 2015, Vol. 35
2015, Vol. 35文章信息
- 张悦, 郭利平, 易雪梅, 曹伟, 王远遐, 吴培莉, 姬兰柱
- ZHANG Yue, GUO Liping, YI Xuemei, CAO Wei, WANG Yuanxia, WU Peili, JI Lanzhu
- 长白山北坡3个森林群落主要树种种间联结性
- Analysis of interspecific associations among major tree species in three forest communities on the north slope of Changbai Mountain
- 生态学报, 2015, 35(1): 106-115
- Acta Ecologica Sinica, 2015, 35(1): 106-115
- http://dx.doi.org/10.5846/stxb201403250548
-
文章历史
- 收稿日期:2014-03-25
- 修订日期:2014-10-31






2. 中国科学院大学, 北京 100049;
3. 中国科学院地理科学与资源研究所, 北京 100101;
4. 吉林省露水河林业局宏伟种子园, 白山 134506
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Institute of Geographic Sciences and Natural Resources Research, Beijing 100101, China;
4. Jilin Province, Lushuihe Forestry Bureau, Hongwei Seed Orchard, Baishan 134506, China
种间联结性是对一定时期内植物群落组成物种之间相互关系的静态描述[1],是以二元数据为基础,检验两个物种是否存在关联,表达的是一种定性关系[2, 3, 4]。种间联结通常是由群落演替过程中物种对环境的趋同性及其相互作用而引起的,它不仅是群落形成、演替的基础,也是划分群落的依据及物种间相互关系的表现形式之一[5]。不同物种间关联程度的测定,对于研究各物种之间的协作关系及其对环境的适应性和在群落中的分布、种群的进化以及群落的演替规律和分类都有重要意义[6, 7, 8]。
长白山是东亚大陆在我国境内唯一具有高山冻原的山地,随海拔高度的变化,呈现出垂直分布植被带[9, 10, 11],北坡的群落组成、结构沿海拔梯度的分布差异最为明显和完整[12]。长白山的重要用材树种主要有红松(Pinus koraiensis)、水曲柳(Fraxinus mandshurica)、紫椴(Tilia amurensis)、胡桃楸(Jugland mandshurica)、红皮云杉(Picea koraiensis)、长白鱼鳞云杉(Picea jezoensis)、臭冷杉(Abies nephrolepis)等,起到保持水土、涵养水源、调节小气候等作用[13, 14]。然而,20个世纪以来长白山原始天然林经历了3个阶段的过度采伐,残存的森林大部分已演变为天然次生林[15, 16, 17]。分析现存的原始阔叶红松林、云冷杉林和天然次生林的乔木种分类特征,了解树种间的关联性至关重要。以往的研究工作曾对长白山北坡锻树阔叶红松林群落、长白山北坡暗针叶林、长白山云冷杉针阔混交林的种间关联性、群落组成结构的特点及其沿海拔梯度的变化规律、两个演替阶段乔木的种间联结性等展开[7, 18, 19]。然而,同时分析长白山北坡3个植物群落,结合TWINSPAN分类、种间联结系数研究不同群落内乔木种的种间关联性尚未见报道。通过数量分类,可以认识长白山北坡海拔700 m到1700 m的3个植物群落的分布特征,对各群落中的主要乔木种进行种间联结性分析,可以反映出群落生境差异以及物种生态位的重叠与分离,为长白山地区植被的经营管理、植被恢复和生物多样性保护提供依据。
1 研究方法 1.1 野外调查与样方设置野外调查于2010、2011年进行,研究地点在长白山北坡安图县,在海拔700—1700m之间,共设置35个大小为30m×30m的样方,每个样方分为36个5m×5m的小样方,在每个小样方中对树高≥1.3m的树木,测定胸径和树高;对树高<1.3m的乔木,记录种名及株数。同时测定各样方的经纬度、海拔、坡度、坡向和坡位。统计各样方中出现的树种数,树种出现的样方数以及种对间的出现关系,排除偶见种,选取各森林群落中重要值较高的树种进行种间关联分析。
1.2 数据分析种群指标包括各样方的树种数、个体总数、相对频度、多度与显著度;并以重要值作为反映树种在群落中的地位和作用的指标[20]:

本研究在长白山海拔700—1700m调查了35个样方(表 1),采用双向指示种分析法划分群落类型。将35个样方中重要值<0. 05 的物种剔除,筛选出乔木种共计34种(表 2),以此构成34×35的物种重要值数据矩阵,利用采用VESPAN 软件包中Hill 1979 年设计的TWINSPAN[21]划分群落类型和树种分类。
| 样方编号Sample No. | 海拔Elevation/m | 样方编号Sample No. | 海拔Elevation/m | 样方编号Sample No. | 海拔Elevation/m |
| sp1 | 700 | sp13 | 898 | sp25 | 1056 |
| sp2 | 713 | sp14 | 898 | sp26 | 1118 |
