 2015, Vol. 35
2015, Vol. 35文章信息
- 黄祥童, 王绍先, 黄炳军, 尹航, 崔凯峰, 赵伟, 范宇光, 顾德峰
- HUANG Xiangtong, WANG Shaoxian, HUANG Bingjun, YIN Hang, CUI Kaifeng, ZHAO Wei, FAN Yuguang, GU Defeng
- 珍稀植物对开蕨与其伴生物种的联结性及群落稳定性
- Analyses of community stability and inter-specific associations between the rare plant Phyllitis scolopendrium and its associated species
- 生态学报, 2015, 35(1): 80-90
- Acta Ecologica Sinica, 2015, 35(1): 80-90
- http://dx.doi.org/10.5846/stxb201404280842
-
文章历史
- 收稿日期:2014-04-28
- 修订日期:2014-11-14






2. 吉林农业大学园艺学院, 长春 130118
2. College of Horticulture, Jilin Agricultural University, Changchun 130118, China
群落环境异质性差异导致物种空间分布的变化,不同物种间相互影响、相互作用所形成的有机联系是种间联结的反映[1, 2]。是群落形成和演化的基础[3]。种群间这种联结和竞争又是种群自身调节演化到群落稳定与否的体现形式。群落的稳定程度最终是以群落中各种群的变化作为承受其结果的表达[4]。
近些年,国内外学者基于种间联结性和群落的稳定性方面进行了卓有成效的研究。如阳含熙[5]、蒋有绪等[6]在种间相关测定技术、种间关联理论和应用等方面做了许多开创性工作。王伯荪[7]基于2×2联列表,应用联结系数、共同出现百分率、点相关系数、X2统计量等,测定了鼎湖山厚壳桂(Cryptocarya chinensis) 群落主要种群的种间联结性。此后,许多学者对珍稀植物或重要模式植物种群进行了研究。如胡理乐、曹培健、林长松、付方林、方彦等人[8, 9, 10, 11, 12]分别对毛柄小勾儿茶(Berchemiella wilsonii var.pubipetiolata)、福建柏(Fokienia hodginsii)、十齿花(Dipentodon sinicus)、毛红椿(Toona ciliates var.pubescens)、野生早樱(Cerasus subhirtella var. ascendens)研究显示:珍稀植物种群与绝大多数种群无关联或显著关联的种群极少[8, 9];即使正联结显著,但呈极显著和显著关联种对比率很低[10];或多物种总体为显著正相关,但与优势种间联结性不显著[11],或总体呈负关联[12]。而肖宜安、陶冶[13, 14]等人分别对长柄双花木(Disanthus cercidifolius. var.longipes)、拟南芥(Arabidopsis thaliana)群落研究得出总体表现出正关联[13];与群落主要优势种关系密切[14],并提出二种群处于比较稳定的阶段。在群落稳定性研究上,目前存在许多异议,有诸多理论[15, 16, 17]和研究方法[18, 19, 20]。但最为经典的是M.Godron提出的贡献定律法(Contribution law)[21]。李育中[22]应用该方法对J.Poissonet等1982年测定的内蒙古天然羊草草原群落稳定性及人工羊草草地的稳定性进行了验证。郑元润[23]对该方法又进行了改进并首次应用于森林群落,并与马尔柯夫链转移概率演替模型对比,结果更为直观与可靠。毕晓丽[24]应用该方法对珍稀植物观光木(Tsoongiodendron odorum)、沉水樟(Cinnamomum micranthum)、罗浮栲(Castanopsis fabri)群落的乔、灌木层及群落总体稳定性分析得出3个群落均为不稳定群落。上述结果无论从种间联结性还是群落稳定性的研究,对珍稀植物在群落中的地位评价、制定优先保护策略等方面都有重要意义。
长白山地处欧亚大陆生物链条的最东端,垂直分布从北温带到极地几千公里自然植被带。是欧亚大陆北部特别是东北亚地区的典型代表。长白山区分布国家级珍稀濒危植物28种[25]。
对开蕨(Phyllitis scolopendrium) 为多年生常绿中型陆生蕨类植物。是中国新记录种[26],被列为国家Ⅱ级保护植物[27, 28]。间断分布于长白山南坡及西南坡,集中产于临江市(花山镇)、集安市(阳岔乡)、长白县(宝泉山乡)等市县[29],数量极其稀少。临江区域发现的对开蕨“不超过50棵”[30]。常小面积斑块状生于山地落叶阔叶林下。喜生于岩石风化的酸性壤土中,要求较高的空气湿度,耐寒性极强,具有较高的绿化观赏、药用和科学研究价值。
自1980年对开蕨首次在中国发现以来,有关对开蕨的研究主要集中在组织结构解剖及配子体发育[31, 32, 33, 34]、孢子育苗及引种栽培[35, 36, 37, 38]、组培扩繁[39, 40]及生理适应性[41, 42, 43, 44, 45, 46]等方面。关于对开蕨种群联结性、群落稳定性研究尚未见报道。
本文采用种群联结性和群落稳定性相结合的分析方法,对对开蕨进行种群现状分析,目的是确定对开蕨与伴生物种的联结关系和在群落中地位,预测群落结构变化及演替趋势。为制定珍稀植物的优先保护策略、种群恢复与重建等提供理论基础。
