 2014, Vol. 34
2014, Vol. 34文章信息
- 张熙灵, 王立新, 刘华民, 清华, 刘东伟, 王炜, 梁存柱, FriedrichRecknagel
- ZHANG Xiling, WANG Lixin, LIU Huamin, QING Hua, LIU Dongwei, WANG Wei, LIANG Cunzhu, Friedrich Recknagel
- 芦苇、香蒲和藨草3种挺水植物的养分吸收动力学
- Kinetics of nutrient uptake by three emergent plants, Phragmites australis, Typha orientalis and Scirpus triqueter
- 生态学报, 2014, 34(9): 2238-2245
- Acta Ecologica Sinica, 2014, 34(9): 2238-2245
- http://dx.doi.org/10.5846/stxb201303210470
-
文章历史
- 收稿日期:2013-3-21
- 修订日期:2013-10-29






2. 中美生态、能源及可持续性科学内蒙古研究中心, 呼和浩特 010021;
3. 阿德莱德大学地球与环境科学系 澳大利亚 阿德莱德 5005;
4. 内蒙古大学生命科学学院, 呼和浩特 010021
2. Sino-US Center for Conservation, Energy and Sustainability Science in Inner Mongolia, Hohhot 010021, China;
3. School of Earth and Environmental Sciences, The University of Adelaide, Adelaide 5005, Australia;
4. College of Life Sciences, Inner Mongolia University, Hohhot 010021, China
离子吸收动力学是20世纪50年代开创和发展起来的、将Miehaelis-Menten方程应用到解释植物对介质中离子吸收动态过程的一种理论[1],它是研究植物元素吸收的重要方法,为描述根系养分吸收特性、评价不同植物种类对环境养分状况的适应性、鉴定并筛选吸收高效的植物品种提供了有利的手段[2]。70年代初,Classen和Barber[3]建立了根系养分吸收动力学方程,该方程中的离子最大吸收速率(Imax)和米氏常数(Km)两个参数可定量描述植物吸收养分离子的特征,用于研究环境条件对植物吸收养分的影响。之后Nielsen和Barber[4]对吸收动力学方程进行了进一步修正,提出了临界浓度或最低平衡浓度Cmin的概念。Imax表示离子吸收所能达到的最大速率,与载体的数目和载体的运转效率有关,Imax越大,离子吸收的内在潜力越大;Km是吸收速率为最大吸收速率的一半时外界离子浓度,反映了载体活性中心与离子的亲和力,Km值小,表明载体对离子的亲和力愈大,即不需要很高的离子浓度就可以很容易地达到 Imax;Cmin是净吸收为0时外界离子的最低浓度,Cmin值小,表明植物能从有效性非常低的环境介质中吸收该养分,对低养分的耐受能力强[5]。
吸收动力学研究在阐述水稻、小麦、玉米等栽培植物对营养物质吸收特性方面的应用己十分普遍[6, 7, 8, 9, 10],也有不少研究集中于水生植物和木本植物对N、P的吸收动力学特征[11, 12, 13, 14, 15, 16, 17]。但是对于淡水生态系统中挺水植物对N、P的养分吸收动力学研究还很少[18, 19, 20]。挺水植物在生长过程中能够吸收大量的N、P,对于改善和维持水质有着重要的作用[21, 22, 23],且适宜的挺水植物的筛选是富营养化湖泊生态修复的关键环节之一。
为筛选适宜的水生植物以开展富营养化湖泊乌梁素海外源污染截留削减和内源污染净化,本文选择了乌梁素海湖滨植被带3种主要优势挺水植物芦苇(Phragmites australis)、香蒲(Typha orientalis)和藨草(Scirpus triqueter)为受试材料,考虑到挺水植物个体差异较大,故采用常规耗竭法[8]研究此3种挺水植物的N、P吸收特性,探讨其对不同程度富营养化水体的适应能力,以期在进行生态修复工程中针对不同营养状态的富营养化水体选取适宜的植物,进而为受损湖泊湿地生态系统的恢复与重建提供理论依据。
1 试验材料与方法 1.1 研究区概况乌梁素海位于内蒙古自治区乌拉特前旗境内,是我国西北部最大的内陆湖泊,湖区界于40°36′—41°03′N,108°43 ′—108°57′E,其水域南北长35—40 km,东西宽5—10 km,湖岸线长130 km;湖盆浅平,略由北向南倾斜,最大水深4 m,平均水深 0.9 m,蓄水量约2.6亿m3[24]。是河套灌排系统的重要组成部分,主要接纳农田退水、山洪和部分生活污水,是黄河中上游重要的保水、蓄水和调水基地,也是全球范围内荒漠半荒漠地区为数不多的具有生物多样性和环境保护等多功能的大型草型湖泊,对调节内蒙古西部干旱区的生态环境和气候以及维持生物多样性等方面具有重要作用[25, 26]。然而,随着工农业现代化进程的不断推进,各种营养盐的汇入加速了乌梁素海的富营养化,使其成为典型的富营养化草型湖泊,也是世界上沼泽化最快的湖泊之一[27]。
1.2 供试材料及培养供试材料为乌梁素海3种优势挺水植物——芦苇、香蒲和藨草。试验设在玻璃温室中,将植物根部用蒸馏水洗净,1/8 Hoagland 营养液(pH值 6.5)中培养28 d,营养液成分为:0.5 mmol/L Ca(NO3)2·4H2O,0.6256 mmol/L KNO3,0.1249 mmol/L NH4NO3,0.1249 mmol/L KH2PO4,0.2505 mmol/L MgSO4·7H2O,0.0025 mmol/L FeSO4·7H2O,0.0025 mmol/L C10H14N2Na2O8,0.6253 μmol/L KI,0.0125 mmol/L H3BO3,0.0165 mmol/L MnSO4·H2O,0.0067 mmol/L ZnSO4·7H2O,0.0125 μmol/L CuSO4·5H2O,0.013 μmol/L CoCl2·6H2O,0.1281 μmol/L Na2MoO4·2H2O 。每 2 d 更换 1次营养液,再用 0.1 mol/L KOH或NaOH调节营养液 pH 值至 6.5。水培 28 d 后选取生长良好的植株用于养分吸收试验。