| sp3 | 736 | sp15 | 900 | sp27 | 1120 |
| sp4 | 749 | sp16 | 901 | sp28 | 1124 |
| sp5 | 769 | sp17 | 935 | sp29 | 1126 |
| sp6 | 771 | sp18 | 937 | sp30 | 1200 |
| sp7 | 793 | sp19 | 995 | sp31 | 1300 |
| sp8 | 800 | sp20 | 1000 | sp32 | 1400 |
| sp9 | 803 | sp21 | 1012 | sp33 | 1500 |
| sp10 | 805 | sp22 | 1040 | sp34 | 1600 |
| sp11 | 821 | sp23 | 1047 | sp35 | 1700 |
| sp12 | 826 | sp24 | 1049 | — | — |
| 编号No. | 乔木种名Tree species | 编号No. | 乔木种名Tree species |
| t1 | 白桦Betula platyphylla | t18 | 糠椴Tilia mandshurica |
| t2 | 东北槭Acer mandshuricum | t19 | 裂叶榆Ulmus laciniata |
| t3 | 茶条槭Acer ginnala | t20 | 黄花落叶松Larix olgensis |
| t4 | 长白鱼鳞云杉Picea jezoensis var. komarovii | t21 | 蒙古栎Quercus mongolica |
| t5 | 稠李Prunus padus | t22 | 三花槭Acer triflorum |
| t6 | 臭冷杉Abies nephrolepis | t23 | 千金榆Carpinus cordata |
| t7 | 春榆Ulmus japonica | t24 | 青楷槭Acer tegmentosum |
| t8 | 大青杨Populus ussuriensis | t25 | 色木槭Acer mono |
| t9 | 枫桦Betula costata | t26 | 沙松冷杉Abies holophylla |
| t10 | 黑桦Betula dahurica | t27 | 櫰槐Maackia amurensis |
| t11 | 红皮云杉Picea koraiensis | t28 | 山荆子Malus baccata |
| t12 | 红松Pinus koraiensis | t29 | 山梨Pyrus ussuriensis |
| t13 | 胡桃楸Jugland mandshurica | t30 | 山杨Populus davidiana |
| t14 | 花楷槭Acer ukurunduense | t31 | 水曲柳Fraxinus mandshurica |
| t15 | 花楸Sorbus pohuashanensis | t32 | 小楷槭Acer komarovii |
| t16 | 黄檗Phellodendron amurense | t33 | 髭脉槭Acer barbinerve |
| t17 | 紫花槭Acer pseudo-sieboldianum | t34 | 紫椴Tilia amurensis |
种间总体关联性反映的是在某地出现的多物种种间是否存在显著关联性,其测定采用[22]提出的方差比率法(VR),并采用统计量W 检验VR 值偏离1 的显著程度[23]。先作零假设,即研究树种间无显著关联,选用下列公式计算检验统计量[1]:
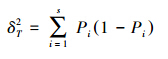
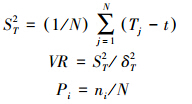
为了求出种间的联结系数和测定其联结的显著性,用2×2联列表统计各种对的具体值,即a、b、c、d 的取值。联结系数AC用来说明种间联结程度的相关系数。
若ad≥bc,则:
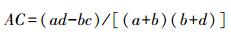
ad<bc,且d≥a,则:
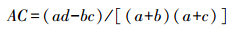
ad<bc,且d<a,则:
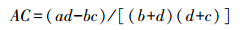
为了克服点联结系数AC受到d值影响大而造成偏差,本文选用测定两树种间关联度较好的Ochiai指数计算种间联结度:
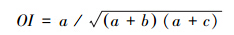
对长白山北坡调查的35个样方用TWINSPAN进行等级分类(图 1),结合调查区域的实际生态意义,采用第三级的划分结果,将海拔700—1700m的森林群落划分为3个群落类型,群落的命名主要依据优势种的类型以及采伐历史作为群落划分结果的命称。

|
| 图 1 长白山北坡35块样方的Twinspan分类树状图 Fig. 1 Dendrogram of the TWINSPAN classification of 35 quadrats in the north slope of Changbai Mountain |
(1)群落Ⅰ 天然次生林群落