1 研究方法 1.1 研究区概况研究地点位于吉林省东南部的临江市,该市地处长白山西南麓的鸭绿江北岸,南与朝鲜民主主义人民共和国隔江相望。东经126°11′—127°35′,北纬41°27′—42°04′。属中温带大陆性季风气候,为温润森林区。年均气温4.7℃,7月均温22.3℃,≥10℃活动积温2759.7℃;年均降水量856.5mm,年均蒸发量1033.4mm,年相对湿度71%。由于其独特的地理位置,生物种类多样而丰富。分布此区域的对开蕨生于海拔720—1008m的落叶次生阔叶杂木林下,群落的物种多样性较低。由于多年的皆伐和后期的择伐构成群落的乔木以遗留乔木和个体矮小的亚乔木为主,主要乔木有暴马丁香(Syringa reticulata var. amurensis)、紫花槭(Acer pseudo-sieboldianum)、小楷槭(A. komarovii)、风桦(Betula costata )、裂叶榆(Ulmus laciniata )、千金榆(Carpinus cordata)、水榆花楸(Sorbus alnifolia)等;灌木有凤城卫矛(Euonymus maximowiczianus)、瓜木(Alangium platanifolium)、东北溲疏(Deutzia parviflora var. amurensis)、长白茶藨(Ribes komarovii)、东北茶藨(R.mandshuricum)等;草本植物以三叉耳蕨(Polystichum tripteron )、粗茎鳞毛蕨(Dryopteris crassirhizoma)、尖齿凤丫蕨(Coniogramme affinis)、野芝麻(Lamium barbatum)、山茄子(Brachybotrys paridiformis)、大叶芹(Pimpinella brachycarpa)等。对开蕨常分布于坡度较大的风化碎石塘上,林分郁闭度在0.7左右的西北坡,土壤有效钾为361.75—387.31 mg/kg、有效磷为99.39—108.23 mg/kg的腐殖质壤土上。
1.2 样地设置与调查在对开蕨分布片区沿等高线,按南北方向设置20m×20m样地,用相邻格子法区划成16个5m×5m样方。以样方为单元,对乔木、灌木、草本和更新层分别调查。乔木层主要调查样方内所有胸径>1.0cm乔木种类、胸径、树高、活枝下高、冠幅及相对坐标。灌木和草本分别在5m×5m样方对角线方向设置3个2m×2m的小样方,调查植物种类、高度、冠径、多度、物候及生长势。
2 012年8月24—25日调查时,仅发现两小片对开蕨原始群落(未见散生)。样地情况见表 1。
|
群落号
Plot No. | 调查地点
Site | 经纬度
Longitude and latitude (东经/北纬) | 海拔
Altitude/m | 坡度
Gradient | 坡向
Slope aspect | 郁闭度
Canopy density | 树种组成(断面积)
Consist from trees |
| 01 | 吉林省临江市花山镇四道洋岔 | 126°56′03.49″
41°53′23.54″ | 1008m | 28° | 西北 | 0.78 | 2风桦(Betula costata)2榆(Ulmus spp.)1胡(Juglans mandshurica)1椴(Tilia amurensis)1水榆(Sorbus alnifolia)1色
(Acer mono)+ 花楸(Sorbus pohuashanensis)-槭(Acer spp.) |
| 02 | 吉林省临江市花山镇五道沟水库风景区 | 127°09′29.9″
41°43′07.16″ | 729m | 56° | 西北 | 0.72 | 4风桦(Betula costata)2紫椴(Tilia amurensis)2色木槭(Acer mono)+榆树(Ulmus spp.)-槭(Acer spp.) |
以X2检验为基础,先用方差比率法(VR)验证整个群落的总体关联性,再以X2统计量等确定物种间联结性指标[47, 48]。
(1)群落总体联结性
多植物种间的总体关联性检验采用D.Schluter提出的方差比率(VR)进行检验[49]:


式中,S为总物种数,N为总样方数,Tj为样方j内出现的物种总数,ni为物种i出现的样方数,t为样方中种群的平均数。
VR值为群落内植物种间的总体联结指数。VR=1,即所有种间无关联;VR>1表示物种间总体表现为正关联,VR<1表示物种间总体表现为负关联。
采用统计量W来检验VR值偏移1的显著程度,W=N×VR。若种间关联不显著,则W落入X(0.95,N)2<W<X(0.05,N)2界限内的概率有90%。
X2检验 把成对物种在取样中存在与不存在排列成2×2联列表。根据联列表的X2值确定对开蕨与其伴生物种联结性。由于取样的非连续性,X2值用Yates的连续校正公式计算[50, 51]:
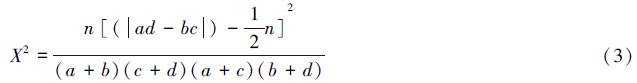
若Xt2<3.841(P>0.05),则表示种对联结不显著;若3.841<Xt2<6.635(0.01<P<0.05),则表示种对联结显著;若Xt2>6.635(P<0.01),则表示种对联结极显著。当ad>bc时确定为正联结,反之为负联结[47, 52]。
(2)对开蕨与伴生物种关联性测度
X2检验仅得出了种间联结性显著与否的结论,而关联不显著的种对仍具有一定程度的联结,需进一步测定联结程度即关联度加以反映。本文采用点相关系数(PCC)、联结系数(AC)、Ochiai指数(OI)和共同出现百分率(PC)。
点相关系数:
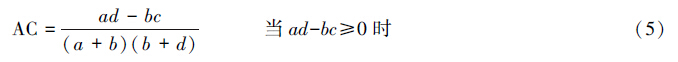
联结系数:
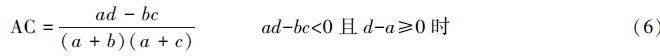
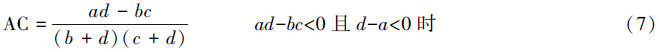
OI指数:

共同出现百分率:

式中,a为两个物种都存在样方数;b为B物种存在而A物种不存在样方数;c为A种存在而B种不存在样方数;d为两个物种都不存在样方数;n为样方总数。
在计算连接系数AC时,为避免a,b或d为0而导致AC=1或AC=-1出现无法比较关联程度的状况,把a、b、c、d凡是为0的都加权为1。
