 2014, Vol. 34
2014, Vol. 34文章信息
- 田媛, 塔西甫拉提·特依拜, 李彦, 唐立松, 范连连
- TIAN Yuan, TASHPOLAT Tiyip, LI Yan, TANG Lisong, FAN Lianlian
- 梭梭幼苗的存活与地上地下生长的关系
- The survival and above/below ground growth of Haloxylon ammodendron seedling
- 生态学报, 2014, 34(8): 2012-2019
- Acta Ecologica Sinica, 2014, 34(8): 2012-2019
- http://dx.doi.org/10.5846/stxb201309092241
-
文章历史
- 收稿日期:2013-9-9
- 修订日期:2014-3-15






2. 中国科学院新疆生态与地理研究所, 荒漠与绿洲生态国家重点试验室, 乌鲁木齐 830011;
3. 中国科学院阜康荒漠生态系统研究站, 新疆阜康 831500;
4. 新疆大学绿洲生态教育部重点实验室, 乌鲁木齐 830046
2. State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Urumqi 830011, China;
3. Fukang Stations of Desert Ecology, Chinese Academy of Sciences, Xinjiang, Fukang 831500, China;
4. Key Laboratory of Oasis Ecology, Xinjiang University, Urumqi 830046, China
植物生长规律一方面受自身生物学特性影响,另一方面受其生境条件的制约[1]。在陆地生态系统中,水分条件是限制植物个体生长发育的关键因素之一[2],水分的变化直接影响着植物的光合作用、生存以及群落生产力[3],直接或间接影响着建群种和优势种的水分利用方式和形态调整,从而决定了植物群落的分布格局和结构组成。同时,水分变化对植物生长和生态系统生产力的影响是复杂而广泛的,因植物物种和生态系统不同而差异显著[4, 5]。
干旱区生态系统结构较为简单,抵御外界干扰的能力较弱,被认为是对全球变化响应最为剧烈的生态系统类型之一[6]。以荒漠地区为例,降雨量和潜在蒸发量反差巨大,长期的干旱使荒漠区植物在生理和形态上形成了一系列对水分匮缺的适应特性和调节能力,但荒漠区植物的生理活动仍然受到水分有效性的严格制约[7, 8, 9],且对水分变化非常敏感。近50年来,中亚干旱区的降水表现出显著增多的趋势[10]。位于天山北坡洪积冲积扇与古尔班通古特沙漠南缘之间的大面积区域,亦在近20年表现出降水增多的明显趋势[11]。与此同时,人类的开发与利用,也极大地改变了该区的各种环境条件,例如,节水灌溉的推广,使得古尔班通古特沙漠南缘绿洲—荒漠交错区的地下水位的显著下降[12]。在这样的背景下,现有的植物群落将做出何种变化,成为了亟待解决的重要生态问题。
藜科梭梭(Haloxylon ammodendron)属植物主要分布在亚非大陆温带和亚热带的干旱区,以其为建群种的群落是中亚荒漠中分布最广的植被类型。在亚非大陆连续分布区,梭梭属有11种,主要生长在流动沙丘、半固定沙丘、盐渍土及砾质戈壁上[13, 14]。在古尔班通古特沙漠,梭梭是该区盐生、旱生荒漠植物群落的建群种,具有非常重要的生态意义。
水分是荒漠植物生长最重要的限制因子,荒漠植物的根系分布和同化器官生长与环境水分有效性之间的关系决定着植物的存活,个体尺度的植物水分关系研究已成为认识荒漠植物水分适应机理、预测干旱生态系统未来的关键环节。近20年来,国内外相关研究主要集中在梭梭等建群种的植物水分关系和抗旱生理方面[15, 16, 17, 18, 19]。也有研究表明,成年梭梭对降水事件的响应与其根功能型紧密相关,成年梭梭可以通过个体形态调节来维持自身水分平衡和碳平衡[20, 21]。但由于野外实验的实际困难,梭梭生活史中最脆弱同时也是决定梭梭种群能否自然更新的关键阶段:幼苗阶段的实验研究仍然不足,梭梭幼苗存活与地上地下生长关系的相关理论尚未到验证。本文以生长在古尔班通古特沙漠南缘中的当年生梭梭幼苗为研究对象,在全年生长季中定期监测幼苗的生长、生存的各项指标,研究梭梭幼苗在自然条件下的生长动态,以期探明梭梭幼苗的存活与其生长规律,以期为认识梭梭群落的更新演替及梭梭植被的人工恢复,提供理论依据。