该群落包含1,5—6,8—15,17—18,21—23,26—27共计18个样方,除了样方9、10外,其余16个样方均分布在海拔700—1118m。主要优势树种有白桦(Betula platyphylla)、红松、山杨(Populus davidiana)、黄花落叶松(Larix olgensis)、蒙古栎(Quercus mongolica)和青楷槭(Acer tegmentosum)等,大多数为20世纪30、40年代日本盘踞东北时皆伐后形成的白桦天然次生林。
(2)群落Ⅱ 原始阔叶红松林群落
该群落包含2—4,7,16,19—20,24—25,28—29共计11个样方,分布在海拔713—1124m的长白山原始阔叶红松林内,主要优势树种有红松、紫椴、水曲柳、蒙古栎和槭树类树种。20世纪以来群落未受到采伐干扰,保存较为完整。
(3)群落Ⅲ 云冷杉林群落
该群落包含30—35 共计6个样方,分布在海拔1200—1700m。主要优势种为长白鱼鳞云杉、臭冷杉、黄花落叶松等针叶树种。
2.1.2 树种分类长白山北坡34个树种的TWINSPAN分类结果见表 3。
| 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | 3 | 3 | 3 | 3 | 3 | 3 | |||||||||||
| 1 | 5 | 6 | 9 | 0 | 1 | 2 | 3 | 4 | 7 | 8 | 5 | 8 | 1 | 2 | 3 | 6 | 7 | 4 | 7 | 6 | 9 | 0 | 4 | 5 | 8 | 9 | 2 | 3 | 0 | 1 | 2 | 3 | 4 | 5 | ||
| 左边数字为树种编号(表 2) ,最上方为样方编号,最下方是样方分组结果,最右边是树种分类结果,中间数字采用“伪种”方法将各指标数值分级,“-”表示植物在该样方不存在,“1”表示一次分类结果,“0”表示另一组 | ||||||||||||||||||||||||||||||||||||
| T3 | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | - | - | - | - | - | - | - | 000 |
| T5 | - | 1 | - | - | - | - | - | 2 | 1 | - | - | - | 2 | 4 | - | - | - | - | - | - | 1 | - | - | - | - | - | 1 | 3 | 1 | - | - | - | - | - | - | 000 |
| T9 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | 2 | 2 | - | - | 4 | 2 | - | - | - | - | - | - | 000 |
| T11 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | 000 |
| T13 | 4 | 4 | 3 | 3 | 5 | 4 | 5 | 4 | 2 | 3 | 3 | 3 | 4 | 3 | 2 | 4 | 4 | 4 | 4 | 4 | 2 | 5 | 4 | 4 | 3 | 4 | 5 | 3 | 3 | 2 | 2 | - | - | - | - | 000 |
| T14 | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | 1 | - | - | - | - | - | - | 000 |
| T20 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | 3 | - | 2 | - | - | 3 | 2 | - | - | - | - | - | - | 000 |
| T24 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 3 | 1 | - | - | - | - | - | - | 000 |
| T27 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | - | 1 | - | - | - | - | - | - | - | - | 000 |
| T32 | 4 | 2 | - | - | 3 | 1 | 2 | 2 | 4 | 2 | 2 | - | 2 | - | 1 | - | - | - | - | 4 | 3 | - | 3 | - | - | 3 | - | 4 | 2 | - | - | - | - | - | - | 000 |
| T33 | - | 1 | - | - | - | - | - | - | - | 1 | - | - | 1 | - | - | 1 | - | - | 1 | - | - | 4 | - | 2 | 2 | - | 1 | - | - | - | - | - | - | - | - | 000 |
| T34 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 2 | 3 | 5 | 4 | - | 4 | 4 | 4 | 5 | - | 4 | - | - | - | - | - | - | 000 |