1.3 养分吸收实验饥饿处理:从营养液中取出植物,用去离子水冲洗根部后转入 0.2 mmol/L的 CaSO4溶液中,置于温室预培养 48 h,使其达到饥饿状态。
吸收实验:采用常规耗竭法进行H2PO-4、NH+4、NO-3吸收动力学参数的测定。将经饥饿处理后的植株移入装有1 L吸收液的玻璃烧杯中进行吸收试验(用黑色卡纸将根部遮光)。视根系生物量的大小,每个烧杯中放入1—2株植物。以0.2 mmol/L的CaSO4为支持液,用去离子水配制各种吸收液,初始浓度分别为:0.075 mmol/L KH2PO4+0.2 mmol/L CaSO4、1 mmol/L (NH4)2SO4+0.2 mmol/L CaSO4、1 mmol/L KNO3+0.2 mmol/L CaSO4;每种植物在各吸收液中分别设置3个平行重复,试验在温度为(23±1)℃、光照强度为3000 lx、相对湿度为70%—80%的温室中进行。分别于处理后 1、2、3、4、6、8、10 h后取样2 mL,并测定吸收液的单盐离子浓度。每次取样后用蒸馏水补充。取样完毕后立即取出植株,剪下根部,用滤纸吸干根表面附着的水分,并在70 ℃下烘干至恒重后称重。根据吸收液浓度与吸收时间绘制离子吸收曲线图,再根据吸收曲线方程求出动力学参数,比较分析不同植物的动力学特性。H2PO-4浓度的测定用钼锑抗分光光度法,NH+4浓度的测定用纳氏试剂分光光度法,NO-3浓度的测定用紫外分光光度法测定分光光度法测定[28],取3次重复的平均值。
1.4 动力学参数的求算及数据处理吸收动力学参数根据蒋廷惠[29]等的计算方法,以X为吸收时间,Y为吸收液中离子浓度,用一元二次方程拟合离子消耗曲线方程:
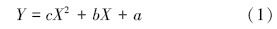
对该方程求负导数得浓度变化速率方程:
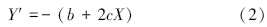
对方程(2)作如下处理:令 X→0,此时,Y′=-b,由此得到浓度最大变化速率;考虑到吸收液体积和根重,利用公式 Imax=-b×v/根干重,即可得到单位根重的最大吸收速率。Imax反映植物吸收养分的内在潜力。Y′=-b/2 代入方程(2)求出 X,将求得X代入离子消耗曲线方程(1)求出 Y,即为Km值(1/2 Imax时介质的浓度),Km值是植物耐瘠能力的评价指标之一,Km值小,说明根系吸收系统对该离子的亲和力大。令 Y′=0,求出吸收速率为 0 的时间 X,代回到方程(1)得到平衡浓度 Cmin,即吸收速率为 0 时介质中离子的最低浓度,Cmin值越小,植物越能从离子浓度很低的介质中吸收养分。
采用 Microsoft Excel 2003 绘制图表,SPSS13.0软件进行统计分析,不同处理间的显著性分析采用LSD方法。
2 结果与分析 2.1 3种挺水植物吸收 H2PO-4的动力学特性吸收液中的H2PO-4浓度随时间变化结果如图 1所示。随着吸收时间的延长,溶液中离子浓度逐渐降低。试验前植物经过“饥饿”处理,因此在吸收试验开始后的2 h内吸收离子的速度最快。据此得出3种挺水植物吸收H2PO-4的离子消耗动力学拟合方程(表 1)及其吸收H2PO-4的动力学参数(表 2)。芦苇、香蒲和藨草吸收H2PO-4的最大速率(Imax)依次为0.3895 μmol g-1干重h-1、0.3053 μmol g-1干重h-1和0.6330 μmol g-1干重h-1。藨草的 Imax显著高于芦苇和香蒲(P<0.05),说明藨草吸收H2PO-4的能力最强。芦苇、香蒲和藨草的 Km值分别为0.0637、0.0645 mmol/L和0.0702 mmol/L,表明芦苇对 H2PO-4的亲和力最强,但3种植物的Km值无显著差异(P>0.05)。芦苇的Cmin值为0.0601 mmol/L,低于另外2种植物,说明芦苇对低浓度的H2PO-4有较强的吸收能力,三者的Cmin值无显著性差异(P>0.05)。

|
| 图1 3种挺水植物吸收过程中H2PO-4浓度变化 Fig. 1 Changes of H2PO-4in the process of absorption for the three emergent plants |
芦苇、香蒲和藨草吸收液中的NH+4浓度随时间变化的情况如图 2所示。芦苇和香蒲在前1h的吸收速率最快,而藨草在试验开始后的前2 h内吸收较迅速,之后逐渐变缓。芦苇和藨草的吸收曲线比较接近。据此得出3种挺水植物对NH+4吸收的离子消耗动力学拟合方程(表 1)及其吸收NH+4的动力学参数(表 2)。芦苇、香蒲和藨草对NH+4的最大吸收速率(Imax)分别为8.995、5.584 μmol g-1干重h-1和13.819 μmol g-1干重h-1,藨草吸收NH+4的 Imax值分别是香蒲和芦苇的2.56和1.54倍,且差异显著(P<0.05),但芦苇和香蒲Imax值差异不显著(P>0.05),3种挺水植物中藨草对NH+4的吸收能力最强。芦苇、香蒲和藨草吸收NH+4的 Km值分别为 1.882 mmol/L、1.787 mmol/L和 1.874 mmol/L,香蒲的Km值显著小于芦苇和藨草(P<0.05),而芦苇和藨草对NH+4的亲和力无显著差异(P>0.05),说明香蒲对NH+4的亲和力最强。3种植物的Cmin值和Km值表现出了相同的特征,香蒲吸收低浓度NH+4的效果最好。

|
| 图2 3种挺水植物吸收过程中NH+4浓度变化 Fig. 2 Changes of NH+4 in the process of absorption for the three emergent plants |
