′E,经、纬度每隔30′设1个站位,格状均匀设置站位。
1.2 分析方法 1.2.1 3种渔业生物分类方法本研究采用的渔获数据共包含340种物种,其中鱼类235种,甲壳类73种,头足类32种[19]。表 1列出的是四季中组成占生物量80%之上的主要物种名称及拉丁文名称。
| 分类Type | 种名Specific name |
| 鱼类 Fish |
刺鲳Psenopsis anomala (Temminck et Schlegel,1844) |
| 带鱼Trichiurus japonicus (Temminck et Schlegel,1844) | |
| 刀鲚Coilia ectenes (Jordan et Seale 1905) | |
| 多刺腔吻鳕Coelorhynchus multispinulosus (Katayama,1942) | |
| 鳄齿鱼Champsodon capensis (Regan,1908) | |
| 发光鲷Acropoma japonicum (Güther,1859) | |
| 黑鮟鱇Lophiomus setigerus (Vahl. 1797) | |
| 黑鳃梅童鱼Collichthys niveatus (Jordan et Starks,1906) | |
| 黄鮟鱇Lophius litulon (Jordan,1912) | |
| 黄鲫Setipinna taty (Valenciennes,1848) | |
| 棘头梅童鱼Collichthys lucidus (Richardson,1844) | |
| 龙头鱼Harpadon nehereus (Hamilton,1822) | |
| 日本鲭Scomber japonicus (Houttuyn,1782) | |
| 鳀Engraulis japonicus (Temminck et Schlegel,1846) | |
| 细条天竺鲷Apogon lineatus (Temmick et Schleger,1842) | |
| 细纹狮子鱼Liparis tanakae (Gilbert et Burke,1912) | |
| 小黄鱼Pseudosciaena polyactis (Bleeker,1877) | |
| 银鲳Pampus argenteus (Euphrasen,1788) | |
| 竹荚鱼Trachurus japonicus (Temminck et schlegel,1842) | |
| 甲壳类 Crustacean |
细点圆趾蟹Ovalipes punctatus (de Haan,1833) |
| 圆板赤虾Metapenaeopis lata (Kubo,1949) | |
| 头足类 Cephalopod |
剑尖枪乌贼Loligo edulis (Hoyle,1885) |
| 神户枪乌贼Loligo kobiensis (Hoyle,1885) | |
| 太平洋褶柔鱼Todarodes pacificus (Steenstrup,1880) |
本研究是基于大面积定点调查数据,其调查站点覆盖面广且分布均匀,资源的聚集、分散等分布特征均能通过该数据有所体现。基于上述内容,进而可以根据不同站点的资源密度指数,寻找资源的主要分布区域。本文定义“资源主要分布区域”为资源重量累积百分比达到指定比例的分布水域,该水域边缘对应的资源密度指数,即该水域密度指数最低的下限,该下限密度指数通过以下公式计算:
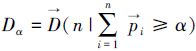
式中,α为期望资源累积百分比,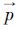 为降序排列的站点密度指数占总量的百分比序列,i是序列中第i个百分比值,n是序列前n个百分比值累加达到α时的序号,
为降序排列的站点密度指数占总量的百分比序列,i是序列中第i个百分比值,n是序列前n个百分比值累加达到α时的序号,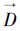 为降序排列的站点密度指数序列,其第n个密度指数即为资源累积百分比达到期望值的下限密度指数(Dα)。
为降序排列的站点密度指数序列,其第n个密度指数即为资源累积百分比达到期望值的下限密度指数(Dα)。
物种之间距离是采用欧氏距离[20],计算公式如下:
