 2014, Vol. 34
2014, Vol. 34文章信息
- 高福元, 赵成章, 卓马兰草
- GAO Fuyuan, ZHAO Chengzhang, MA Lancao
- 高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性
- Spatial distribution and spatial association of Stellera chamaejasme population in the different altitude in in degraded alpine grassland
- 生态学报, 2014, 34(3): 605-612
- Acta Ecologica Sinica, 2014, 34(3): 605-612
- http://dx.doi.org/10.5846/stxb201209191317
-
文章历史
- 收稿日期:2012-9-19
- 修订日期:2013-5-27






2. 西北师范大学地理与环境科学学院, 兰州 730070;
3. 甘肃民族师范学院, 甘南 747000
2. College of Geography and Environment Science, Northwest Normal University, Lanzhou 730070, China;
3. Gansu Normal University For Nationalities, gannan 747000, China
种群空间格局是指种群内个体的空间分布方式或配置特点,是植物种群在群落中相对位置可以定量化描述的基本特征之一,是种群生态特性、种内种间关系及生物与环境相互作用等综合作用的结果[1, 2]。植物种群的空间分布有3种基本类型:集群分布、随机分布和均匀分布,聚集分布体现了种群内部正向(相互有利)的生态关系,均匀分布反映了种群内部负向(相互排斥)的生态关系,随机分布则意味着种群内部没有明确的生态关系[3, 4, 5]。不同的格局类型可以反映出种群利用环境资源的状况,是其在群落中地位与生存能力的外在表现[6, 7]。从20世纪90年代,我国学者陆续运用点格局分析法研究森林或草地物种的种群分布格局或空间关联性,得以很好地从多尺度上理解植物种群的生态学特性或过程[8, 9, 10],同时借助“点格局”研究方法对退化草地中的优势种群分布格局和空间关联性进行了分析;王鑫厅[11]等利用摄影定位法结合点格局方法测定了羊草+大针茅草原退化群落中4个优势种群的空间分布格局,得到了退化草地中优势种群的一些重要特征。因此,种群空间分布格局的研究对于揭示草地退化机制具有十分重要的意义。
狼毒(Stellera chamaejasme L.)是广泛分布于我国北方地区的一种毒杂草。由于全球气候变化以及对草地的过度利用,狼毒种群增长迅速,严重威胁到天然草地中优良牧草的生长,受到生态学家的重视,关于狼毒的研究包括不同退化梯度下狼毒种群土壤种子库[12];狼毒种群土壤中养分的变化研究[13];狼毒个体年龄的判断方法及其种群的年龄结构研究[14]。同时,邢福[15, 16]等对内蒙古典型草原区狼毒种群的空间分布格局和种子种群分布格局强度、类型与规模进行了分析,认为狼毒种群具有“近母株散布”的种子散布机制,基本散布半径为0—50cm,并引起生物学斑块。随着“点格局”方法在种群格局分析中普遍应用,赵成章[17, 18]等对不同分盖度下的狼毒种群和不同龄级的狼毒种群进行了点格局分析,并得出了种群的扩散规律。近年来,狼毒种群的分布区域不断扩大,在一些高海拔地区定植、繁殖、扩散,形成重度退化草地,但对于狼毒种群能够在高海拔地区生存、扩散的原因和机理等方面的研究较薄弱,阻碍了对于狼毒大量滋生的退化草地的全面认识。鉴于此,本文采用点格局分析方法,通过分析不同海拔高度狼毒种群的大小组成、空间分布格局和空间关联性,探讨了狼毒种群空间格局和空间关联性对海拔高度的响应,为深刻理解狼毒种群对环境变化的响应机制和“狼毒型”退化草地分布区扩散机制提供科学参考。
1 材料与方法 1.1 研究区概况石羊河是甘肃河西走廊三大内陆河流之一,发源于祁连山区,恶劣的自然条件和人类不合理开发利用,使该流域已成为我国西北内陆河流域生态环境问题最严重的区域,上游天然草地生态系统发生了严重退化,对祁连山地、河西走廊和青藏高原的陆地生态系统产生了重要影响。研究区位于祁连山北坡中山区皇城草原(37°51′ N,101°50′ E),海拔2700—3000m,年均温1—2 ℃,年平均日照时数2800 h,无霜期90 d,年降水量320—400 mm,蒸发量1500—1800 mm,相对湿度65%,土壤为山地栗钙土,pH值8.3。研究区属山地草原,狼毒(Stellera chamaejasme)斑块分布,伴生有赖草(Leymus secalinus)、克氏针茅(Stipa krylovii)、阿尔泰狗哇花(Aster tataricus)、冷蒿(Artemisia frigida)、扁穗冰草(Agropyron cristatum)、苔草(Carex tristachya)等,受干旱气候影响,草地生产能力处于较低水平[19]。