1.3.2 对开蕨群落稳定性采用贡献定律法,首先把样地中所有植物的频度由大到小的顺序排列,换算成相对频度后逐步累积;然后将样地内植物种类的总和取倒数,按着植物种类排列顺序也逐步对应累积;再将植物种类倒数百分率同累积相对频度一一对应的数据,建立模糊散点平滑曲线模型(或以植物种类倒数百分率为x轴,累积相对频度为y轴,画出平滑曲线,由图可以看出百分之多少的种类占有多大的累积相对频度)。然后再与直线y=100-x相交,交点即为所求群落的稳定性比值。一般地,稳定群落的种类倒数百分率与累积相对频度比值为20/80是群落的稳定点。计算出的稳定性比值越接近20/80,群落就越稳定。
平滑曲线模拟模型为:

直线方程为:

将(11)代入(10)得:

交点坐标为:
2 结果与分析 2.1 对开蕨群落总体关联性
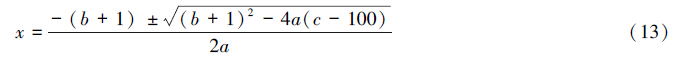

计算统计结果见表 2。由表可知,01群落和02群落的总体关联性,其方差比率VR分别为1.6340和1.4288,VR值均>1。说明种群间均表现出净的正关联。但通过统计量W来检验VR值的偏离程度,W值均落入X0.95(N)2与X0.05(N)2之间,意味着有90%的可靠性说明VR偏离1不显著。同时,01群落总体关联性高于02群落,反映出02群落形成晚于01群落,均为演替初期阶段,群落成熟和稳定性较差。由此看出,群落总体关联程度亦反映了群落的稳定性。
|
群落
Community | 方差比率
Variance ratio | 检验统计量W
Test statistics W | X2临界值
X2 Critical value (X0.95,N2,X0.05,N2) | 测定结果
Results |
| 海拔1008m的01群落 | 1.6340 | 26.1440 | (7.962,26.296) | 总体正关联,但不显著 |
| 海拔729m的02群落 | 1.4288 | 22.8608 | (7.962,26.297) | 总体正关联,但不显著 |
在群落总体关联性检验的基础上,以2×2联列表为基础,分别计算对开蕨与乔木、灌木、草本层的每一物种的统计量X2、点相关系数(PCC)、联结系数(AC)、OI指数(OI)、成对出现的百分率(PC)来分析对开蕨与伴生物种关联程度。
2.2.1 对开蕨与乔木物种的联结对开蕨与乔木物种的联结结果见表 3。从表 3可知:对开蕨与乔木种对统计量Xt2<3.841(P>0.05),说明对开蕨与所有乔木联结不显著。但水榆花楸和色木槭(Acer mono)Xt2>1.5,有负联结倾向。从AC、PCC结果看,与对开蕨正联结的种对有6种,负联结的种对有10种。而 AC >0.5的只有3种。其中千金榆、暴马丁香正联结,水榆花楸负联结。0.3<|AC|<0.5有3种,均为槭树(Acer spp.);0.1<|AC|<0.3有10种。并且|PCC|都<0.4,尤其是正联结的|PCC|均<0.2,从此也印证了绝大部分乔木与对开蕨无关联或稍倾向负联结。
|
树种
Tree species | 联结性测度指标Association indices | ||||
| X2 | PCC | AC | OI | PC | |
| 暴马丁香Syringa reticulata var. amurensis | 0.4023 | 0.1937 | 0.5429 | 0.4835 | 0.2609 |
| 千金榆Carpinus cordata | 0.4023 | 0.1937 | 0.5429 | 0.4835 | 0.2609 |
| 裂叶榆Ulmus laciniata | 0.2741 | 0.1631 | 0.2846 | 0.6052 | 0.4167 |
| 青楷槭Acer tegmentosum | 0.1343 | 0.1511 | 0.4667 | 0.4352 | 0.2174 |
| 小楷槭A. komarovii | 0.0043 | 0.1044 | 0.3600 | 0.3814 | 0.1739 |
| 紫花槭A. pseudo-sieboldianum | 0.0831 | 0.0510 | 0.2000 | 0.3198 | 0.1304 |
| 水榆花楸Sorbus alnifolia | 2.0779 | -0.3568 | -0.6364 | 0.1066 | 0.0400 |
| 色木槭A. mono | 1.6835 | -0.3028 | -0.3333 | 0.2843 | 0.1481 |
| 风桦Betula costata | 0.3730 | -0.1943 | -0.2727 | 0.2611 | 0.1200 |
| 东北槭A. mandshuricum | 0.0831 | -0.1325 | -0.1688 | 0.3223 | 0.1600 |
| 稠李Padus racemosa | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 红松Pinus koraiensis | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 秋子梨Pyrus ussuriensis | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 胡桃楸Juglans mandshurica | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 花楸树Sorbus pohuashanensis | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 紫椴Tilia amurensis | 0.0000 | -0.0778 | -0.0909 | 0.3769 | 0.2000 |