1 材料与方法 1.1 研究区概况古尔班通古特沙漠地处准噶尔盆地中央,地理范围为44°11′N—46°20′N,84°31′E—90°00′E,为中国第二大沙漠。实验在位于中国科学院阜康荒漠生态系统国家野外科学试验研究站(44°22′N,87°55′E,海拔448m)附近的古尔班通古特沙漠南缘开展。该区处于欧亚大陆腹地,夏季炎热干燥,冬季寒冷,年均温6.6℃,年均降水量160mm,年日照时数为3079h,6—8月日照时数超过10h,年均蒸发力超过1000mm,属于典型的温带大陆性荒漠气候,地表分布着梭梭(丘间地)、白梭梭(沙丘顶部)为优势种的荒漠植物群落。
1.2 样地设计与管理在古尔班通古特沙漠南缘,选取20m×20m平坦且无大型植株的丘间地(梭梭天然分布地),于2009年3月9日积雪融化前人工播种梭梭种子。播种方法:成行列播种,行间距与列间距均为0.5m,在每个行列交叉处积雪上下种约10粒,然后覆盖0.5—1cm厚的沙土。实验所用梭梭种子是2008年秋季采集,为了在冬季抵御鼠害的同时又不破坏种子的自然萌发过程,在经过初步筛选后,保存在-18℃的冰箱中至来年3月初播种前。
1.3 观测方法 1.3.1 土壤重量含水率从2009年3月31号到2009年9月2号,以10d为周期取样,取样深度随梭梭幼苗根系长度的增加不断加深,从开始的50cm增加到后期的70cm。每次分别在样地的东、南、西、北、中5个位置以内径5cm土钻取样,0—10cm每5cm取一个样,10—70cm每10cm取1个样,用烘干法测定土壤湿度。
1.3.2 梭梭生长动态监测4月中旬,大部分梭梭幼苗地上部分的高度达到1cm左右,开始梭梭个体形态监测,持续到9月初梭梭幼苗同化枝开始脱落为止,监测时段为4月15日至9月2日10d进行1次监测。每次选取高度和冠幅具有代表性的8株梭梭幼苗,在离植株根茎为10—15cm处挖开半圆型坑,沿坑壁将根系慢慢从上到下挖出将幼苗根系完整的取出。利用普通卷尺,测量梭梭幼苗的株高与根长。测量完成后,带回实验室,用水洗去附着在幼苗上的泥土,对地上和地下部分分别进行扫描,测定光合面积和根系表面积。因为梭梭幼苗地上部分的同化枝到7月中旬才开始出现木质化,并且只有主干少量木质化,所以用地上部分面积代表同化枝面积。未木质化的根系为白色,扫描效果不好,因此先用甲基红染色后进行扫描。扫描后的样品,放入烘箱105℃杀青2h,65℃烘干48h,用天平称取其地上及地下生物量。
1.3.3 幼苗死亡率监测在4月中旬,大部分梭梭幼苗株高约1cm时,给每株梭梭编号并记录,同样以10d为周期,按幼苗编号监测其死亡数量,直至9月初,与生长动态监测同步进行。梭梭幼苗死亡率用以下公式计算:
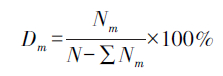
式中,Dm 是月平均死亡率,Nm是相邻两月梭梭幼苗总数的差值,即幼苗死亡数量,N是幼苗萌发时的总量。
1.3.4 气象因子气象因子的监测由自动气象站完成,测量项目包括气温、相对湿度、水气压、风速、风向、总辐射与降水量等。每30s测量1次,每30min取1个平均值,数据由CR10数据采集器(Campbell Scientific Cor,USA)记录。
大气水汽压饱和差d计算公式:
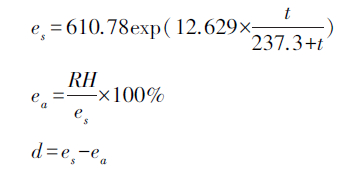
式中,es是饱和水汽压,ea是实际水汽压,RH是相对湿度[22]。
1.3.5 数据分析利用SPSS13进行数据分析,计算平均值和标准方差,单因素方差分析(ANOVA)用于处理间差异的显著性检验,平均值的标准方差用误差棒示。
2 结果与分析 2.1 梭梭幼苗生长期环境因子图 1给出了2009年生长期各月土壤平均含水率、气温、大气水汽压饱和差(d)和梭梭幼苗死亡率的变化。图 1显示了4—8月梭梭主根区(0—60cm)土壤的水分变化,其中,4、5月份的土壤水分充沛,土壤水含量在3.5—4%左右,而从6月起土壤含水率显著下降至2左右%(P=0.05),到了7月和8月,各个土层的含水率几乎都低于1.5%。同时,在生长期中,各月10:00—18:00平均气温呈阶梯式上升,7月最高,为37.1℃,8月气温略有下降。大气水汽压饱和差也逐月增高,到了7月达到峰值,也就是说随着气温的升高,空气越来越干燥,蒸发需求愈加强烈(图 1)。但是由图 1得知,梭梭幼苗死亡高峰期并非发生在最干旱的7、8月,而是发生在幼苗萌生初期。