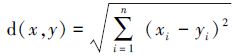
式中,x为第1类物种在各站点上调查获得的资源密度序列,y为第2类物种在各站点上调查获得的资源密度序列,i是对应各个站点的编号。
1.2.4 等值面的插值方法本文采用Akima插值方法[21],该方法是一种二项式插值方法,多数情况下可以达到三次多项式的精确度,是一种局部的、基于三角形算法的插值方法,可以对不规则分布数据进行平面平滑适应插值[21, 22]。Akima方法是一种容易实现、适应性强、计算迅速的插值方法,但估算边缘值相对偏弱[23]。本研究目的是找出生物主要分布区域,边缘数据影响不大,考虑其方便快捷,从而采用了该插值方法。Akima插值方法是利用R统计软件[24]akima工具包来实现[21]。
1.2.5 聚类分析方法采用的是系统聚类法,选择的聚类统计量是欧氏距离,聚类方法采用最长距离法[25],具体计算过程是通过R统计软件[24]amap工具包[26]中的hcluster函数实现。
2 结果 2.1 三类生物四季空间分布三类生物春夏秋冬四个季节的分布情况如图 1所示。观察可以看到,鱼类的生物量明显高于其它两类生物。分布空间上来看,春季鱼类主要集中在近海水域,头足类主要分布在东海中部的外海水域,而甲壳类只有零星水域分布;夏季鱼类的分布水域有所扩大,在近海和外海均出现相对集中分布水域,甲壳类分布相对春季有所扩大,在东海中部外海有一个相对集中水域,头足类分布范围相比春季也有所扩大,在偏南水域出现一个相对集中水域;秋季鱼类分布范围扩大,主要分布在东海北部水域,东海中部偏南水域也有一个相对集中水域,甲壳类分布范围扩大,也主要集中在东海北部水域,头足类分布范围相比夏季有所扩大,但分布范围仍较小,主要集中在东海中部偏南水域;冬季鱼类的分布范围缩小,但主要还是分布在黄海南部,甲壳类分布范围相比秋季缩小,东海中部和黄海南部都有相对集中水域,头足类分布范围缩小,主要集中在东海中部偏南水域。

|
| 图 1 三类生物四个季节的分布情况(此文所有地图数据均由国家测绘局提供) Fig.1 The distribution of 3 types of organisms in 4 seasons |
在前面分析的基础上,本研究进一步分析三类生物主要分布范围之间的关系,即把资源相对集中的范围区分出来,从而能更清晰的分析三类生物空间上的相互联系。分别把资源量累积百分比达到90%、80%和60%的主要分布水域描绘出来后,发现三类生物的主要分布水域有明显分离的现象,并且随着累积百分比的降低,这种现象越为明显。图 2是资源量累积百分比分别达到90%、80%和60%的情况。

|
| 图 2 三类生物累积生物量达到90%、80%和60%的集中分布水域四季分布情况 Fig.2 Concentrated areas of 3 types of organisms in 4 seasons at 90%,80% and 60% biomass accumulations |
3类生物空间分布会随着季节变化而变化,从而导致3类生物之间的距离发生改变,表 2列出了3类生物在不同季节之间的欧氏距离。可以看到春季和冬季,分别根据重量和尾数计算的距离表现一致的趋势,即在春季甲壳类与头足类之间的距离最远,而头足类与鱼类的最近;在冬季鱼类与甲壳类之间的距离最远,而头足类与鱼类的最近。夏季和秋季根据重量和尾数计算的距离趋势不一致;在夏季,根据重量计算的最远距离出现在甲壳类与头足类之间,而根据尾数计算最远距离出现在鱼类与甲壳类之间,而最近距离均出现在头足类与鱼类之间;在秋季,根据重量计算的最远和最近距离分别出现在甲壳类与头足类之间和鱼类与甲壳类之间,根据尾数计算的最远和最近距离分别出现在鱼类与甲壳类之间和甲壳类与头足类之间。以表 2中的数据分别对4个季节和3类生物之间关系做了聚类分析,分析结果见图 3。根据重量和尾数数据分析的结果类似,结果均显示秋季和冬季相似,春季和夏季相似,鱼类与甲壳类关系和甲壳类与头足类关系相比头足类与鱼类关系更相似。
| 采样季节 Season |
数据类型 Item |
物种分布距离Euclidean distances | ||
| 鱼类-甲壳类Fish-Crus. | 甲壳类-头足类Crus.-Ceph. | 头足类-鱼类Ceph.-Fish | ||
| Crus.:甲壳类Crustacea;Ceph.:头足类Cephalopoda | ||||