| T2 | 1 | 1 | 1 | - | - | - | - | 3 | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T12 | - | - | - | - | - | - | 1 | - | - | 2 | - | - | 3 | 3 | 1 | 1 | 4 | 4 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T17 | 1 | 1 | - | - | 1 | 1 | - | 2 | - | 1 | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 | 1 | - | - | 2 | - | - | - | - | - | - | 001 |
| T18 | 3 | 3 | 4 | 5 | 5 | 1 | 2 | 3 | 3 | 1 | 5 | 2 | 1 | - | - | - | - | - | - | - | - | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T19 | 3 | 2 | 3 | - | - | - | - | - | 1 | - | - | - | 1 | - | 2 | 3 | - | 1 | - | - | 1 | - | - | - | - | 2 | - | 1 | - | - | - | - | - | - | - | 001 |
| T22 | 2 | 3 | 4 | 3 | 3 | 3 | 3 | 2 | 3 | 3 | 3 | 4 | 2 | - | - | - | - | - | 4 | 2 | 3 | - | 1 | - | - | 4 | - | - | 1 | - | - | - | - | - | - | 001 |
| T23 | 1 | 2 | 2 | - | - | 1 | 2 | - | 2 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | 3 | - | - | - | - | - | - | 001 |
| T26 | 5 | 4 | 4 | 4 | 4 | 3 | 3 | 4 | 5 | 4 | 3 | 3 | 3 | 2 | 3 | 2 | - | - | 1 | 3 | 3 | 1 | 4 | 1 | 3 | 4 | 1 | 3 | 4 | - | - | - | - | - | - | 001 |
| T28 | - | 1 | 2 | - | 2 | 1 | 2 | 1 | 2 | 4 | 2 | 1 | 3 | 1 | - | - | - | - | 3 | 3 | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T29 | - | 1 | 1 | 1 | - | 2 | 2 | 2 | - | - | - | - | 1 | - | - | 1 | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T30 | - | - | - | - | - | 1 | 1 | - | - | 1 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 001 |
| T31 | - | 4 | 4 | - | - | 3 | 4 | - | - | 2 | 2 | - | 2 | 2 | 3 | 2 | 2 | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 001 |
| T1 | - | 4 | 4 | - | - | 5 | 4 | 4 | 4 | 5 | 3 | - | 5 | 4 | 5 | 4 | 5 | 5 | 2 | - | - | - | 1 | - | 1 | - | 1 | - | - | 3 | 4 | - | - | - | - | 01 |
| T7 | 2 | 3 | 2 | - | 2 | 2 | 2 | 2 | 2 | 1 | - | - | - | 1 | 1 | 1 | - | - | - | 2 | 4 | - | - | 1 | - | 1 | - | - | 2 | 1 | - | 2 | - | - | - | 01 |
| T8 | 3 | - | - | 3 | - | 2 | - | - | 1 | 1 | - | - | 1 | - | - | 3 | - | 1 | - | - | - | - | - | - | 4 | - | - | - | - | - | 2 | - | - | - | - | 01 |
| T25 | - | 2 | 1 | - | 2 | - | 1 | 4 | 2 | 1 | 1 | 1 | 2 | 1 | 3 | 3 | 2 | 3 | 1 | 1 | 3 | 3 | 2 | 3 | 4 | 2 | 2 | 2 | - | 3 | 2 | - | - | - | - | 01 |
| T16 | - | - | - | - | - | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 1 | - | - | - | - | - | - | - | - | 1 | - | - | 2 | 1 | - | - | 2 | 10 |