2.2 生长期梭梭幼苗地上部分高度和地下部分深度变化动态图 2a中,梭梭幼苗株高从4月初萌发起至6月初增加迅速,尤其在5月地上高度显著增加,6月中下旬增长开始减缓并略有波动(P=0.05)。进入7月,幼苗株高有显著下降(P=0.05),平均值从6.75cm降至3.58cm,几乎下降了一半,梭梭幼苗同化枝出现了比较严重凋落现象。从梭梭幼苗地上部分的构成来看,其上半部主要是由未木质化的同化枝构成。同化枝的凋落,使幼苗株高显著降低。但在整个生长期,梭梭幼苗的根系基本保持了明显的增长趋势,虽然有一定的波动。

|
| 图 1 生长期梭梭幼苗主根区域 (0—60cm) 土壤含水率、月平均白天温度 (10:00—18:00) 、饱和差和梭梭幼苗死亡率的变化动态 Fig. 1 Dynamic changes of Haloxylon ammodendron seedling root region (0—60cm) soil water content,monthly mean daytime temperature (10:00—18:00) ,saturation deficit,seedling mortality in the whole growing period |

|
| 图 2 生长期梭梭幼苗株高和根长的变化动态 Fig. 2 Dynamic changes of Haloxylonammodendron seedlings plant heights,rooting depths in the whole growing period |
图 3分别显示了年生长期中梭梭幼苗地上和地下部分表面积变化状况。幼苗地上部分表面积在土壤水分较为充沛的4月初到6月中旬持续增加,其中5月增长最为迅速(P=0.05),在6月中旬达到最大表面积3.6cm2/株。但从6月下旬起,地上部分的表面积显著下降直至1.06cm2/株 (P=0.05),随后虽然在8月略有增加,但仍然处于较低水平。幼苗地下部分表面积在整个生长期中主要有3个显著增加期,分别为:5月中旬至5月底;6月;7月中旬至7月底(P=0.05),并在7月底达到峰值5.48cm2(图 3)。

|
| 图 3 生长期梭梭幼苗地上部分表面积和地下部分表面积的变化动态 Fig. 3 Surface area of Haloxylonammodendron seedlings aboveground parts and underground parts dynamic change in the whole growing period |
图 4为梭梭幼苗在生长期中地上和地下生物量的动态变化。在土壤水分较充沛的4至6月,幼苗的地上生物量持续增加,在6月底达到峰值0.18g/株。但在随后的7月,显著下降至0.05g/株(P=0.05),随后略有上升,基本稳定在0.12g/株。地下生物量同样是在4—6呈现快速增长,在7月显著下降后。但与地上生物量不同的是,地下生物量在8月持续增长,其峰值达到0.10g/株(图 4)。梭梭幼苗地上和地下部分的生物量数据与其表面积数据所表现的趋势基本一致,在7月时,幼苗地上和地下部分都会出现不同程度的凋落。随后可能幼苗调整了生长策略,地下部分的生物量迅速恢复并持续增长,超过了6月份的峰值,但地上部分的生物量增长相对缓慢,并且没有超过以前的峰值。

|
| 图 4 生长期梭梭幼苗地上和地下生物量的变化动态 Fig. 4 Biomass of Haloxylon ammodendron seedlings aboveground and underground parts dynamic changes in the whole growing period |
图 5展现了生长期中梭梭幼苗地上和地下生物量的月增长状况。如图所示,幼苗地上和地下部分的生物量在的5月、6月都显著增长,在7月,都显著降低。数据表明,在土壤水分充沛的前期,地上和地下部分的生物量的都快速增长,呈现相似的趋势。进入7月后,地上和地下部分的生物量开始显著下降。但从减少的幅度来看,地上部分生物量的减少幅度远大于地下部分(地上和地下部分的生物量减少分别为0.10和0.01g/株)。此后,梭梭幼苗逐渐开始适应了土壤水分匮缺的环境,虽然8月份的土壤水分条件更差,但幼苗的生长有所恢复,地上与地下部分的生物量都开始增加。从增加的幅度来看,地下部分生物量的增加大于地上部分生物量的增加。图 5的数据表明,随着土壤水分的减少,在水分胁迫的压力下,梭梭幼苗会向根系分配更多的光合产物。

|
| 图 5 梭梭幼苗地上和地下生物量月增长量 Fig. 5 Monthly biomass growth of Haloxylon ammodendron seedlings aboveground and underground parts |