芦苇、香蒲和藨草吸收液中NO-3浓度随时间的变化情况见图 3。随着吸收时间的延长,吸收液中NO-3浓度逐渐降低,藨草的吸收效果最好。据此得出3种挺水植物对NO-3吸收的离子消耗动力学拟合方程(表 1)及其吸收NO-3的动力学参数(表 2)。藨草吸收NO-3的最大速率(Imax)为10.146 μmol g-1干重h-1,分别是香蒲和芦苇的4倍和3倍,存在显著差异(P<0.05),但芦苇和香蒲无显著差异(P>0.05)。藨草吸收NO-3的能力最强。芦苇、香蒲和藨草的 Km值分别为0.9243 mmol/L、0.8844 mmol/L和 0.8472 mmol/L,芦苇和藨草的Km值存在显著性差异(P<0.05),二者与香蒲均无显著性差异(P>0.05),藨草的Km值最小,对 NO-3的亲和力最大;芦苇、香蒲和藨草的Cmin值依次为0.8989 mmol/L、0.8511 mmol/L和0.7992 mmol/L,排序与 Km一致,同样是芦苇和藨草之间存在显著性差异(P<0.05),二者与香蒲的差异均不显著(P>0.05),藨草的Cmin值最小,具有在 NO-3浓度很低的环境中吸收该离子的能力。

|
| 图3 3种挺水植物吸收过程中NO-3浓度变化 Fig. 3 Changes of NO-3 in the process of absorption for the three emergent plants |
|
植物种类
Plant species | H2PO-4 | NH+4 | NO-3 | |||
| 回归方程
Regression equation | R2 | 回归方程
Regression equation | R2 | 回归方程
Regression equation | R2 | |
| 注:*表示决定系数达到显著水平(P<0.05);**表示决定系数达到极显著水平(P<0.01) | ||||||
| 芦苇 Phragmites australis | 1.7628×10-5x2-
0.00093x +0.0745 | 0.9738* * | 0.00119x2-0.0249x+
1.9801 | 0.9478* * | 4.1368×10-4x2-
0.0109x+1.0004 | 0.99611* * |
| 香蒲 Typha orientalis | 1.0394×10-4x2-
0.00233x +0.0744 | 0.9783* * | 0.00233x2-0.0485x+
1.9775 | 0.9785* * | 1.1343×10-3x2-
0.0245x+0.9845 | 0.94191* * |
| 藨草Scirpus triqueter | 3.6451×10-5x2-
0.0009x +0.0746 | 0.9695* * | 0.00156x2-0.0318x+
1.9952 | 0.9757* * | 1.1216×10-3x2-
0.0280x+0.9914 | 0.9871* * |
|
植物种类
Plant species | H2PO-4 | NH+4 | NO-3 | ||||||
| Imax/
(μmolg-1 干重h-1) | Km/
(mmol/L) | Cmin/
(mmol/L) | Imax/
(μmolg-1 干重h-1) | Km/
(mmol/L) | Cmin/
(mmol/L) | Imax/
(μmolg-1 干重h-1) | Km/
(mmol/L) | Cmin/
(mmol/L) | |
| 同列标有不同小写字母表示差异显著(P<0.05),标有相同小写字母表示差异不显著(P>0.05) | |||||||||
|
芦苇
Phragmites australis | 0.3895±
0.0348b | 0.0637±
0.0056a | 0.0601± 0.0074a | 8.995±
1.9338b | 1.882±
0.0091a | 1.865±
0.0199a | 3.374±
1.183b | 0.9243±
0.0326a | 0.8989±
0.0431a |
|
香蒲
Typha orientalis | 0.3053±
0.0134b | 0.0645±
0.0008a | 0.0613± 0.0011a | 5.584±
0.1430b | 1.787±
0.0113b | 1.724±
0.0124b | 2.544±
0.066b | 0.8844±
0.0220ab | 0.8511±
0.0281ab |
|
藨草
Scirpus triqueter | 0.6330±
0.0432a | 0.0702±
0.0009a | 0.0688± 0.0012a | 13.819±
0.5309a | 1.874±
0.0026a | 1.833±
0.0058a | 10.146±
2.129a | 0.8472±
0.0310b | 0.7992±
0.0407b |
植物对营养离子的吸收作用是以低浓度下的高亲和力机制和高浓度下的低亲和力机制来共同调节的[30]。Cacco[31]等认为不同的基因型可能会形成一种“亲和力策略”(低Km值)或“速度策略”(高Imax值),提出了用Imax和Km值评价不同种类的植物对环境养分状态的适应性理论:(1)具有高Imax和低Km值的植物能适应广范的养分条件;(2)具有高Imax和高Km值的植物最能适应高水平的养分条件;(3)具有低Imax和低Km值的植物最能适应低水平的养分条件;(4)而具有低Imax和高Km值的植物在任何养分浓度条件下都是不利的。