| T21 | - | - | - | - | - | - | - | - | - | 2 | 2 | 4 | 4 | - | 2 | 4 | 2 | 3 | - | - | - | - | - | - | - | - | - | - | - | 5 | 3 | 4 | 4 | 2 | 4 | 10 |
| T4 | - | - | - | - | - | 1 | 1 | - | - | - | 3 | - | - | 2 | 4 | 3 | - | - | 3 | - | 2 | 3 | 4 | 4 | 4 | - | 2 | - | - | 5 | 5 | 5 | 5 | 5 | 5 | 11 |
| T6 | - | - | - | - | - | - | - | - | - | - | 4 | 3 | 2 | 2 | 4 | 4 | 5 | 5 | 3 | - | 3 | 4 | 2 | 3 | - | - | 5 | 4 | - | 5 | 5 | 5 | 5 | 5 | 4 | 11 |
| T10 | - | - | - | - | - | - | - | - | - | - | - | - | - | - | 2 | - | - | 1 | - | - | - | 1 | - | - | 3 | - | - | - | 1 | 1 | 1 | 2 | 1 | - | - | 11 |
| T15 | - | - | - | - | - | - | - | - | - | - | 1 | 4 | - | - | 3 | 2 | - | 1 | - | 3 | - | 1 | 4 | 2 | 2 | - | - | 1 | - | 3 | 3 | 4 | 5 | 2 | 1 | 11 |
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | ||
| 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | ||||||||
本研究采用5级分类水平将长白山北坡主要乔木种分为5个植被类型,以生活型和生境为依据,根据外业调查数据,将34个乔木种划分为3个类群。
(1)类群Ⅰ
包括000和001内的所有树种,主要为阴性或中性偏阴性的树种,如稠李(Prunus padus)、黑桦(Betula dahurica)、红松、胡桃楸、裂叶榆(Ulmus laciniata)、千金榆(Carpinus cordata)、沙松冷杉(Abies holophylla)、水曲柳、紫椴和槭树类树种。
(2)类群Ⅱ
包括01内的所有树种,如白桦、春榆、大青杨、青楷槭等,该组树种主要为阳性或中性偏阳的软阔叶树种。
(3)类群Ⅲ
包括10和11内的所有树种,如花楸、落叶松、长白鱼鳞云杉、臭冷杉、枫桦、花楷槭等,主要为高海拔地区的针叶树种。
根据《中国植被》,植被型是一定特殊生境的产物[24],从上述样方和树种的分类结果可知,原始阔叶红松林包括水曲柳、胡桃楸、紫椴、黄檗等硬阔叶树种,主要以红松等阴性或中性偏阴性的树种为主;而天然次生林以白桦、大青杨等阳性或中性偏阳的软阔叶树种为主。虽然原始阔叶红松林与天然次生林的海拔相似,但是受历史人为采伐因素的影响,群落内乔木种组成差异较大。云冷杉林以分布在高海拔地区的落叶松、长白鱼鳞云杉、臭冷杉等针叶树种为主。以上分类结果说明TWINSPAN 数量分类结果与实际植被空间格局分布较为一致,是一种行之有效的群落分类方法。
2.2 种间联结性 2.2.1 原始阔叶红松林重要值是综合反映群落中不同树种的相对重要性,为了客观地选出能概括各森林群落特征的主要树种,选取最小值3 作为阈值,将阔叶红松林内重要值大于3的树种(表 4)进行种间关联分析(图 2)。红松的重要值是蒙古栎的4.24倍,原始阔叶红松林内树种组成复杂、种类丰富,各树种所占资源差异不大,建群种优势度差异不显著。
| 排序Rank | 树种Species | 重要值Ⅵ | 排序Rank | 树种Species | 重要值Ⅵ |
| 1 | 红松 | 14.02 | 6 | 色木槭 | 7.11 |
| 2 | 髭脉槭 | 11.55 | 7 | 长白鱼鳞云杉 | 4.94 |
| 3 | 紫花槭 | 9.52 | 8 | 青楷槭 | 3.92 |
| 4 | 紫椴 | 8.63 | 9 | 水曲柳 | 3.82 |
| 5 | 臭冷杉 | 7.39 | 10 | 蒙古栎 | 3.31 |

|
| 图 2 阔叶红松林内主要树种种间联结OI指数半矩阵图 Fig. 2 OI index half matrix figure of the main tree species in broad-leaved koran pine forest (S1: 二渡河(N27°27′20.7″,E110°53′40.9″),S2: 河流入库口(N27°26′58.8″,E110°53′38.2″),S3: 库尾(N27°26′22.8″,E110°53′44.5″),S4: 水库中心(N27°25′04.9″,E110°53′34.9″),S5: 库区内(N27°24′06.0″,E110°53′23.3″),S6: 主库区渡口(N27°23′21.0″,E110°54′19.5″),S7: 水库大坝(N27°22′29.2″,E110°55′21.5″)) |