从梭梭幼苗根冠比的变化(图 6)可以看出,在4月到6月幼苗根冠比虽然呈现一定的波动,但基本处于稳定的范围内。进入7月后,随着土壤水分的减少,根冠比呈现明显的增长趋势,尤其在7月上旬根冠比增长迅速,直至8月底基本稳定在0.8左右。图 6说明,随着土壤水分的减少,梭梭幼苗通过将更多的光合产物分配到根系的策略,维持了植株的水分平衡和生长。

|
| 图 6 生长期梭梭幼苗根冠比的动态变化 Fig. 6 Dynamic changes of Haloxylon ammodendron seedlings root shoot ratio in the whole growing period |
土壤水分条件的改变对植物的光合产物分配有着显著地影响,植物可以通过改变地上和地下部分比例,来维持水分供应与需求之间的平衡[23, 24]。荒漠灌木可以通过调节根系向着最优(最有利)表现型发展,从而最大程度地获取水分[25]。另外,根据Eagleson的生态最优化理论[26],植物趋向于优化同化器官密度以适应环境水分有效性。
赵兴梁[27]对沙坡头固沙植物与沙层水分关系的研究表明,当沙层湿度≥2%时,固沙植物能正常生长;当沙层湿度在1%—2%时,固沙植物生长开始衰退、萎缩。本研究中,从图 1可以看到,在生长期前期的4—6月份,融雪提供了大量水分,土壤水分充沛。这一时期也是梭梭幼苗快速生长的时期,地上和地下部分生长趋势也基本一致(图 2),同时维持了比较低的根冠比(图 6)。在土壤水分较为充沛的4—6月,梭梭幼苗的死亡率较高(图 1),比较充分的土壤水分使得梭梭幼苗不需要太多的根系,就可以维持植株的水分平衡。根据已有研究(28-29),这一时期比较高的死亡率,可能是由于较高密度导致的种内与种间的竞争。随着植物生长与土壤蒸发的消耗,进入7月,土壤水分含量开始低于1.5%,并且随着气温的升高,饱和压差显著增加,大气蒸发潜力也显著增加(图 1),梭梭幼苗地上和地下部分都出现了不同程度的凋落。这表明,随着水分的减少,虽然根系在不断向下层土壤扩展(图 2),但并不能满足同化枝蒸腾的需求。原有的地下部分(根系)供水与地上部分(同化枝)耗水之间的平衡被打破。在水分胁迫的压力下,梭梭幼苗的同化枝开始凋落以减少水分消耗。由于同化枝凋落会导致光合产物减少(图 4),这造成了地下部分生物量的减少(图 5)。但相对而言,地下部分生物量的减少要远小于地上部分生物量的减少,同化枝的凋落使梭梭幼苗可以维持地下部供水与地上部分耗水之间的平衡,进而保证了存活同化器官的活性,并且有效地降低了梭梭幼苗的死亡率(图 1),使梭梭幼苗能够渡过干旱期。在Schulze的研究中,干旱时期荒漠灌木利用脱落减少水分消耗,且地上部分脱落速率要大于地下部分[30],这与实验中梭梭幼苗在遭遇干旱时的表现一致。在生长期后期(8月),虽然土壤水分含量进一步降低(图 1),但由于地上地下不同程度的脱落,梭梭幼苗可以维持植株的水分平衡,地上和地下部分都恢复了生长,但地上部分生长缓慢,表明梭梭幼苗仍然经受着比较严重的水分胁迫。根冠比的增加 (图 6)和地下部分的迅速生长(图 3,图 4)显示:梭梭幼苗将更多的光合产物分配给根系,这与许皓[20]对成年梭梭的研究结果一致。实验表明,梭梭幼苗在土壤水分条件恶化时,通过大量减少地上部分生物量和将更多地将光合产物分配给地下部分,来建立新的水分平衡,使梭梭幼苗可以在土壤水分含量低于1.5%的条件下,仍然维持一定程度的生长。
4 结论在土壤水分不同时梭梭幼苗地上地下部分的不同生长特征是梭梭幼苗对干旱区生境长期遗传适应的结果。这种生长特征包括两个方面:一方面,在土壤水分条件恶化时,地上部分的凋落使梭梭幼苗能够大幅度降低耗水,虽然由于光合能力的降低,地下部分也有一定程度的减少,但减小程度远低于地上部分,从而可使其迅速建立植株耗水与供水的新平衡。另一方面,在新的水分平衡建立后,梭梭幼苗开始恢复生长,但将更多的光合产物分配给地下部分,以确保植株的水分供应。这种随着土壤水分变化的地上地下生长关系不但能使梭梭幼苗在水分匮乏的干旱期存活,而且还可以使梭梭幼苗维持较低但相对稳定的生长。
| [1] | Sun D X, Li S X, Qian S T, Liu Z H, Han G, Wen L F. An analysis on the community biomass dynamics and economic characters of artificial Caragana intermedia community in semi-desert area. Journal of Northwest Forestry University, 2005, 20(2): 24-27. |