植物因种类不同对养分的吸收状况也不同[32, 33, 34],本研究表明,芦苇、香蒲和藨草这3种挺水植物对H2PO-4、NH+4、NO-3的吸收特性存在较大差异,各自的适用范围也不同。其中,NH+4-N和NO-3-N都是植物生长的重要氮源,并且不同植物对某种氮源的偏好有着重要的生态意义和实际意义[35]。本研究中3种植物对NH+4的最大吸收速率(Imax)均大于NO-3,有研究表明,NH+4-N是香蒲、芦苇等湿地植物主要吸收的无机氮形态[36, 37, 38],与本研究的结论一致。藨草吸收NH+4和NO-3的Imax值最大,适宜用于修复高浓度NH+4和NO-3污染的水体,香蒲吸收NH+4和NO-3的Imax值最低,但香蒲吸收NH+4的Km和Cmin均最小,对NH+4的亲和力最高,能适应低浓度 NH+4环境,可以考虑用香蒲来维持修复后的水质。藨草吸收NO-3的Imax值最大,同时它的Km值和Cmin值也是最小的,说明藨草吸收NO-3的速率最快,并且在低浓度养分条件下仍能吸收NO-3,可以用于修复不同程度NO-3污染的水体。藨草吸收H2PO-4的Imax值是3种植物中最高的,芦苇的Km值和Cmin值最低。因此,当环境中H2PO-4浓度相对较高时,藨草对H2PO-4的吸收速率高于芦苇和香蒲,能够适应较高浓度P污染的水体。而当环境中H2PO-4浓度相对较低时,芦苇对H2PO-4的吸收速率要高于香蒲和藨草,适于净化低P水体。
挺水植物修复富营养化水体,主要是通过植物根系的吸收来实现的,藨草吸收H2PO-4、NH+4、NO-3的Imax均高于另外两种植物,这可能与植物根系形态有关[39]。湿地植物的净化功能与根系的发达程度相关[40]。与根茎型芦苇、香蒲相比,具须根系的藨草根系数量多且发达,表面积大,因此吸收速率较快。在选择合适的植物净化水体时,应该充分考虑其根系的生长状况。此外,本研究中植物对H2PO-4、NH+4、NO-3的Imax和Km值均低于文献最高值。这是因为在不同研究中植物所处环境不同所致,如 Brix[36, 37, 38, 39, 40]等通过研究发现植物根系环境的pH值、氮源含量等都会影响香蒲对NH+4和NO-3的吸收;同时也和植物所处的生长期不同有关。Bot[41]等发现,Imax和Km值明显受植物年龄,物种和栽培条件的影响,实验方法的不同也会对结果产生影响。有研究表明[42],冷凉条件(10 ℃)会降低山荆子幼苗对NH+4和NO-3的吸收能力,表现为最大吸收速率降低(Imax降低),亲和力下降(Km增加),其中对吸收NO-3的影响大于NH+4。由于条件限制,本试验只设置了常温处理,在后续试验中可以设置不同温度来研究高温和低温对养分吸收动力学参数的影响
本研究所选取的 3 种水生植物是乌梁素海湖滨带主要挺水植物,具有一定的生态价值和经济价值,易栽培存活,在退化湖泊湿地生态系统恢复与重建中可予以推广。在实际应用中,富营养化水体中往往不是单一的某种离子,要根据不同修复阶段的各离子浓度选取不同的水生植物来进行水体生态修复。另外,吸收动力学参数只是吸收特征的量化描述,不能反映吸收能力的深层次机理[43]。且Imax、Km和Cmin是在较短时间、相对均一的溶液中测得,与湖泊水体中挺水植物的实际吸收特性还有一定的差距,但仍然可以利用这一数量化特征作为乌梁素海退化湿地湖滨带植物生态修复中植物筛选的重要依据,开展更深入的对比和测试研究。
总之,开展多种挺水植物对不同形态N、P吸收动力学研究,对进一步明确它们对水体中N、P的去除机理,并针对不同营养特征的富营养化水体的生态修复工程中挺水植物的选取等方面均具有理论和实践意义。在没有外源性污染源输入时,水生植物能够抑制湖泊沉积物中的N、P向上覆水中释放,并能大量去除上覆水和间隙水中的N和P。由于沉积物间隙水和上覆水中的H2PO-4、NH+4、NO-3含量往往有很大差别,水生植物在生长的过程中对间隙水中的N和P也到了主要的吸收作用[44, 45, 46],所以在将植物修复应用到湖泊治理中时,不但要考虑上覆水中营养盐的浓度,也要考虑沉积物间隙水的营养负荷,使得挺水植物的修复发挥最大作用。
4 结论(1)芦苇、香蒲和藨草吸收H2PO-4、NH+4、NO-3的动力学特征存在差异。
(2)藨草对H2PO-4的最大吸收速率Imax大于芦苇和香蒲;芦苇对H2PO-4的耐瘠能力比香蒲和藨草强(Km和Cmin最小),能在环境H2PO-4浓度较低的情况下继续吸收该养分离子。
(3)藨草吸收NH+4的最大速率Imax大于芦苇和香蒲,对NH+4的吸收能力最强;香蒲的Km和Cmin值最小,对NH+4的耐瘠能力较芦苇和藨草强。
(4)藨草对NO-3的最大吸收速率Imax比芦苇和香蒲大,同时吸收NO-3的Km和Cmin值最小,藨草具有最大的Imax值和最小的Km值,能适应广泛浓度的含NO-3水体。
| [1] | Epstein E, Hagen C E. A kinetic study of the absorption of alkali cations by barley roots. Plant Physiology, 1952, 27(3): 457-474. |
| [2] | Yu Q, Kuang Y H. Kinetics of nutrient uptake by plant root system and its application. Journal of South China Agricultural University, 1997, 18(2): 105-110. |
| [3] | Claassen N, Barber S A. A method for characterizing the relation between nutrient concentration and flux into roots of intact plants. Plant Physiology, 1974, 54(4): 564-568. |
| [4] | Nielsen N E, Barber S A. Differences among genotypes of corn in the kinetics of P uptake. Agronomy Journal, 1978, 70(5): 695-698. |