同时,表 3中还反映原长白山地带性顶级植被中的红松(Pinus koraiensis)、紫椴(Tilia amurensis)、胡桃楸(Juglans mandshurica)、色木槭、风桦都处在与对开蕨负关联状态,但关联系数较低,并没有达到显著性程度。为今后引种回归到红松针阔叶混交林提供理论依据。
2.2.2 对开蕨与灌木物种的联结表 4是计算出的灌木联结性,从表看出:种对统计量Xt2值,只有对开蕨与凤城卫矛(Euonymus maximowiczianus)的卡方值Xt2=5.9345>3.841,达到正关联的显著水平。黄花忍冬(Lonicera chrysantha)和狗枣猕猴桃(Actinidia kolomikta)Xt2值分别为3.1030和1.9066,与对开蕨有负联结的倾向;从AC、PCC结果看,与对开蕨正联结的种对有6种,负联结的种对有10种。|A C|>0.5的只有1种,即凤城卫矛,其|PCC|、|A C|、OI、PC值均排在第一位。而0.3<|A C|<0.5的有4种,正联结分别是瓜木、东北溲疏;负联结的分别是黄花忍冬、狗枣猕猴桃。0.1<|A C|<0.3有11种。其中,以刺五加(Acanthopanax senticosus)、毛接骨木(Sambucus williamsii var. miquelii)、东北山梅花(Philadelphus schrenkii)、五味子(Schisandra chinensis)、李叶溲疏(D.hamata)、毛榛(Corylus mandshurica)、黄花忍冬及鸡树条(Viburnum opulus var. calvescens)为长白山地带顶级群落——红松针阔叶混交林中代表种,均为负联结,但没有达到显著性程度。|PCC|排序结果与|A C|相同。
|
灌木物种
Shrub species | 联结性测度指标Association indices | ||||
| X2 | PCC | AC | OI | PC | |
| 凤城卫矛Euonymus maximowiczianus | 5.9345* | 0.4982 | 0.7867 | 0.7707 | 0.6087 |
| 瓜木Alangium platanifolium | 0.1343 | 0.1511 | 0.4667 | 0.4352 | 0.2174 |
| 东北溲疏Deutzia parviflora var. amurensis | 0.0043 | 0.1044 | 0.3600 | 0.3814 | 0.1739 |
| 长白茶藨Ribes komarovii | 0.0831 | 0.0510 | 0.0909 | 0.3198 | 0.1304 |
| 东北茶藨R. mandshuricum | 0.0831 | 0.0306 | 0.0857 | 0.4029 | 0.2083 |
| 软枣猕猴桃Actinidia arguta | 0.0831 | 0.0306 | 0.0857 | 0.4029 | 0.2083 |
| 刺五加Acanthopanax senticosus | 0.0703 | -0.0281 | -0.0261 | 0.4264 | 0.2400 |
| 毛接骨木Sambucus williamsii var. miquelii | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 东北山梅花Philadelphus schrenkii | 0.0831 | -0.0306 | -0.0857 | 0.7249 | 0.5667 |
| 五味子Schisandra chinensis | 0.0025 | -0.0798 | -0.0857 | 0.4500 | 0.2692 |
| 李叶溲疏D. hamata | 0.1619 | -0.1423 | -0.1391 | 0.3956 | 0.2222 |
| 毛榛Corylus mandshurica | 0.1351 | -0.1449 | -0.1883 | 0.3152 | 0.1538 |
| 鸡树条Viburnum opulus var. calvescens | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 少花瘤枝卫矛E. verrucosus var. pauciflorus | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 狗枣猕猴桃Actinidia kolomikta | 1.9006 | -0.3133 | -0.3600 | 0.3693 | 0.2143 |
| 黄花忍冬Lonicera chrysantha | 3.1030 | -0.3892 | -0.4545 | 0.2261 | 0.1111 |
对于草本植物而言,对开蕨与其处于同一层次的草本植物关系较乔木、灌木层更为复杂和紧密(表 5,表 6)。
|
草本物种
Herb species | 联结性测度指标Association indices | ||||
| X2 | PCC | AC | OI | PC | |
| 三叉耳蕨Polystichum tripteron | 10.6415* * | 0.6476 | 0.6952 | 0.8840 | 0.7917 |
| 粗茎鳞毛蕨Dryopteris crassirhizoma | 4.5515* | 0.4587 | 0.3600 | 0.8528 | 0.7407 |
| 尖齿凤丫蕨Coniogramme affinis | 0.4023 | 0.1937 | 0.5429 | 0.4835 | 0.2609 |