| [2] | Boyer J S. Plant productivity and environment. Science, 1982, 218: 443-448. |
| [3] | Phoenix G K, Gwynn-Jones D, Callaghan T V, Sleep D, Lee J A. Effects of global change on a sub-Arctic heath: effects of enhanced UV-B radiation and increased summer precipitation. Journal of Ecology, 2001, 89(2): 256-267. |
| [4] | Vitousek P M. Beyond global warming: ecology an d global change. Ecology, 1994, 75: 1861-1876. |
| [5] | McCarty J P. Ecological consequences of recent climate change. Conservation Biology, 2001, 15: 320-331. |
| [6] | Smith S D, Huxman T E, Zitzer S F, Charlet T N, Housman D C, Coleman J S, Fenstermaker L K, Seemann J R, Nowak R S. Elevated CO2 increases productivity and invasive pieces success in an arid ecosystem. Nature, 2000, 408: 79-82. |
| [7] | Molles M C. Ecology: Concepts and Applications.New York: McGraw-Hill Companies, Inc, 2001: 344-345. |
| [8] | Horton J L, Kolb T E, Hart S C. Leaf gas exchange characteristics differ among Sonoran desert riparian trees pieces. Tree Physiology, 2001, 21: 233-241. |
| [9] | Zhang L X, Li W R, Bi Y R. Water availability affects photosynthetic gene expression in desert plant Ammopiptanthus mongolicus. Israel Journal of Plant Science, 2002, 50(4): 243-250. |
| [10] | Qian W, Zhu Y. Climate change in China from 1880 to 1998 and its impact on the environmental condition. Climatic Change, 2001, 50: 419-444. |
| [11] | Xu G Q, Wei W S. Climate change of Xinjiang and its impact on eco-Environment. Arid land geography, 2004, 27(1): 14-17. |
| [12] | Luo G P, Zhou C H, Chen X. Process of land use/land cover change in the oasis of arid region. Acta Geographica Sinica, 2003, 58(1): 63-72. |
| [13] | Jia Z Q, Lu Q, Guo B G. Progress in the study of Psarmnophyte Haloxylon. Forest Research, 2004, 17(1): 12-132. |
| [14] | Li H S, Zhang X L, Hou C X, Ma J. The study on diversity of adaptation of Haloxylon Ammodendron to drought. Arid Zone Research, 1995, 12(1): 15-17. |