| [5] | Xu L L, Chen S N, Cheng Z Q, Yang M Z. Study on absorption efficiency of ions and its kinetics of Yunnan wild rice. Plant Physiology Communications, 2006, 42(3): 406-410. |
| [6] | Santos L A, Santos W A, Sperandio M V L, Bucher C A, de Souza S R, Fernandes M S. Nitrate uptake kinetics and metabolic parameters in two rice varieties grown in high and low nitrate. Journal of Plant Nutrition, 2011, 34(7): 988-1002. |
| [7] | Sun M, Guo W S, Zhu X K, Feng C N, Guo K Q, Peng Y X. Kinetics of Nitrate and Ammonium Uptake by Different Wheat Genotypes at Seedling Stage. Journal of Triticeae Crops, 2006, 26(5): 84-87. |
| [8] | Tian X H, Li S X, Wang Q J. Preliminary study on the methods for determining absorption kinetic parameters of NO3- by using some crops. Chinese Journal of Soil Science, 2001, 32(1): 16-18. |
| [9] | Sun X M, Feng K, Wang X L. Kinetics of high affinity system for K+ and effects of NH4+. Plant Nutrition and Fertilizer Science, 2007, 13(2): 208-212. |
| [10] | Kronzucker H J, Glass A D M, Siddiqi M Y, Kirk, G J D. Comparative kinetic analysis of ammonium and nitrate acquisition by tropical lowland rice: implications for rice cultivation and yield potential. New Phytologist, 2001, 145(3): 471-476. |
| [11] | Cai S M, Liu W T, Zhang Z, Bai Y C, Qian X Q. Phosphorus uptake kinetics of different types of duckweed. Journal of Ecology and Rural Environment, 2011, 27(2): 48-52. |
| [12] | Zhang Y J, Liu C Q, Li H B, Wu Yi H, Wang Y B. Nitrogen and phosphorus removal performance and kinetics absorption by Ipomoea aquatica. Chinese Journal of Environmental Engineering, 2011, 5(5): 1057-1061. |
| [13] | Wei H X, Xu C Y, Ma L Y, Jiang L N , Li X L, Yang Z. Dynamic kinetic characteristics of different forms of nitrogen absorbed by Larix olgensis seedling. Plant Nutrition and Fertilizer Science, 2010, 16(2): 407-412. |
| [14] | Chen S Y, Xu C, Yao Y, Huang L Z, Zhang Y T. Uptake kinetics of ammonia, nitrate and phosphorus by submerged macrophytes Hydrilla verticillata and Vallisneria natans. Environmental Science and Technology, 2012, 35(8): 34-36. |
| [15] | Wang Y, Zhang J F, Chen G C. Removal efficiency and uptake kinetics of nitrogen in water body by Salix integra. Chinese Journal of Ecology, 2012, 31(9): 2305 -2311. |
| [16] | Gras A F, Koch M S, Madden C J. Phosphorus uptake kinetics of a dominant tropical seagrass Thalassia testudinum. Aquatic Botany, 2003, 76(4): 299-315. |
| [17] | Pérez-Lloréns J L, Niell F X. Short-term phosphate uptake kinetics in Zostera noltii Hornem: a comparison between excised leaves and sediment-rooted plants. Hydrobiologia, 1995, 297(1): 17-27. |