| 春季Spring | 重量 | 0.3449 | 0.3623 | 0.1564 |
| 尾数 | 0.5898 | 0.6274 | 0.2097 | |
| 夏季Summer | 重量 | 0.3140 | 0.3298 | 0.2870 |
| 尾数 | 0.4507 | 0.4053 | 0.3022 | |
| 秋季Autumn | 重量 | 0.1404 | 0.2168 | 0.1877 |
| 尾数 | 0.2140 | 0.1836 | 0.2011 | |
| 冬季Winter | 重量 | 0.2160 | 0.2017 | 0.1590 |
| 尾数 | 0.2971 | 0.2285 | 0.2151 | |

|
| 图 3 根据重量计算距离聚类分析结果 Fig.3 Clustering results by weight data |
由图 2可以看到,四个季节三类生物集中分布区域在空间上存在一定程度的分离,三类生物越集中的范围相对分离现象越为显著。本研究调查采用双拖渔轮,采集渔获物处于海洋底层。虽然从垂直方向来看,三类生物同处于海洋底层同一水层,但从水平分布来看,三类生物却能自然分离,均能找到各自适合的空间。这可能是三类生物相互竞争、相互适应、长期演化的结果,在同一水层找到各自适合的生存空间。
生态位是生态学中的一个重要概念,是指物种在生物群落或生态系统中的地位和角色[27]。生态位的概念抽象、含义广泛,可以粗略分为空间或栖息地生态位、营养生态位和多维或超体积生态位[28, 29]。本研究结果发现的三类生物水平空间上的相对分离现象,可以归属于空间或栖息地生态位分化的一种现象。生态学中的生态位一般描述的对象是种群之间的关系,而本研究的对象是生物类群之间的关系,可见不同生物类群之间也存在着一定程度的生态位分化现象。对生物生态位的研究,了解栖息地仅仅是开始,对生物的营养、能源、资源分配,以及与其它生物联系的研究,是对生物生态位的深入研究[29]。本研究的结果仅对大类生物存在空间生态位的分化提供了初步证据,大类生物更深层次的生态位分化的认识和分析还有待进一步研究。
3.2 三类生物空间距离四季变化特征春季和夏季期间三类生物之间的距离相对远,而在秋季和冬季三类生物之间的距离相对近。春季和夏季一般是鱼类的产卵季节,如东海小黄鱼的产卵季节主要集中在4—5月份[30],带鱼在春夏季有相对集中出生的早生群[31, 32];产卵群体在产卵前一般都要经历一段洄游,从而抵达一个环境相对适宜的水域进行集中产卵,以便提高产卵后的成活率[1]。鱼类在这个季节的分布特征一般是相对集中,而对于虾蟹类和头足类该季节的相关研究知之甚少;但从本文研究的结果来看,有两种可能,一种情况是这两类生物的分布并没有发生变化,只是因为鱼类生物分布发生了相对集中的变化,从而导致了生物之间的距离增加;另一种情况,这两类生物也可能因为产卵发生空间聚集的变化,但聚集的水域不一致,从而也会导致生物之间距离增大的结果。秋季和冬季是许多鱼类索饵洄游和越冬洄游期,其空间分布特点是向饵料聚集、水温相对温暖的水域,分布不如产卵期那么集中[31],从而导致三类生物之间距离相对更近。
头足类与鱼类之间的距离相对近,而鱼类与甲壳类、甲壳类与头足类之间的距离相对要远。从三类生物的活动方式来看,成体或大个体的甲壳类(本研究拖网船能够捕获的甲壳类)在水体的活动能力相对差,它们多数时间均处于底层,仅在特殊时间内,如受到攻击,会在垂直空间上有短暂的移动,其它时间一般均在海底作水平移动。而鱼类和头足类的活动范围则明显广阔得多,它们不仅能自由快速的在水平上移动,而且可以自由的在不同水层之间变动;两类生物之间相比,鱼类活动能力又明显强于头足类,鱼类可以维持较长时间的持续运动,而头足类仅能进行间隙性的短程运动。由于三类生物活动能力的不同,导致了三类生物在活动水层上或空间上的不同,从而导致了三类生物在生存空间上相互关系的不同。