| 毛金腰Chrysosplenium pilosum | 0.4023 | 0.1937 | 0.5429 | 0.4835 | 0.2609 |
| 掌叶铁线蕨Adiantum pedatum | 0.2645 | 0.1636 | 0.3600 | 0.5394 | 0.3333 |
| 木贼Equisetum hyemale | 0.0831 | 0.0510 | 0.2000 | 0.3198 | 0.1304 |
| 类叶升麻Actaea asiatica | 0.0831 | 0.0510 | 0.2000 | 0.3198 | 0.1304 |
| 东北蛾眉蕨Lunathyrium pycnosorum | 0.0831 | 0.0510 | 0.2000 | 0.3198 | 0.1304 |
| 布朗耳蕨Polystichum braunii | 0.0831 | 0.0510 | 0.2000 | 0.3198 | 0.1304 |
| 假升麻Aruncus sylvester | 0.0043 | 0.1044 | 0.3600 | 0.3814 | 0.1739 |
|
草本物种
Herb species | 联结性测度指标Association indices | ||||
| X2 | PCC | AC | OI | PC | |
| 野芝麻Lamium barbatum | 4.5515* | -0.4587 | -0.5844 | 0.1612 | 0.0741 |
| 香车叶草Galium odoratum | 2.7429 | -0.3637 | -0.3905 | 0.3214 | 0.1786 |
| 碎叶山芹Ostericum grosseserratum | 2.0779 | -0.3568 | -0.6364 | 0.1066 | 0.0400 |
| 水金凤Impatiens noli-tangere | 1.2800 | -0.2727 | -0.2727 | 0.3371 | 0.1852 |
| 东北南星Arisaema amurense | 0.8237 | -0.2280 | -0.3600 | 0.5171 | 0.3448 |
| 山茄子Brachybotrys paridiformis | 0.7758 | -0.2335 | -0.2727 | 0.3015 | 0.1538 |
| 珠芽艾麻Laportea bulbifera | 0.7279 | -0.2218 | -0.2381 | 0.3857 | 0.2222 |
| 五叶黄精Polygonatum acuminatifolium | 0.5417 | -0.2458 | -0.5152 | 0.1231 | 0.0417 |
| 龙常草Diarrhena manshurica | 0.5417 | -0.2458 | -0.5152 | 0.1231 | 0.0417 |
| 直酢浆草Oxalis corniculata var. stricta | 0.5417 | -0.2458 | -0.5152 | 0.1231 | 0.0417 |
| 大叶芹Pimpinella brachycarpa | 0.3401 | -0.1781 | -0.1919 | 0.3553 | 0.1923 |
| 山荷叶Astilboides tabularis | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 透骨草Phryma leptostachya subsp. asiatica | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 茜草Rubia cordifolia | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 光岩蕨Woodsia glabella | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 蔓假繁缕Pseudostellaria davidii | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 白花碎米荠Cardamine leucantha | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 虎耳鳞毛蕨Dryopteris saxifraga | 0.3277 | -0.0145 | -0.0303 | 0.2462 | 0.0870 |
| 荨麻叶龙头草Meehania urticifolia | 0.0831 | -0.0306 | -0.0857 | 0.5217 | 0.3500 |
| 大叶蟹甲草Parasenecio firmus | 0.0831 | -0.1529 | -0.2727 | 0.2132 | 0.0833 |
| 细叶鳞毛蕨Dryopteris woodsiisora | 0.0831 | -0.1529 | -0.2727 | 0.2132 | 0.0833 |
| 过山蕨Camptosorus sibiricus | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 荫地冷水花Pilea pumila | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 毛叶沼泽蕨Thelypteris palustris var. pubescens | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 深山堇菜Viola selkirkii | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 岩蕨Woodsia ilvensis | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