| [18] | Tang Y X, Zheng J M, Lou L P, Zhang Q C. Comparisons of NH4+、NO3-、and H2PO4- uptake kinetics in three different macrophytes in waterlogged condition. Chinese Journal of Eco-Agriculture, 2011, 19(3): 614-618. |
| [19] | Tylova-Munzarova E, Lorenzen B, Brix H, Votrubova O. The effects of NH4+ and NO3-on growth, resource allocation and nitrogen uptake kinetics of Phragmites australis and Glyceria maxima. Aquatic Botany, 2005, 81(4): 326-342. |
| [20] | Zhang Z H, Rengel Z, Meney K. Kinetics of ammonium, nitrate and phosphorus uptake by Canna indica and Schoenoplectus validus. Aquatic Botany, 2009, 91(2): 71-74. |
| [21] | Xu D F, Xu J M, Wang H S, Luo A C, Xie D C, Ying Q S. Absorbability of wetland plants on N and P from eutrophic water. Plant Nutrition and Fertilizer Science, 2005, 11(5): 597-601. |
| [22] | Sun R L, Zhang J, Wang W X. Effect on polluted water purification by eight emergent plants. Journal of Shandong University: Natural Science, 2009, 44(1): 12-16. |
| [23] | Cui L J, Li W, Zhang M Y, Zhao Y H, Wang Y F, Zhao X S, Shang X J. Different wetland plant roles of removing nitrogen and phosphorus on sewage water. Journal of Lake Sciences, 2011, 23(2): 203-208. |
| [24] | Wei W, Cui L J, Li S N, Guo J. Study on the wetland changes monitoring of Wuliangsuhai Lake based on average deviation. Forest Research, 2012, 25(6): 719-725. |
| [25] | Sun H M, He J, Gao X D, Lv C W, Fan Q Y, Xue H X. Distribution of total phosphorus in sediments of Wuliangsuhai Lake. Acta Sedimentologica Sinica, 2006, 24(4): 579-584. |
| [26] | Li X, Zhang S L, Li C Y, Li W P, Wang L X. Ecological characteristics of phytoplankton community of Wuliangsuhai Lake in Inner Mongolia. Ecology and Environmental Sciences, 2012, 21(11): 1865-1869. |
| [27] | Yu R H, Li C Y, Liu T X, Xu Y P. Change of wetland environment in Wuliangsuhai. Acta Geographica Sinica, 2004, 59(6): 948-955. |
| [28] | Chinese SEPA. Water and wastewater analytical method. 4th ed. Beijing: China Environmental Sciences Press, 2002. |
| [29] | Jiang T H, Zheng S J, Shi J Q, Hu A T, Shi R H, Xu M. Several considerations in kinetic research on nutrients uptake by plants. Plant Nutrition and Fertilizer Science, 1995, 1(2): 11-17. |
| [30] | Fageria N K. Kinetics of phosphate absorption by intact rice plants. Australian Journal of Agricultural, 1974, 25(3): 395-400. |
| [31] | Cacco G, Ferrari G, Saccomani M. Pattern of sulfate uptake during root elongation in maize: its correlation with productivity. Physiologia Plantarum, 2006, 48(3): 375-378. |