| [15] | Su P X, Zhao A F, Zhang L X, Du W M, Chen H S. Characteristic in photosynthesis, transpiration and water use efficiency of Haloxylon ammodendron Calligonum mongolicum of desert.. Acta Botanica Boreali-occidentalia Sinica.2003, 23(1): 11-17. |
| [16] | Li X M, Natoli T, Kenji O. Comparative studies on relation of photosynthesis to water status of two species of Haloxylon under control environments. Acta Botanica Sinica, 1993, 35(10): 758-765. |
| [17] | Jiang T R, Zhang L X, Bi Y R, Jia X H, Fen J C, Tao L, Liu X M. Effects of water stress on gas exchange characteristics of Haloxylon Ammodendron leaves. Journal of LAN Zhou University (Natural Sciences), 2001, 37(6): 57-62. |
| [18] | Yang W B, Bao X F, Yang M R, Yang M. Research on relationship of physic-ecological water to drought resistance for Saxau1. Acta Ecologica Sinica, 1991, 11(4): 3l8-323. |
| [19] | Hou T Z, Liang Y Q. Research on physiological ecology of photosynthesis and water relation of Saxau/forests in Gua Jia Hu area of Xinjiang. Acta Phytoecologica et Geobotanica Sinica, 1991, 15(2): 141-150. |
| [20] | Xu H, Li Y, Ecophysiological response and morphological adjustment of Haloxylon ammodendron towards variation in summer precipitation. Acta Ecologica Sinica, 2007, 27(12): 5019-5028. |
| [21] | Xu H, Li Y. Water use strategies and corresponding leaf physiological performance of three deserts shrubs. Acta Botanica. Boreal-Occidental Sinica, 2005, 25(7): 1309-1316. |
| [22] | Duan R X, Jiang H F. Agricultural Meteorology. China Meteorological Press, 2002. |
| [23] | Donovan L A, Ehleringer J R. Water stress and use of summer precipitation in a Great Basin shrub community. Functional Ecology, 1994, 8: 289-297. |
| [24] | Li Y, Xu H, Cohen S. Long-term hydraulic acclimation to soil texture and radiation load in cotton. Plant, Cell and Environment, 2005, 28: 492-499. |
| [25] | Schwinning S, Ehleringer J R. Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. Journal of Ecology, 2001, 89: 464-480. |