|
乌苏里黄芩
Scutellaria pekinensis var. ussuriensis | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
| 香根芹Osmorhiza aristata | 0.0388 | -0.1044 | -0.2727 | 0.1508 | 0.0435 |
结果显示:对开蕨与三叉耳蕨的Xt2=10.6415>6.635(P<0.01),达到极显著的正联结,并且PCC、AC、OI指数和PC值均最高。与粗茎鳞毛蕨和野芝麻的Xt2值均在3.841<Xt2<6.635(0.01<P<0.05)之间,已达到显著水平,其中与粗茎鳞毛蕨显示为正联结,与野芝麻为负联结。
在正联结中,对开蕨虽然与尖齿凤丫蕨、毛金腰(Chrysosplenium pilosum)和掌叶铁线蕨(Adiantum pedatum)没有达到显著性水平,但其AC、O I指数和PC值均较高。说明这3种植物与对开蕨表现出相似的喜阴喜湿生境需求。而粗茎鳞毛蕨虽达到显著的正联结,但AC值却不如PCC、OI指数和PC值高;在负联结中,尽管与野芝麻达到显著负联结水平,但其AC、OI指数和PC的值却不是最高,反映出对开蕨与草本植物联结的复杂性。
表 5和表 6显示的正、负联结种对分别为10种和28种。而|A C|>0.5的只有8种,其中正联结3种,负联结5种;而0.3<|A C|<0.5的有5种,其中正联结2种,负联结3种;0.1<|A C|<0.3的有25种,占草本植物总数的65.8%,这些物种可以认为无关联。说明对开蕨在群落中都具有很大的独立性或随机性,它只是群落的伴生种或偶见种。
2.3 对开蕨群落的稳定性根据M. Godron稳定性测定方法,将群落所有植物进行累计并绘图。得到不同海拔下的对开蕨群落稳定性图解(图 1)。图中显示:海拔1008m的01样地整体群落的种类倒数百分率与累积相对频度比值为35/65,海拔729m的02样地其整体群落的种类倒数百分率与累积相对频度比值为34/66,均远离20/80。

|
| 图 1 对开蕨群落稳定性图 Fig.1 Community stability graph of P. scolopendrium |
为精确判定每群落各层次的稳定程度,各群落按乔木层、灌木层、草本层分别绘图(图略),并以二次函数模拟计算与y=100-x的解,结果见表 7。
|
群落编号
Communitie | 曲线类型
Type of curves | 决定系数(R2)
Determination coefficient | P | 交点坐标Coorlinate | 结果
Results | ||
| x | y | ||||||
| 01群落 | 群落整体 | y=-0.0117x2+2.026x+9.3304 | 0.9846 | P<0.01 | 34.6 | 65.4 | 不稳定 |
| 乔木层 | y=-0.0093x2+1.837x+6.8757 | 0.9967 | P<0.01 | 37.4 | 62.6 | 不稳定 | |
| 灌木层 | y=-0.0103x2+1.923x+7.1329 | 0.9908 | P<0.01 | 36.5 | 63.5 | 不稳定 | |
| 草本层 | y=-0.0124x2+2.152x+3.0813 | 0.9896 | P<0.01 | 35.8 | 64.2 | 不稳定 | |
| 02群落 | 群落整体 | y=-0.0103x2+1.847x+14.772 | 0.9882 | P<0.01 | 34.2 | 65.8 | 不稳定 |
| 乔木层 | y=-0.0071x2+1.662x+5.6769 | 0.9986 | P<0.01 | 39.6 | 60.4 | 不稳定 | |
| 灌木层 | y=-0.0079x2+1.620x+16.011 | 0.9966 | P<0.01 | 35.9 | 64.1 | 不稳定 | |
| 草本层 | y=-0.0114x2+1.892x+18.865 | 0.9709 | P<0.01 | 32.1 | 67.9 | 不稳定 | |
图示与精确计算交点坐标结果显示:两处对开蕨群落无论群落整体,还是各层次均处于不稳定状态。各层次稳定性排序均为草本层>灌木层>乔木层。
3 结论与讨论种群联结性是稳定性基础,群落稳定性是种群联结性的表达形式。种间联结指标是对一定时期内群落组成物种之间相互关系的静态描述,随群落演替阶段的变化而变化。一般来说,处于演替顶级的植物群落,群落内各种群间联结紧密,多物种联结的方差比率VR远大于1,且显著;呈显著正联结的种对数量较多,显著负联结或负关联的种对少。从非顶级群落至顶级群落,种群间由负关联至多物种的正联结逐渐加大加强,最终达到稳定的平衡状态。因此,种间联结性又与群落的稳定性相关。
在一定意义上来说,群落稳定性是种群联结性的体现形式,群落的稳定程度最终是以群落中各种群的变化承受其结果的表达。从种群联结性到群落稳定性是一个复杂的生态过程。在研究对开蕨群落生存状况评价时,将种群联结性和群落稳定性放在一起,二者得出的结论可以相互印证,有很大的实践意义。
(1)调查显示:吉林省临江区域自然分布的对开蕨种群为小片状聚集分布,数量较少,局部大尺度上分布频度极低,为偶见种。与对开蕨伴生的植物共有70种,其中乔木16种,灌木16种,草本植物38种。
(2)卡方检验表明:对开蕨群落总体表现出正关联(VR>1),但没有达到显著水平(W<X20.05(N))。对开蕨种群与乔木、灌木联结性不如与草本联结复杂和紧密。与乔木联结种对均不显著(X2<3.841);与灌木正联结的种对以凤城卫矛达到显著正联结(X2>3.841);与草本植物三叉耳蕨达到极显著的正联结(X2>6.635),与粗茎鳞毛蕨为显著正联结,与野芝麻为显著负联结(3.841<X2<6.635)。验证了该群落种对间独立性较强、相关性较弱的特点。说明对开蕨群落尚未发育到成熟阶段,群落结构和功能还不稳定,群落组成物种处于动态调节中,没有形成多物种共存复杂而稳定的群落。