根据10 个优势种群的存在与不存在矩阵,计算VR 值( VR = ST2/ δ2T = 0.6033< 1),说明10个种群间表现出负联结。对以上测定结果,采用统计量W 检测VR偏离1的显著性, W=N×(VR) =6.64;查表得相应的 χ2 值,χ0.052(11)=19.68 ,χ0.952(11)=4.57 ,因此 W 落入 χ0.952(11)与χ20.05(11) 之间,说明阔叶红松林内主要树种总体上表现出不显著的负关联。反映出阔叶红松林群落主要乔木种对环境的适应性相同或相似,但又存在资源利用上的竞争,总体间不显著的负关联,则是利用种间一定程度的分离,以降低竞争强度。
联结系数(AC)度量种间联结性的强度,可作为对种间联结性的一种说明。阔叶红松林内AC值范围在<-0.5、[-0.5,-0.1]、0、[0.1,0.5]、>0.5、不存在的种对分别占27%、6%、18%、4%、11%、33%,正负关联比为0.45,说明51%的种对间呈独立分布。阔叶红松林内种间联结度OI指数半矩阵图(图 4),表明89%的种对(40对)的OI值在[0.6,0.1],反映出原始阔叶红松林内主要树种89%的种对间关联程度紧密。
2.2.2 天然次生林天然次生林内选取重要值大于3的树种(表 5)进行种间关联分析(图 3)。白桦的重要值是青楷槭的4.84倍,天然次生林内树种组成简单,建群种优势度差异显著。
| 排序Rank | 树种Tree species | 重要值VI | 排序Rank | 树种Tree species | 重要值VI |
| 1 | 白桦 | 15.55 | 6 | 臭冷杉 | 5.92 |
| 2 | 红松 | 11.76 | 7 | 蒙古栎 | 5.42 |
| 3 | 色木槭 | 10.72 | 8 | 山杨 | 3.98 |
| 4 | 紫花槭 | 7.8 | 9 | 黄花落叶松 | 3.66 |
| 5 | 紫椴 | 7.69 | 10 | 青楷槭 | 3.21 |
根据12个优势种群的存在与不存在矩阵,计算VR 值( VR = S2T / δ2T =1.35>1),说明种群间表现出负联结。对以上测定结果,采用统计量W 检测VR偏离1的显著性, W=N×(VR) =24.3;查表得相应的 χ2 值, χ20.05(18)=28.87 , χ0.952(18)=9.39 ,因此 W 落入 χ0.952(18) 与 χ20.05(18) 之间,说明天然次生林主要树种总体上表现出不显著的正关联,反映出各物种对群落环境有较一致的要求。
天然次生林内联结系数(AC)的范围在< - 0.5、[-0.5,-0.1]、[0.1,0.5]、>0.5、不存在的种对分别占18%、17%、18%、14%、33%,正负关联比为0.91,33%的种对间呈独立分布。种间联结度OI指数半矩阵图(图 3),表明76%的种对(50对)的OI值在[0.6,1],说明主要乔木树种间76%的种对关联程度紧密。

|
| 图 3 天然次生林内主要树种种间联结OI值半矩阵图 Fig. 3 OI index half matrix figure of the main tree species in natural secondary forest |
云冷杉林内重要值大于等于3的树种有5种(表 6),高海拔地区林下树种组成简单,长白鱼鳞云杉的重要值是白桦的12.9倍,群落内建群种优势度差异很大,优势树种极为明显。
| 排序Rank | 树种Tree species | 重要值VI |
| 1 | 长白鱼鳞云杉 | 38.38 |
| 2 | 臭冷杉 | 23.80 |
| 3 | 黄花落叶松 | 12.62 |
| 4 | 花楷槭 | 8.92 |
| 5 | 白桦 | 2.98 |
根据5个优势种群的存在与不存在矩阵,计算VR 值(VR = S2T / δ2T = 2> 1),说明11个种群间表现出正联结。对以上测定结果,采用统计量W 检测VR偏离1的显著性, W=N×(VR) =12;查表得相应的 χ2 值, χ20.05(6)=12.59 , χ0.952(6)=1.64 ,因此 W 落入 χ0.952(6) 与 χ20.05(6) 之间,说明云冷杉林内主要树种总体上表现出不显著的正关联。
为了客观的分析树种间关系,将群落内所有树种进行种间关联性分析(图 4)。云冷杉林内AC值范围在<-0.5、0、[0.1,0.5]、>0.5、不存在的种对分别占2%、31%、13%、16%、38%,正负关联比为14.5,69%的种对间呈独立分布。种间联结度OI指数半矩阵图(图 4),表明26种对(占47%)的OI值在[0.6,1],说明11个树种间47%的种对关联程度紧密。

|
| 图 4 云冷杉林内主要树种种间联结OI值半矩阵图 Fig. 4 OI index half matrix figure of the main tree species in spruce-fir mixed forest |
本文采用双向指示种分析法对调查样地和树种进行分类,35个样地被分为3个群落,即海拔700—1120m为原始阔叶红松林和天然次生林,因为20世纪30、40年代日本盘踞东北时部分阔叶红松林被皆伐后形成的杨桦、白桦天然次生林;1120—1700m为云冷杉林,划分结果与前人的结论大致相同[10, 25, 26, 27]。