1.2 样地设置2012年8月,在研究区内选择坡度(38—40°)、坡向(南坡)相同的山坡,沿海拔2700—3000 m,每隔100 m海拔高度设置一个样带,共设置4个水平样带,每个样带内选取71%—80%狼毒分盖度下的重度退化草地。根据以前对狼毒种群分布格局的研究[17, 18]和狼毒自生的特征,2 m×2 m的样方能够满足实验的要求,并且使种群的各种格局均能出现,因此在各样带分别设置71%—80%狼毒分盖度下的2 m×2 m的正方形样地1个。
1.3 取样方法 1.3.1 点格局取样方法点格局分析取样要求取一个样地。在上述样地内设置邻接格子样方,每个样地用4个1 m×1 m的样方框(样方框内小格子的面积为10 cm×10 cm) 按从左到右,从上到下的顺序依次放置放置4次进行取样,记录样方内每个狼毒植株的地面投影和分枝数,在样方中记录每个狼毒个体的位置,以坐标值表示,用坐标值直接表示距离[20]。在室内对每一个样方内提取的狼毒个体的相对位置图(每个样地4个),按照取样时样方框放置的顺序将4个图进行拼接,合并成2 m×2 m的正方形样地所对应的图。按枝条数将狼毒株丛划分为4级:Ⅰ级株丛(枝条数1—10枝)、Ⅱ级株丛(枝条数11—20枝)、Ⅲ级株丛(枝条数21—30枝)、Ⅳ级株丛(30以上),并统计每个样方内的各级狼毒个体数量。
1.4 数据分析 1.4.1 点格局分析研究区域内,位于二维空间的点构成点事件,用k(d)函数来反映点事件空间布局的依赖程度[21],其定义如下:
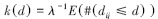
式中,λ表示研究区域内每单位面积点事件的密度,E表示在一定距离尺度下点事件数量的期望,#表示数量,dij表示研究区域内任意一点到其他点的距离,d表示距离尺度。如果在一定距离尺度下点事件的分布是随机的,则有:

此时,k(d)=πd2。当点事件为聚集分布时,如果距离尺度较小,k(d)的观测值将大于πd2。估计式为:
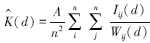
式中,A表示研究区域面积,n是研究区域内点事件的数量,i、j是样方内任意两个具有相同特征的点事件,Iij=1(当dij≤d成立时,dij是i到j之间的距离),wij(d)是边缘校正的权重,等于以i为圆心,以dij为半径的圆落在研究区域内的弧长和整个圆周长的比值。
通常用L(d)函数的估计值来判断在此尺度下点事件的分布类型。在距离尺度d下,如果L(d)>0,为聚集分布;如果L(d)=0,为随机分布;如果L(d)<0,则为均匀分布:
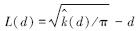
一般利用Monte Carlo方法来求解随机分布的上下包迹线,如果研究对象实际的值落在上下包迹线的区间内,则在此距离尺度下植物种群是随机分布的;如果研究对象实际的值落在上下包迹线的区间以上,则在此距离尺度下植物种群是聚集分布的;如果研究对象实际的值落在上下包迹线的区间以下,则在此距离尺度下植物种群是均匀分布的[7]。具体方法是选取m组n个随机数,每组都利用上述公式求解在不同距离尺度下的L(d)值,然后把对应的每个d下的m个L(d)由小到大进行排序,分别保留最大值和最小值作为上下包迹线的取值范围。
1.4.2 空间关联性两个种群之间的空间关联性分析,实际上是两个种群之间的点格局分析,也叫多元点格局分析。单种格局分析可以认为是一个种群个体间的关系研究[7],因此对第1个种群k(d)可以写成k11(d),对第2个种群可以写成k22(d)。现在要考虑两个不同大小种群的个体在距离(尺度)d内的数目,就是要求k12(d),其定义和计算原理与单种格局相近。不难证明k12(d)可以用下式估计[22]:
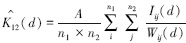
这里n1和n2分别为种群1和种群2的个体数(点数),A、Iij(d)和Wij(d)含义同(1)式,不同的是i和j分别代表种群1和种群2的个体,同样计算:
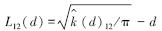