3.3 三类生物分布特征在渔业上的管理意义本研究结果显示,三类渔业生物在水平空间分布上有着明显的分离特征;换句话说,三类生物在水平分布上处于不同水域,那么以不同种类生物为目标的捕捞渔船将会奔赴不同水域进行生产,其结果是各作业渔船分布在不同水域有条不紊的进行捕捞。但现实情况却大相径庭,渔船之间为抢占渔场发生打架斗殴事情时有发生。目前因渔船数量较多,而海洋资源并没有增加、却因过度捕捞而有所下降,因而频繁出现渔民因抢夺资源而发生争斗[33, 34]。若在知道渔业资源不同分布区域前提下,管理部门有意识引导不同作业类型渔船到不同水域进行生产,那样不仅能提高渔民寻找渔场效率,同时也会有效减少渔民之间矛盾的发生。如双拖作业渔船主要捕捞对象是经济鱼类,而横桁拖虾和笼壶作业渔船主要捕捞对象是虾蟹类,这几种渔船在同一海区生产时,就可以给予适当引导,找到各自的渔场、减少和避免渔船相互摩擦。
4 研究不足及展望本研究的结果是根据双拖作业获得的结果,主要反应的是底层游泳生物的一个基本情况。鱼类相关情况,主要反映的是底层鱼类,而不能全面反应中、上层鱼类情况,中上层鱼类的研究还有待进一步调查,可采用其它专用网具,如单拖(快速浮拖)、围网等作业方式,对中、上层鱼类的分布特点进行调查和分析。甲壳类一般处于海水最底层,对于双拖作业来说仅是附属渔获物,虽不能全面反映甲壳类情况,但基本可以反映一个大概情况;若要全面了解甲壳类的分布情况,还需借助专门捕捞虾、蟹类的作业,如桁杆拖虾、笼壶等作业方式。头足类有趋光和垂直移动习性,大部分产量由光诱作业和鱿钓作业捕捞[17],其全面分布特征也需进一步调查和研究。
致谢: 感谢参与海上调查、样品测定、数据收录等工作的同志。
| [1] | 殷名称. 鱼类生态学. 北京: 中国农业出版社, 1995: 171-187. |
| [2] | 陈大刚. 渔业资源生物学. 北京: 中国农业出版社, 1997: 101-138. |
| [3] | 韦晟. 黄海带鱼 (Trichiurus haumela Forsk I) 的摄食习性. 海洋水产研究, 1980, (1): 49-57. |
| [4] | 邓景耀, 孟田湘, 任胜民. 渤海鱼类食物关系的初步研究. 生态学报, 1986, 6(4): 356-363. |
| [5] | 韦晟, 姜卫民. 黄海鱼类食物网的研究. 海洋与湖沼, 1992, 23(2): 182-192. |
| [6] | Pikitch E K, Santora E A, Babcock A, Bakun A, Bonfil R, Conover D O, Dayton P, Doukakis P, Fluharty D, Heheman B, Houde E D, Link J, Livingston P A, Mangel M, Mcallister M K, Pope J, Sainsbury K J. Ecosystem-based fishery management. Science, 2004, 305(5682): 346-347. |
| [7] | Arkema K K, Abramson S C, Dewsbury B M. Marine ecosystem-based management: from characterization to implementation. Frontiers in Ecology and the Environment, 2006, 4(10): 525-532. |
| [8] | Mcleod K, Leslie H. Ecosystem-Based Management for the Oceans. Cambridge: Cambridge University Press, 2009. |
| [9] | 丘建文, 吴宝铃. 黄海渔业: 从单种和多种资源管理走向生态系统水平的管理. 自然资源学报, 1995, 10(3): 259-266. |
| [10] | 苏纪兰, 唐启升. 我国海洋生态系统基础研究的发展-国际趋势和国内需求. 地球科学进展, 2005, 20(2): 139-143. |
| [11] | 唐启升, 苏纪兰, 孙松, 张经, 黄大吉, 金显仕, 仝龄. 中国近海生态系统动力学研究进展. 地球科学进展, 2005, 20(12): 1288-1299. |
| [12] | 邱永松. 南海北部大陆架鱼类群落的区域性变化. 水产学报, 1988, 12(4): 303-313. |
| [13] | 邓景耀, 孟田湘, 任胜民, 邱显寅, 朱建元. 渤海鱼类种类组成及数量分布. 海洋水产研究, 1988, 9(1): 11-11. |
| [14] | 李圣法, 程家骅, 严利平. 东海大陆架鱼类群落的空间结构. 生态学报, 2007, 27(11): 4377-4386. |