| [32] | Li Y Q, Jiang Y H, Li Z H, Zhao L Y. Nitrogen removal from eutrophic water by terrestrial economic plants floating-bed system. Environmental Science and Technology, 2010, 33(8): 103-107. |
| [33] | Yin L C , ZhangY Z, Zhou W J, Mao Z G. NO3-, H2PO4- and K+ uptake by three weed species, Erysimum cheiranthoides, Chorispora tenella, and Veronica agrestis in winter wheat cropping land. Chinese Journal of Soil Science, 2007, 38(1): 68-71. |
| [34] | Dang C X, Shen Q R, Wang G. Tomato growth and organic acid changes in response to partial replacement of NO3-N by NH4+-N. Pedosphere, 2004, 14(2): 159-164. |
| [35] | Forde B G, Clarkson D T. Nitrate and ammonium nutrition of plants: physiological and molecular perspectives. Advances in Botanical Research, 1999, 30: 1-90. |
| [36] | Brix H, Dyhr-Jensen K, Lorenzen B. Root-zone acidity and nitrogen source affects Typha latifolia L. growth and uptake kinetics of ammonium and nitrate. Journal of Experimental Botany, 2002, 53(379): 2441-2450. |
| [37] | Fang Y Y, Babourina O, Rengel Z, Yang X E, Pu P M. Ammonium and nitrate uptake by the floating plant Landoltia punctata. Annals of Botany, 2007, 99(2): 365-370. |
| [38] | Jampeetong A, Brix H. Nitrogen nutrition of Salvinia natans: Effects of inorganic nitrogen form on growth, morphology, nitrate reductase activity and uptake kinetics of ammonium and nitrate. Aquatic Botany, 2009, 90(1): 67-73. |
| [39] | Chen W Y, Chen Z H, He Q F, Wang X Y, Wang C R, Chen D F, Lai Z L. Root growth of wetland plants with different root types. Acta Ecologica Sinica, 2007, 27(2): 450-457. |
| [40] | Chen S P. Advances in biological fundamental studies on artificial wetland wastewater treatment system. Journal of Lake Sciences, 1996, 8(3): 268-272. |
| [41] | Le Bot J, Adamowicz S, Robin P. Modelling plant nutrition of horticultural crops: a review. Scientia Horticulturae, 1998, 74(1): 47-82. |
| [42] | Lü D G, Wang Y, Qin S J, Ma H Y, Liu G C, Du G D, Meng Q. Effects of cool and cold conditions on nitrogen uptake kinetics in Malus baccata Borkh. seedlings. Acta Horticulturae Sinica,2010, 37(9): 1493-1498. |
| [43] | Jin C H, Lu K H, Hu Z Yong, Xie L F, Liang J J, Zhang C J. Kinetics of ammonium and nitrate uptake by Myriophyllum aquaticum and Eichhornia crassipes. Acta Hydrobiologica Sinica, 2011, 35(1): 75-79. |
| [44] | Li R Z, Li F, Zhou A J. Sediment and pore water nutrient characteristics in growing zones of Alternanthera philoxeroides in the Shiwuli river, Chaohu lake. Environmental Science, 2012, 33(9): 3014-3023. |
| [45] | Tong C H, Yang X E, Pu P M. Effects and mechanism of hydrophytes on control of release of nutrient salts in lake sediment. Journal of Agro-Environmental Science, 2003, 22(6): 673-676. |