| [26] | Sharpotou station of institute of desert research, Lanchow, CAS edited. Studies on Sand Dunes Controlled in Tengger Desert. Yingchuan: Ningxia People Press, 1991: 27-57. |
| [27] | EagIeson P S. Ecologica optimality in water limited natural soil vegetation systems. Water Resouwes Research, 1982, 18: 325-354. |
| [28] | Eissenstat D M, Caldwell M M. Competitive ability is linked to rates of water extraction. Oecologia, 1988, 75:1-7. |
| [29] | Tian Y, Li J G, Pan L P, Zhao Y. The key factors affecting Haloxylon ammodendron germination and survival at very early stage. Acta Ecologica Sinica, 2010, 30(18): 4898-4904. |
| [30] | Schulze E D, Mooney H A, Sala O E, Jobbagy E, Buchmann N, Bauer G, Canadell J, Jackson R B, Loret J, Desterheld M, Ehleringer J R. Rooting depth, water availability and vegetation cover along an aridity gradient in Patagonia. Oecologia, 1996, 108: 503-511. |
| [1] | 孙德祥, 李少熊, 钱拴提, 刘智宏, 韩刚, 汶录凤. 半荒漠风沙区人工柠条群落生物量动态及经济性状分析.西北林学院学报, 2005, 20(2): 24-27. |
| [11] | 徐贵青, 魏文寿. 新疆气候变化及其对生态环境的影响.干旱区地理, 2004, 27(1): 4-18. |
| [12] | 罗格平, 周成虎, 陈曦. 干旱区绿洲土地利用与覆被变化过程. 地理学报, 2003, 58(1): 63-72. |
| [13] | 贾志清, 卢琦, 郭保贵. 沙生植物一梭梭研究进展. 林业科学研究, 2004, 17(1):125-132. |
| [14] | 李洪山, 张晓岚.梭梭适应干旱环境的多样性研究. 干旱区研究, 1995, 12(1): 11-17. |
| [15] | 苏培玺, 赵爱芬, 张立新, 杜明武, 陈怀顺. 荒漠植物梭梭和沙拐枣光合作用、蒸腾作用及水分利用效率特征.西北植物学报, 2003, 23(1): 11-17. |
| [16] | 李小明, 名取俊树, 大政歉次. 人工环境下两种梭梭幼苗光合水分关系的比较研究. 植物学报, 1993, 35(10): 758-765. |
| [17] | 江天然, 张立新, 毕玉蓉, 贾小红, 冯金朝, 陶玲, 刘新民. 水分胁迫对梭梭叶片气体交换特征的影响. 兰州大学学报(自然科学版), 2001, 37(6): 57-62. |
| [18] | 杨文斌, 包雪峰, 杨茂仁, 杨明. 梭梭抗旱的生理生态水分关系研究. 生态学报, 1991, 11(4): 318-323. |
| [19] | 侯天侦, 梁远强. 新疆甘家湖梭梭林的光合、水分生理生态的研究. 植物生态学与地植物学学报, 1991, 15(2): 141-150. |
| [20] | 徐皓, 李彦, 邹婷, 谢静霞, 蒋理学. 梭梭生理与个体用水策略对降水改变的响应.生态学报, 2007, 27(12):5019-5028. |
| [21] | 许皓, 李彦. 3种荒漠灌木的用水策略及相关的叶片生理表现. 西北植物学报, 2005, 25(7): 1309-1316. |
| [22] | 段若溪, 姜会飞. 农业气象学. 北京: 气象出版社, 2002: 88-92. |
| [29] | 田媛, 李建贵, 潘丽萍, 赵岩.梭梭萌生与初期存活的关键影响因素.生态学报, 2010, 30(18): 4898-4904. |