| [15] | 刘勇, 程家骅. 东海、黄海底层鱼类数量分布季节变化的因子分析. 海洋学报, 2008, 30(4): 123-130. |
| [16] | 俞存根, 宋海棠, 姚光展. 东海蟹类群落结构特征的研究. 海洋与湖沼, 2005, 36(3): 213-220. |
| [17] | 宋海棠, 丁天明, 徐开达. 东海头足类的数量分布与可持续利用. 中国海洋大学学报, 2008, 38(6): 911-915. |
| [18] | 薛利建, 卢占晖. 东海中部海域蟹类群落结构及其多样性研究. 南方水产科学, 2011, 7(6): 66-71. |
| [19] | 郑元甲, 陈雪忠, 程家骅, 王云龙, 沈新强, 陈卫忠, 李长松. 东海大陆架生物资源与环境. 上海: 上海科学技术出版社, 2003: 767-790. |
| [20] | Sokal R R, Sneath P H A. Principles of Numerical Taxonomy. San Francisco: W. H. Freeman & Co Ltd., 1963. |
| [21] | Akima H. A method of bivariate interpolation and smooth surface fitting for irregularly distributed data points. ACM Transactions on Mathematical Software (TOMS), 1978, 4(2): 148-159. |
| [22] | Akima H. Algorithm 761: scattered-data surface fitting that has the accuracy of a cubic polynomial. ACM Transactions on Mathematical Software (TOMS), 1996, 22(3): 362-371. |
| [23] | Franke R. Scattered data interpolation: tests of some methods. Mathematics of Computation, 1982, 38(157): 181-200. |
| [24] | R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing, 2013 [2013-6-20]. http://www.R-project.org/. |
| [25] | 王斌会. 多元统计分析及 R 语言建模. 广州: 暨南大学出版社, 2010: 131-149. |
| [26] | Lucas A, Jasson S. Using amap and ctc Packages for Huge Clustering. The Newsletter of the R Project Volume 6/5, 2006, 1(3): 58-60. |
| [27] | 孙儒泳, 李博, 诸葛阳, 尚玉昌. 普通生态学. 北京: 高等教育出版社, 1993: 104-112. |
| [28] | 沈国英, 施并章. 海洋生态学. 厦门: 厦门大学出版社, 1990: 133-140. |
| [29] | 奥德姆, 巴雷特. 生态学基础. 陆健健, 王伟, 王天慧, 何文珊, 李秀珍, 译. 5版. 北京: 高等教育出版社, 2009: 281-284. |
| [30] | 刘勇, 程家骅, 陆健健. 利用Bootstrap统计方法探讨体长-体重关系中的幂指数对小黄鱼产卵期的指示作用. 渔业科学进展, 2011, 32(2): 1-6. |
| [31] | 罗秉征, 卢继武, 黄颂芳. 带鱼不同出生季节的鉴别. 海洋与湖沼, 1982, 13(5): 451-459. |
| [32] | 罗秉征. 东海北部带鱼性成熟的研究: 雄鱼成熟过程与特征. 海洋与湖沼, 1997, 28(6): 561-566. |
| [33] | 梁剑. 海上治安形势不容乐观. 中国水产, 2001, (3): 29-29. |
| [34] | 徐晓明. 海上治安现状及建立海上治安防控体系的设想. 科协论坛, 2010, (6): 135-136. |