当L12(d)=0表明两个种在d尺度下无关联性,当L12(d)>0表明二者为正关联,当L12(d)<0表明二者为负关联。用Monte-Carlo检验拟合包迹线,以检验两个种是否显著地关联。如果研究对象实际值落在上下包迹线的区间内,则在此距离尺度下两个植物种群空间关联性不显著;如果研究对象实际的值落在上下包迹线的区间以上,则在此距离尺度下两个植物种群空间关联性为显著正关联;如果研究对象实际的值落在上下包迹线的区间以下,则在此距离尺度下两个植物种群空间关联性为显著负关联。采用GetData Graph Digitizer 2.22进行数据采集,Visual Fortran 6.5进行编程计算,Origin 7.5进行绘图。本文中坐标原点为样方左下角的交点,采用的步长为5 cm,Monte-Carlo随机模拟的数目为20次,得到上下两条包迹线围成的95%置信区间。
2 结果与分析 2.1 狼毒种群的组成格局在坡度、坡向、盖度相同的条件下,随着海拔升高Ⅰ、Ⅱ级狼毒株丛个体数逐渐减少,Ⅲ、Ⅳ级狼毒株丛个体数逐渐增多。在所有海拔梯度上,Ⅰ级狼毒株丛个体数最多,Ⅳ级狼毒株丛个体数最少。
|
海拔/m
Altitude | 株级/株 Class/Plant | |||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | |
| 2700 | 124 | 21 | 4 | 3 |
| 2800 | 108 | 17 | 5 | 3 |
| 2900 | 92 | 15 | 5 | 4 |
| 3000 | 74 | 13 | 6 | 5 |
在2700、2800、2900m和3000m海拔高度上,Ⅰ级狼毒株丛分别在0—82cm、0—75cm、0—70cm和0—62cm尺度范围内表现为聚集分布,聚集强度随海拔升高逐渐增强,在80—100cm、75—100cm、70—100cm和62—100cm尺度范围内表现为随机分布;Ⅱ级狼毒株丛分别在0—80cm、0—73cm、0—68cm和0—61cm尺度范围内表现为聚集分布,聚集强度也随海拔升高逐渐增强,在80—100cm、73—100cm、68—100cm和61—100cm尺度范围内表现为随机分布;Ⅲ、Ⅳ级狼毒株丛在所有研究尺度上以随机分布为主。
2.3 不同狼毒株丛之间的空间关联性在2700、2800、2900m和3000m海拔高度上(图2),Ⅰ和Ⅱ级狼毒株丛在0—100cm尺度上均表现为负关联;Ⅰ和Ⅲ级狼毒株丛分别在0—50cm、0—42cm、0—38cm和0—36cm尺度上为正关联,在50—100cm、42—100、38—100cm和36—100cm尺度上表现为无关联;Ⅰ和Ⅳ级狼毒株丛分别在0—55cm、0—44cm、0—40cm和0—38cm尺度上表现为正关联,在55—100cm、44—100cm、40—100cm和38—100cm尺度上表现为无关联;Ⅱ和Ⅲ级狼毒株丛在0—100cm尺度上均表现为负关联,负关联强度随海拔升高逐渐减弱。Ⅱ和Ⅳ级狼毒株丛,2700、2800m海拔高度上所有尺度上表现为无关联,2900m和3000m海拔高度上,分别在0—38cm和0—34cm尺度上表现为正关联,随海拔升高关联强度增强,分别在38—100cm和34—100cm尺度上表现为无关联。Ⅲ和Ⅳ级狼毒株丛在所有海拔高度的所有尺度上表现为负关联。
3 讨论尤海舟[23]等通过对川滇高山栎种群2400—3600m海拔高度上种群数量分布的研究表明高山栎植株数量随着海拔梯度的升高呈现先升高后降低的趋势,2800m是高山栎分布最多的海拔,并推断2800m海拔的环境比较适合高山栎的生存。与高山栎种群数量分布相比,狼毒种群数量分布规律较为复杂,随海拔变化不同株级表现出不同的变化规律。在坡度、坡向和狼毒分盖度相同的条件下,狼毒种群随着海拔升高幼小个体数逐渐减少,这是狼毒种群对环境条件变化的响应,一方面是繁殖投入方面的原因,有研究表明种群间的繁殖对策和繁殖成功会随海拔发生变化[24, 25],随着海拔升高温度降低,风速增大,为了个体和种群的生存,成株个体可能会放弃一些繁殖机会,减少繁殖不仅可以增加成株个体生存的机率,同时减小了种内竞争的强度,使部分幼株能够生存,对种群的发展具有重要意义;另一方面由于环境的变化,使得幼小个体的存活率降低,因此随海拔升高幼小个体数减少。随海拔升高成株个体数 逐渐增多,这是因为随海拔升高环境条件改变,幼小个体数量减少,生存机率也降低,导致相同龄级的狼毒个体数减少,有效的减小了种内竞争的压力,幼株个体只要能够克服外界环境生存下来,生长为较大成株个体的概率较大;在低海拔地区虽然幼株个体生存概率较大,但是相同龄级的个体数较多,种内竞争压力增大,向更大狼毒过渡时死亡率高,所以随海拔升高较大的成株个体数逐渐增多。