闫琰等[28]的研究表明长白山温带次生杨桦林和次生针阔混交林中,小尺度(10m×10m)中性过程和生态位过程均能解释群落物种多度分布,但生态位过程的解释能力相对较大,而在中等尺度(30m×30m)上中性过程开始逐渐取代生态位过程程成为维持物种多度分布的主要机制。本研究的调查采用中等尺度(30m×30m),原始阔叶红林内主要树种中第一、二类的树种占74%,第三、四类树种占20%;天然次生林内主要树种中第一、二类的树种占50%、第三、四类的树种占15%。历史采伐干扰改变了林分中的资源和空间状况[29],原始林与天然次生林在资源上优势树种宽度值不同,物种多度分布也不同。云冷杉林内主要树种中第三、五类的树种占72%,是由于臭冷杉、长白鱼鳞云杉具有最大的生态位宽度[30],是该区森林群落的主要建群种。生态位宽度较大物种对林内资源环境的适应力较强,占有的空间范围较广,在群落内的重要值也较大。应用生态位理论可以较好的解释中等尺度研究的树种分类结果。
3.2 种间关联性分析从群落中乔木种的重要值看,随着海拔高度的升高,建群种数量显著减少,云冷杉林建群种数量是原始林和次生林的一半。原始林和次生林建群种数量虽然相同,但受历史采伐的影响,次生林内白桦的重要值明显高于红松,建群种之间的优势度差异较大。从群落内种间总体关联性结果看,在较大的尺度下(样方单位面积30 m×30 m),原始阔叶红松林的建群种总体呈不显著负关联,天然次生林建群种总体呈不显著正关联。一般认为,物种总体联结随着演替的发展变化反映出次生演替各阶段群落的稳定性,特别是演替发展至后期总体上往往表现出显著正相关[31]。本研究中原始阔叶红松林未受采伐等人为影响,是处于演替后期的顶级群落;天然次生林伐后恢复近半个世纪,是处于演替中期的群落,群落内建群种总体关联性的研究结果不支持上述观点。林木间竞争是森林生态系统中的普遍现象[32],建群种是对群落结构和群落环境的形成有明显控制作用的物种,原始阔叶红松林经过长期的演替,建群种间的竞争使得该生态系统达到平衡状态。
从种间联结性结果看,3个群落内种对之间多呈正联结关系,树种间正联结是生态学特性及对环境要求的差异和互补性,导致各种之间相互兼容,互相促进生长[32],常常相伴出现。原始阔叶红松林内,黄檗与色木槭、水曲柳都为主要伴生树种,在群落中占有重要的优势度,表现出显著的正联结关系。天然次生林中,臭冷杉与落叶松都喜湿生生境,为主要伴生种。云冷杉林中长白鱼鳞云杉与臭冷杉、红松的生态习性也较为一致,都为阴性、喜冷湿环境及酸性土壤的针叶树种。杜志等[13]研究发现从云冷杉针阔混交林到云冷杉针叶混交林和近原始云冷杉林,云杉、冷杉和红松两两间的空间关联性,表现为负相关的现象增多。本研究中的云冷杉林分布在海拔1200—1700m范围内,研究范围较广,云杉与冷杉相伴出现的几率较大;随着海拔升高,红松的数量减少,因而云杉和冷杉分别于红松相伴出现的几率降低。3个群落中,绝大多数种对的联结强度存在明显的差异,因为树种对微环境适应性不同,导致它们在不同群落中存在几率不同[3, 18, 33]。只有臭冷杉与青楷槭在阔叶红松林与天然次生林中,白桦与青楷槭在云冷杉林和天然次生林中联结性均较高。了解不同群落不同演替阶段的种间联结性和联结强度,可以为森林演替不同阶段的经营管理提供理论依据。
| [1] | 黄云鹏. 武夷山米槠林主要树种种间关联性. 山地学报, 2008, 26(6): 692-698. |
| [2] | 张先平, 王孟本, 佘波, 肖扬. 庞泉沟国家自然保护区森林群落的数量分类和排序. 生态学报, 2006, 26(3): 754-761. |
| [3] | 高永恒, 曾晓阳, 周国英, 王根绪. 长江源区高寒湿地植物群落主要种群种间关系分析. 湿地科学, 2011, 9(1): 1-7. |
| [4] | 王慧敏, 庞春花, 张峰, 赵彩莉. 汾河流域中下游湿地植被优势种种间关联性分析. 生态学杂志, 2012, 31(10): 2507-2512. |
| [5] | 王伯荪, 余世孝, 彭少麟. 植物群落学实验手册. 广州: 广东高等教育出版社, 1996: 119-128. |
| [6] | 黄世能, 李章德, 骆土寿, 王伯荪. 海南岛尖峰岭次生热带山地雨林树种间的联结动态. 植物生态学报, 2000, 24(5): 569-574. |
| [7] | 龚直文, 顾丽, 亢新刚, 杨华, 赵浩彦. 长白山森林次生演替过程中林木空间格局研究. 北京林业大学学报, 2010, 32(2): 92-99. |
| [8] | Cox G W. General Ecology Laboratory Manual, Jiang Y X translate. Beijing: Science Press, 1979: 32-35. |
| [9] | 赵淑清, 方精云, 宗占江, 朱彪, 沈海花. 长白山北坡植物群落组成、结构及物种多样性的垂直分布. 生物多样性, 2004, 12(1): 164-173. |
| [10] | 曹伟, 李媛媛. 中国长白植物区系的垂直分布格局. 北京林业大学学报, 2008, 30(4): 53-58. |
| [11] | Hao Z Q, Zhang J, Song B, Ye J, Li B H. Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest. Forest Ecology and Management, 2007, 252(1/3): 1-11. |
| [12] | 姜萍, 赵光, 叶吉, 崔国发, 邓红兵. 长白山北坡森林群落结构组成及其海拔变化. 生态学杂志, 2003, 22(6): 28-32. |
| [13] | 杜志, 亢新刚, 包昱君, 杨鑫霞. 长白山云冷杉林不同演替阶段的树种空间分布格局及其关联性. 北京林业大学学报, 2012, 34(2): 14-19. |