(3)联结系数和点相关系数分析表明:对开蕨与乔木正关联的种占乔木总数的37.5%,而负关联种占62.5%,且均不显著;与灌木正关联种占灌木总数的37.5%,但显著正联结的种对只有1种;与草本植物正关联种占草本总数的26.32%,显著正联结的种对有2种,显著负联结种对有1种。总体而言,无显著关联种占所有植物总数的94.29%。由此看出:对开蕨与大多数植物种群关联不显著或显著关联的种群极少,种间联结关系较为松散而趋向于相互独立,有很大的随机性。胡理乐等在研究濒危植物毛柄小勾儿茶的种间联结性时也得到类似的结果。
种间联结测定值在一定程度上衡量了物种间的相互关系和植物对环境因子的适应程度。正联结体现了物种利用资源的相似性和生态位的重叠性,说明它们之间具有相似的生境要求。对开蕨与乔木联结不显著,说明对开蕨与乔木树种间没有直接的内在联系。灌木和草本对对开蕨的生长和存活有直接的影响,如凤城卫矛、三叉耳蕨、粗茎鳞毛蕨都有相似的生境需求,是对开蕨有益的伴生种。
(4)群落稳定性分析表明:对开蕨整体群落的种类倒数百分率与累积相对频度比值分别为35/65和34/66,均远离20/80这一稳定点,群落整体稳定性较差,生态脆弱。
(5)杜道林认为,随着群落演替的进行,群落结构及其种类组成将逐渐趋于稳定,种间关系也将趋于正相关以求得多物种共存[53]。从演替阶段上来说吉林省临江市分布的对开蕨还处于初期或中期,五道沟分布的对开蕨群落更为年轻,样地内对开蕨有91株,幼苗数量较多;四道洋岔样地内有对开蕨31株,均为大苗。随着群落的发展,目的物种对开蕨很难说不被其它物种所取代。
(6)对开蕨分布区狭窄,自然繁殖力较弱,种群面临灭绝的危险,目前尚无保护措施。建议有关部门进行详细的资源考察,摸清现存数量。对主要产地严加保护,禁止采挖。同时,在就地保护基础上积极开展人工辅助措施促进其种群更新和扩繁。如采取保护与之正联结较强的物种,削弱或控制负联结物种等措施来保护对开蕨的生存环境;积极开展种群扩繁研究,采用组织培养等手段扩大不同群体的幼苗数量,最终将其回归和重新引入自然为目标。顾德峰等已将对开蕨组培成功,部分苗木已经回归到吉林省的6个试验小区,进行回归和重引入试验,期待这项意义重大的工作取得突破性成果。
| [1] | 王伯荪, 李明光, 彭少麟. 植物种群学. 广州: 广东高等教育出版社, 1989. |
| [2] | Greig-Smith P. Quantitative Plant Ecology. 3rd ed. Oxford: Blackwell Scientific Publications, 1983. |
| [3] | 刘萍萍, 程积民. 植物种间联结关系的研究. 水土保持研究, 2000, 7(2): 179-184. |
| [4] | 彭少麟. 森林群落稳定性与动态测度-年龄结构分析. 广西植物, 1987, 7(1): 67-72. |
| [5] | 阳含熙, 卢泽愚. 植物生态学的数量分类方法. 北京: 科学出版社, 1981. |
| [6] | 蒋有绪. 川西亚高山森林植被的区系、种间关联和群落排序的生态分析. 植物生态学与地植物学丛刊, 1982, 6(4): 281-301. |
| [7] | 王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究 Ⅰ. 种间联结的探讨与修正. 植物生态学与地植物学丛刊, 1985, 9(4): 274 -285. |
| [8] | 胡理乐, 江明喜, 党海山, 黄汉东. 从种间联结分析濒危植物毛柄小勾儿茶在群落中的地位. 植物生态学报, 2005, 29(2): 258-265. |
| [9] | 曹培健, 丁炳扬, 李伟成, 哀建国, 金孝锋. 凤阳山福建柏群落主要种群种间联结性研究. 浙江大学学报: 理学版, 2006, 33(6): 676-681. |
| [10] | 林长松, 李玉英, 左经会, 廖雯. 珍稀植物十齿花群落乔木优势种群种间联结性. 生态学杂志, 2008, 27(2): 178-184. |
| [11] | 付方林, 张露, 杨清培, 梁跃龙, 陶丹. 毛红椿天然林优势种群的种间联结性研究. 江西农业大学学报, 2007, 29(6): 982-987. |
| [12] | 方彦, 谢春平, 伊贤贵, 王贤荣. 野生早樱群落主要乔木种群种间联结性研究. 林业资源管理, 2008, (6): 50-54. |
| [13] | 肖宜安, 何平, 李晓红, 邓洪平, 时明芝. 长柄双花木分布群落中优势种群间联结性研究. 西南师范大学学报: 自然科学版, 2003, 28(6): 952-957. |
| [14] | 陶冶, 刘彤, 邱东, 李惠, 张荣. 天山西部果子沟地区拟南芥生存群落种间联结性分析. 干旱区地理, 2010, 33(6): 930-938. |
| [15] | 张继义, 赵哈林. 植被(植物群落)稳定性研究评述. 生态学杂志, 2003, 22(4): 42-48. |
| [16] | 徐坤, 谢应忠, 郑国琴. 植被稳定性研究进展. 宁夏农学院学报, 2004, 25(4): 59-66. |
| [17] | 党承林, 王崇云, 王宝荣, 李彦玲, 黄其明. 植物群落的演替与稳定性. 生态学杂志, 2002, 21(2): 30-35. |
| [18] | 张立敏, 陈斌, 李正跃. 应用中性理论分析局域群落中的物种多样性及稳定性. 生态学报, 2010, 30(6): 1556-1563. |
| [19] | 阳含熙, 潘愉德, 伍业钢. 长白山阔叶红松林马氏链模型. 生态学报, 1988, 8(3): 211-219. |