由于大龄级个体由中小龄级发育而来,大龄级个体还影响着其他龄级个体的存活和生长,所以分析不同龄级个体间的空间格局和种间关联可以更好地理解群落的时空动态和小龄级个体生长过程的影响因素[26]。Hegazy[27]等认为典型山地垂直地带上的植物种群空间格局会随海拔变化而变化,结合图1、图2资料,在高寒草地狼毒种群的空间分布格局与空间关联性随海拔表现出规律变化。Ⅰ级狼毒株丛在低海拔地区小尺度(0—50 cm)上表现为聚集分布,随海拔的升高聚集强度增强,聚集尺度减小,Ⅰ级狼毒株丛与Ⅲ、Ⅳ级狼毒株丛小尺度上表现为正关联关系,并且随着海拔的升高,正关联关系发生的尺度减小,一方面是因为Ⅲ、Ⅳ级狼毒株丛能够为Ⅰ级狼毒株丛抵挡风沙,对幼小个体起到保护作用,二者之间形成了正关联关系[28],同时形成以Ⅲ、Ⅳ级狼毒个体为中心的狼毒斑块,表现出聚集分布[17, 18],低海拔地区热量好,风速小,Ⅲ、Ⅳ级狼毒个体能够对较多的幼小个体提供保护作用,形成斑块的尺度较大,所以形成聚集分布的尺度较大,随着海拔升高,环境条件发生变化,热量减少,风速变大,Ⅲ、Ⅳ级狼毒自身所处的环境也相对恶劣,需要自我保护,所以形成斑块的尺度变小,正关联关系发生的尺度减小,聚集分布的尺度相应减小,但是强度增强;另一方,幼小个体对水分等资源的竞争相对微弱,同时为抵御风沙和提高存活率,它们需要相互庇护,于是个体之间以正向相互关系为主,进而表现为聚集分布[29, 30]。Ⅰ级狼毒株丛与Ⅱ级狼毒株丛在小尺度上,所有海拔上表现为负关联,这是因为他们处于同一个斑块内,种内竞争引起负关联。

|
| 图1 不同海拔高度狼毒株丛点格局分析 Fig. 1 Results of point pattern analysis for Stellera chamaejasme plants in the different altitude A、B、C、D表示2700mⅠ、Ⅱ、Ⅲ、Ⅳ级狼毒株丛的格局;E、F、G、H表示2800mⅠ、Ⅱ、Ⅲ、Ⅳ级狼毒株丛的格局;I、J、K、L表示2900mⅠ、Ⅱ、Ⅲ、Ⅳ级狼毒株丛的格局;M、N、O、P表示3000mⅠ、Ⅱ、Ⅲ、Ⅳ级狼毒株丛的格局 |
Ⅱ级狼毒株丛在所有海拔上,小尺度范围内表现为聚集分布,并随海拔升高聚集强度增强,聚集尺度减小,但是聚集强度明显低于Ⅰ级狼毒株丛。Ⅱ级狼毒株丛与Ⅲ级狼毒株丛之间表现为负关联,在低海拔地区与Ⅳ级狼毒株丛之间表现为无关联,在高海拔地区表现为正关联,强度较弱,随海拔升高正关联强度增强。这说明Ⅱ级狼毒株丛较Ⅰ级狼毒株丛具有更强的竞争力和自我保护、生存的能力,在低海拔地区Ⅱ级狼毒株丛已经能够适应环境,逐渐能够独立生存,聚集强度降低,分布格局有向随机分布转化的趋势,并开始逐渐和Ⅲ级狼毒株丛竞争资源,导致二者之间表现为负关联,在高海拔地区Ⅳ级狼毒株丛对Ⅱ级狼毒株丛的庇护作用依然非常明显,二者之间仍然表现为正关联。
Ⅲ和Ⅳ级狼毒株丛形成随机分布和负关联关系可能有两个原因:1)个体之间具有较强的竞争能力,相互之间的竞争导致负关联和随机分布;2)Ⅲ和Ⅳ级狼毒株丛年龄、体积都相对较大,它们已经占据了一定的空间资源,逐渐形成了以自己为核心的狼毒斑块,斑块间的竞争导致随机分布[16]以及负关联关系,同时这也是Ⅰ、Ⅱ级狼毒株丛在较大尺度上表现为随机分布和负关联的原因。

|
| 图2 不同海拔高度狼毒株丛之间的关联性分析 Fig. 2 Spatial association analysis between Stellera chamaejasme plants in the different altitude a、b、c、d表示Ⅰ和Ⅱ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性;e、f、g、h表示Ⅰ和Ⅲ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性;i、j、k、l表示Ⅰ和Ⅳ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性;m、n、o、p表示Ⅱ和Ⅲ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性;q、r、s、t表示Ⅱ和Ⅳ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性;u、v、w、x表示Ⅲ和Ⅳ级狼毒株丛在2700、2800、2900m和3000m海拔高度上的关联性 |
综上所述,对不同海拔条件下不同大小狼毒株丛空间关联性和空间分布格局的研究说明,狼毒种群对环境变化具有明显的响应机制,在所有海拔高度上较大的成株对较小的幼株具有有效的保护作用[26],在高海拔地区这种作用更加突出,强度更强。狼毒种群能够通过减少繁殖、减弱同龄级狼毒之间的竞争以及提高种群的聚集强度等策略来适应高海拔地区的环境,这是狼毒种群能够在高海拔地区生存-繁殖-扩散的内在机制,也是狼毒种群能够在西北干旱区形成优势种的重要条件。
| [1] | Qu Z X. Plant Ecology. 2nd ed. Beijing: Higher Education Press, 1983. |