| [46] | Wang W W, Li D J, Gao L. Vegetation influence on nutrients distribution in pore water of salt marsh sediment. Environmental Science, 2009, 30(11): 3209-3217. |
| [2] | 余勤, 邝炎华. 根系养分吸收动力学研究及应用. 华南农业大学学报, 1997, 18(2): 105-110. |
| [5] | 徐玲玲, 陈善娜, 程在全, 杨明挚. 云南野生稻离子吸收效率及其动力学特征研究. 植物生理学通讯, 2006, 42(3): 406-410. |
| [7] | 孙敏, 郭文善, 朱新开, 封超年, 郭凯泉, 彭永欣. 不同氮效率小麦品种苗期根系的NO-3、NH+4吸收动力学特征. 麦类作物学报, 2006, 26(5): 84-87. |
| [8] | 田霄鸿, 李生秀, 王清君. 几种作物NO-3吸收动力学参数测定方法初探. 土壤通报, 2001, 32(1): 16-18. |
| [9] | 孙小茗, 封克, 汪晓丽. K+高亲和转运系统吸收动力学特征及其受NH+4影响的研究. 植物营养与肥料学报, 2007, 13(2): 208-212. |
| [11] | 蔡树美,刘文桃,张震, 柏彦超, 钱晓晴. 不同品种浮萍磷素吸收动力学特征. 生态与农村环境学报, 2011, 27(2): 48-52. |
| [12] | 张亚娟, 刘存歧, 李洪波, 吴亦红, 王亚斌. 蕹菜对富营养化水体的氮磷去除及吸收动力学研究.环境工程学报,2011, 5(5): 1057-1061. |
| [13] | 魏红旭, 徐程扬, 马履一, 江俐妮, 李雪莲, 杨卓. 长白落叶松幼苗对铵态氮和硝态氮吸收的动力学特征. 植物营养与肥料学报, 2010, 16(2): 407-412. |
| [14] | 陈少毅, 许超, 姚瑶, 黄立章, 张云涛. 黑藻和苦草对氨氮、硝态氮和磷吸收动力学研究. 环境科学与技术, 2012, 35(8): 34-36. |
| [15] | 王瑛, 张建锋, 陈光才. 柳树对水体氮素的去除率及其吸收动力学. 生态学杂志, 2012, 31(9): 2305 -2311. |
| [18] | 唐艺璇, 郑洁敏, 楼莉萍, 张奇春. 3种挺水植物吸收水体NH+4、NO-3、H2PO-4的动力学特征比较. 中国生态农业学报, 2011, 19(3): 614-618. |
| [21] | 徐德福, 徐建民, 王华胜, 罗安程, 谢丹超, 应求实. 湿地植物对富营养化水体中氮、磷吸收能力研究. 植物营养与肥料学报, 2005, 11(5): 597-601. |
| [22] | 孙瑞莲, 张建, 王文兴. 8种挺水植物对污染水体的净化效果比较.山东大学学报: 理学版, 2009, 44(1): 12-16. |
| [23] | 崔丽娟, 李伟, 张曼胤, 赵玉辉, 王义飞, 赵欣胜, 商晓静. 不同湿地植物对污水中氮磷去除的贡献. 湖泊科学, 2011, 23(2): 203-208. |
| [24] | 韦玮, 崔丽娟, 李胜男, 郭嘉. 基于偏差平均值的乌梁素海湿地变化监测研究.林业科学研究, 2012,25(6): 719-725. |
| [25] | 孙惠民, 何江, 高兴东, 吕昌伟, 樊庆云, 薛红喜. 乌梁素海沉积物中全磷的分布特征. 沉积学报, 2006, 24(4): 579-584. |
| [26] | 李兴, 张树礼, 李畅游, 李卫平, 王立新.乌梁素海浮游植物群落特征分析. 生态环境学报, 2012, 21(11): 1865-1869. |
| [27] | 于瑞宏, 李畅游, 刘廷玺, 许有鹏.乌梁素海湿地环境的演变. 地理学报, 2004, 59(6): 948-955. |
| [28] | 国家环境保护总局. 水和废水监测分析方法(第四版).中国环境科学出版社, 2002. |
| [29] | 蒋廷惠, 郑绍建, 石锦芹, 胡霭堂, 史瑞和, 徐茂. 植物吸收养分动力学研究中的几个问题. 植物营养与肥料学报, 1995, 1(2): 11-17. |
| [32] | 李艳蔷, 姜应和, 李兆华, 赵丽娅. 陆生经济植物浮床去除富营养化水中氮素研究. 环境科学与技术, 2010, 33(8): 103-107. |
| [33] | 尹力初, 张杨珠, 周卫军, 毛政国. 婆婆纳、离子草与小花糖芥三种麦田杂草吸收NO-3, H2PO-4, K+的动力学研究. 土壤通报, 2007, 38(1): 68-71. |
| [40] | 成水平. 人工湿地废水处理系统的生物学基础研究进展. 湖泊科学, 1996, 8(3): 268-272. |
| [42] | 吕德国, 王英, 秦嗣军, 马怀宇, 刘国成, 杜国栋, 孟倩. 冷凉条件对山荆子幼苗根系氮素吸收动力学参数的影响. 园艺学报, 2010, 37(9): 1493-1498. |
| [43] | 金春华, 陆开宏, 胡智勇, 谢丽凤, 梁晶晶, 张春景. 粉绿狐尾藻和凤眼莲对不同形态氮吸收动力学研究. 水生生物学报, 2011, 35(1): 75-79. |
| [44] | 李如忠, 李峰, 周爱佳. 巢湖十五里河水花生生长区沉积物及间隙水中营养盐的基本特性. 环境科学, 2012, 33(9): 3014-3023. |
| [45] | 童昌华, 杨肖娥, 濮培民. 水生植物控制湖泊底泥营养盐释放的效果与机理. 农业环境科学学报, 2003, 22(6): 673-676. |
| [46] | 王伟伟, 李道季, 高磊. 盐沼植物对沉积物间隙水营养盐分布的影响.环境科学,2009, 30(11): 3209-3217. |