| [20] | 闫东锋, 王向阳, 杨喜田. 主成分分析法和稳定度指数法评价森林群落稳定性比较. 河南农业大学学报, 2011, 45(2): 166-171. |
| [21] | Godron M, Daget P, Poissonet J, Poissonet P. Some aspects of heterogeneity in grasslands of Cantal//Patil G P, Pielou E C, Waters W E. Statistical Ecology. Volume 3: Many Species Populations, Ecosystems, and Systems Analysis. University Park, PA: Pennsylvania State University Press, 1971: 397-415. |
| [22] | 李育中. 植物群落稳定性的一种测定方法. 中国草地, 1991, (2): 78-81. |
| [23] | 郑元润. 森林群落稳定性研究方法初探. 林业科学, 2000, 36(5): 28-32. |
| [24] | 毕晓丽, 洪伟, 吴承祯, 闫淑君, 封磊, 王新功. 珍稀植物群落多样性及稳定性分析. 福建林学院学报, 2003, 23(4): 301-304. |
| [25] | 周繇. 长白山区珍稀濒危植物的现状与保护. 浙江林学院学报, 2004, 21(3): 263-268. |
| [26] | 钱家驹. 对开蕨属首次在我国发现. 植物分类学报, 1980, 18(4): 482-483. |
| [27] | 中华人民共和国国务院. 国家重点保护野生植物名录(第一批). 植物杂志, 1999, (5): 4-11. |
| [28] | 傅立国. 中国珍稀濒危植物. 上海: 上海教育出版社, 1989: 1-3. |
| [29] | 刘保东, 时述武. 长白山的珍稀观赏植物——对开蕨. 中国野生植物, 1991, (4): 37-38. |
| [30] | 李玺. 临江国家级珍稀濒危植物调研与保护. 中国林业, 2012, (9): 32 |
| [31] | 古安根, 刘仪娴. 对开蕨属导管的发现. 植物学报, 1987, 29(4): 377-378. |
| [32] | 古安根, 汪矛. 对开蕨属次生维管组织的发现. 植物研究, 1989, 9(4): 87-88. |
| [33] | 刘保东, 包文美, 敖志文. 中国产对开蕨配子体发育的研究. 植物研究, 1991, 11(2): 93-96. |
| [34] | 刘保东. 对开蕨孢子育苗. 生命世界, 1993, (3): 27. |
| [35] | 王立军, 张友民, 钟岩. 东北对开蕨的解剖研究. 吉林农业大学学报, 1996, 18(增刊): 1-2. |
| [36] | 田洪, 王殿芝, 赵占英, 张宝国. 东北对开蕨的栽培. 人参研究, 1997, (2): 16-17. |
| [37] | 李彬. 东北对开蕨人工栽培技术. 中国林副特产, 2013, (3): 63-64. |
| [38] | 岳晓晶, 岳桦. 对开蕨在哈尔滨地区的繁殖技术研究. 黑龙江农业科学, 2010, (7): 81-83. |
| [39] | 顾德峰, 李东升, 王蕾, 齐广勋, 赵和祥, 董然. 东亚对开蕨离体快繁的研究. 园艺学报, 2008, 35(9): 1373-1376. |
| [40] | 顾德峰, 王蕾, 赵和祥, 董然. 东北对开蕨的组织培养. 植物生理学通讯, 2008, 44(5): 957-958. |
| [41] | 岳晓晶. 对开蕨光适应能力及繁殖技术研究[D]. 哈尔滨: 东北林业大学, 2010. |
| [42] | 岳桦, 岳晓晶. 对开蕨对遮荫处理的响应. 园艺学报, 2010, 37(9): 1517-1522. |
| [43] | 孙笑丛. 对开蕨在干旱胁迫下形态生理及哈尔滨露地生境生长发育节律特征[D]. 哈尔滨: 东北林业大学, 2011. |
| [44] | 岳桦, 吴妍, 姜丽颖. 不同栽培基质对对开蕨的影响. 黑龙江农业科学, 2011, (2): 66-68. |
| [45] | 岳桦, 孙笑丛. PEG渗透胁迫对对开蕨生理特性影响. 北方园艺, 2011, (1): 91-94. |
| [46] | 吴妍. 不同栽培基质及营养液对对开蕨的影响[D]. 哈尔滨: 东北林业大学, 2011. |
| [47] | 王乃江, 张文辉, 陆元昌, 范少辉, 王勇. 陕西子午岭森林植物群落种间联结性. 生态学报, 2010, 30(1): 67-78. |
| [48] | 张倩媚, 陈北光, 周国逸. 鼎湖山主要林型优势树种种间联结性的计算方法研究. 华南农业大学学报, 2006, 27(1): 79-83. |
| [49] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998-1005. |
| [50] | Ludwig J A, Reynolds J F. Statistical Ecology. New York: Academic Press, 1991. |
| [51] | 张金屯. 群落中物种多度格局的研究综述. 农村生态环境, 1997, 13(4): 48-54. |
| [52] | 郭志华, 卓正大, 陈洁, 吴梅凤. 庐山常绿阔叶、落叶阔叶混交林乔木种群种间联结性研究. 植物生态学报, 1997, 21(5): 424-432. |
| [53] | 杜道林, 刘玉成, 李睿. 缙云山亚热带栲树林优势种群间联结性研究. 植物生态学报, 1995, 19(2): 149-157. |