| [2] | Cao G X, Zhong Z C, Liu Y. The study of distribution pattern of Camellia rosthorniana population in Jinyun Mountain. Journal of Biology, 2003, 20(1): 10-12. |
| [3] | Philips D L,Macmahon J A.Competition and spacing patterns in desert shrubs.Journal of Ecology,1981, (69):97-115. |
| [4] | Kenkel N C.Pattern of self thinning in Jack pine:testing the random mortality hypothesis.Ecology,1988, (69):1017-1024. |
| [5] | Manuel C M. Ecology: Concept and Application. Science Press&McGraw_Hill, Beijing 2000. |
| [6] | Dale M R T. Spatial Pattern Analysis in Plant ecology. Cambridge: Cambridge University Press, 1999. |
| [7] | Zhang J T. Analysis of spatial point pattern for plant species. Chinese Journal of plant Ecology, 1998, 22(4): 344-349. |
| [8] | Yang H X, Zhang J T, Wu B, Li X S, Zhang Y Y. Point pattern analysis of Artemisia ordosica population in theMu us sandy land. Chinese Journal of plant Ecology, 2006,30(4):563-570. |
| [9] | Liu Z G, Li Z Q. Fine-scale spatial pattern of Artemisia frigida population under different grazing intensities. Acta Ecologica Sinica, 2004, 24(2): 227-234. |
| [10] | Zhang J T, Meng D P. Spatial pattern analysis of individuals in different age-classes of Larix principis-rupprechtii in Luya Mountain Reserve, Shanxi, China. Acta Ecologica Sinica, 2004, 24(1): 35-40. |
| [11] | Wang X T, Hou Y L, Liu F, Chang Y, Wang W, Liang C Z, Miao B L. Point pattern analysis of dominant populations in a degraded community in Leymus chinensis + Stipa grandis steppe in Inner Mongolia, China. Chinese Journal of plant Ecology, 2011, 35 (12): 1281-1289. |
| [12] | Zhao C Z,Zhang Q P,Dong X G. Soil seed bank of Potentilla acaulis community in arid grassland of the upstream of Shiyang River.Arid Land Geography,2010,33(1):71-76. |
| [13] | G Sun, P Luo, N Wu.Stellera chamaejasme L.increases soil N availability, turnover rates and microbial biomass in an alpine meadow ecosystem on the eastern Tibetan Plateau of China.Soil Biology and Biochemistry,2009,41:86-91. |