| [14] | Dai L M, Qi L, Wang Q W, Su D K, Yu D P, Wang Y, Ye Y J, Jiang S W, Zhao W. Changes in forest structure and composition on Changbai Mountain in Northeast China. Annals of Forest Science, 2011, 68(5): 889-897. |
| [15] | Zhou S X. Forestry in China: Historical Transitions and Industry Developments. Singapore: Thomson Learning, 2006. |
| [16] | Ye Y, Fang X Q. Land use change in Northeast China in the twentieth century: a note on sources, methods and patterns. Journal of Historical Geography, 2009, 35(2): 311-329. |
| [17] | Yu D P, Zhou L, Zhou W M, Ding H, Wang Q W, Wang Y, Wu X Q, Dai L M. Forest management in northeast china: history, problems, and challenges. Environmental Management, 2011, 48(6): 1122-1135. |
| [18] | 孙中伟, 赵士洞. 长白山北坡椴树阔叶红松林群落木本植物种间联结性与相关性研究. 应用生态学报, 1997, 7(1): 1-5. |
| [19] | 郝占庆, 吴钢, 邓红兵, 李静, 曹伟. 长白山北坡暗针叶林群落特征. 生态学报, 2000, 20(6): 916-921. |
| [20] | 张金屯. 数量生态学. 北京: 科学出版社, 2004. |
| [21] | Hill M O, Šmilauer P. TWINSPAN for Windows version 2. 3 Centre for Ecology and Hydrology and University of South Bohemia, Huntingdon and České Budějovice. 2005. |
| [22] | Schluter D. A variance test for detecting species associations, with some example applications. Ecology, 1984, 65(3): 998-1005. |
| [23] | 简敏菲, 刘琪, 朱笃, 游海. 九连山常绿阔叶林乔木优势种群的种间关联性分析. 植物生态学, 2009, 33(4): 672-680. |
| [24] | 吴征镒. 中国植被. 北京: 科学出版社, 1980: 143-156. |
| [25] | 王战, 徐振邦, 李昕, 彭永山, 钱家驹, 刘子德, 杨野, 魏成录, 李延增. 长白山北坡主要森林类型及其群落结构特点. 森林生态系统研究, 1980, 1: 25-42. |
| [26] | 傅沛云, 李冀云, 曹伟, 于兴华, 朱彩霞. 长白山种子植物区系研究. 植物研究, 15(4): 491-500. |
| [27] | Yu D, Liu J, Lewis B J, Zhou L, Zhou W M, Fang X M, Wei Y W, Jiang S W, Dai L M. Spatial variation and temporal instability in the climate-growth relationship of Korean pine in the Changbai Mountain region of Northeast China. Forest Ecology and Management, 2013, 300: 96-105. |
| [28] | 闫琰, 张春雨, 赵秀海. 长白山不同演替阶段针阔混交林群落物种多度分布格局. 植物生态学报, 2012, 36(9): 923-934. |
| [29] | 王惠, 李倩, 韩雪梅, 邵国凡, 代力民. 采伐干扰下长白山阔叶红松林优势树种生态位变化动态. 东北林业大学学报, 2011, 38(10): 18-20, 53-53. |
| [30] | 刘巍, 曹伟. 长白山云冷杉群落主要种群生态位特征. 生态学杂志, 2011, 30(8): 1766-1774. |
| [31] | 张志勇, 陶德定, 李德铢. 五针白皮松在群落演替过程中的种间联结性分析. 生物多样性, 2003, 11(2): 125-131. |
| [32] | 刘金福, 洪伟, 樊后保, 林荣福. 天然格氏栲林乔木层种群种间关联性研究. 林业科学, 2001, 37(4): 117-123. |
| [33] | Hurlburt S H. A coefficient of interspecific assciation. Ecology, 1969, 50(1): 1-9. |