| [14] | Xing F,Guo J X,Wei C Y. Judging method of individual age and age structure of Stellera chamaejasme population in degraded steppe. Chinese Journal of Applied Ecology,2004,15(11):2104-2108. |
| [15] | Xing F, Song R. Population distribution pattern and dynamics of poisonous Stellera chamaejasme on grassland. Pratacul -tural Sinica, 2002, 19(1): 16-19. |
| [16] | Xing F, Wang Y H, Guo J X. Spatial distribution patterns and dispersal mechanisms of the seed population of Stellera chamaejasmeon degraded grasslands in Inner Mongolia,China. Acta Ecologica Sinica,2004, 24(1): 143-148. |
| [17] | Zhao C Z, Gao F Y, Wang X P, Sheng Y P, Shi F X. Fine-scale spatial patterns of Stellera chamaejasme population in degraded alpine grassland in upper reaches of Heihe, China. Chinese Journal of plant Ecology, 2010, 34(11):1319-1326. |
| [18] | Zhao C Z, Gao F Y, Sheng Y P, Dong X G, Zhou W.Fine-scale spatial distribution and spatial association of Stellera chamaejasme population.Arid Land Geography, 2011, 34(3):492-498. |
| [19] | Zhao C Z,Fan S Y,Yin C Q.Study on Vegetation Community's Structure of Degraded Grassland of Noxious and Miscellaneous Grass Type. Journal of Desert Research,2004,24(4):507-512. |
| [20] | Riggle B D. Spatial statistics,Wiley, New York, 1981. |
| [21] | Manly B F J. Randomization and Monte Carlo Methods in Biology. Chapman and Hal, New York.1991. |
| [22] | Diggle P J.Statistical analysis of spatial point patterns, Academic Press, NewYork, 1983. |
| [23] | You H Z, Liu X l,Miao N, He F,Ma Q Y. Individual association and scale effect of spatial pattern of Quercus aquifolioides populations along the elevation gradients. Acta Ecologica Sinica, 2010,30(15):4004-4011. |
| [24] | Bingham R A,Orthner A R.Efficient pollination of alpine plants.Nature,1998,391:238-239. |
| [25] | Kudo G Molau U.Variations in reproductive traits at florescence and flower levels of an arctic legume.Astragalus alpinus L Comparisons between asubalpine and an alpine population.Plant Species Biology,1999,14:181-191. |
| [26] | Li L, Wei S G, Huang Z L. Spatial patterns and interspecific associations of three canopy species at different life stages in a subtropical forest, China.Journal of Integrative Plant Biology, 2008,50: 1140-1150. |
| [27] | Hegazy A K,Hosni H A,El-Demerdash M A.Vegetation,species diversity and floristic relations along an altitudinal gradient in south-west SaudiArabia.Journal of Arid Environment,1998,38:3-13. |
| [28] | Gao F Y, Zhao C Z. Pattern-controlling mechanics of different age classes of Stellera chamaejasme population in degraded alpine grassland. Acta Ecologica Sinica,2013,33(10): 3114-3121. |
| [29] | Allee W C.Animal Aggregations:A study in general sociology.Chicago: University of Chicago Press,1931: 41-65. |
| [30] | Cai F.A study on the structure and dynamics of Cyclobalanopsis glauca population at hills around west lake in HangZhou. Scientia Silvae Sicinae,2000,36(3):67-72. |
| [1] | 曲仲湘. 植物生态学 (第二版).北京: 高等教育出版社, 1983. |
| [2] | 操国兴, 钟章成, 刘芸, 谢德体. 缙云山川鄂连蕊茶种群空间分布格局研究.生物学杂志, 2003, 20(1): 10-12. |
| [7] | 张金屯. 植物种群空间分布的点格局分析.植物生态学报, 1998, 22(4): 344-349. |
| [8] | 杨洪晓,张金屯,吴波,李晓松,张友炎.毛乌素沙地油蒿种群点格局分析.植物生态学报,2006,30(4):563-570. |
| [9] | 刘振国,李镇清.不同放牧强度下冷蒿种群小尺度空间格局.生态学报,2004,24(2):227-234. |
| [10] | 张金屯,孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析.生态学报, 2004, 24(1): 35-40. |
| [11] | 王鑫厅,侯亚丽,刘芳,常英,王炜,梁存柱,苗百岭.羊草+大针茅草原退化群落优势种群空间点格局分析.植物生态学报, 2011, 35 (12): 1281-1289. |
| [12] | 赵成章,张起鹏,董小刚,等.石羊河上游干旱草原星毛委陵菜群落土壤种子库.干旱区地理,2010,33(1):71-76. |
| [14] | 邢福,郭继勋,魏春雁. 退化草原狼毒个体年龄判定方法及其种群年龄结构的研究.应用生态学报,2004,15(11):2104-2108. |
| [15] | 邢福,宋日.草地有毒植物狼毒种群分布格局及动态.草业科学,2002,19(1):16-19. |
| [16] | 邢福,王艳红,郭继勋.内蒙古退化草原狼毒种子的种群分布格局与散布机制.生态学报, 2004, 24(1): 143-148. |
| [17] | 赵成章,高福元,王小鹏,盛亚萍,石福习.黑河上游高寒草地狼毒种群小尺度点格局分析.植物生态学报,2010,34(11):1319-1326. |
| [18] | 赵成章,高福元,盛亚萍,董小刚,周伟.狼毒种群小尺度空间分布格局及空间关联性研究.干旱区地理,2011,34(5):492-497. |
| [19] | 赵成章,樊胜岳,殷翠琴.祁连山区天然草原退化原因分析与可持续利用对策.中国沙漠,2004,24(2):207-210. |
| [23] | 尤海舟,刘兴良,缪宁,何飞,马钦彦. 川滇高山栎种群不同海拔空间格局的尺度效应及个体间空间关联.生态学报, 2010,30(15):4004-4011. |
| [28] | 高福元,赵成章. 高寒退化草地狼毒种群株丛间格局控制机理.生态学报, 2013,33(10): 3114-3121. |
| [30] | 蔡飞. 杭州西湖山区青冈种群结构和动态的研究.林业科学, 2000, 36(3): 67-72. |