 2014, Vol. 34
2014, Vol. 34文章信息
- 吴寿国, 余学军, 李凯, 蒋玉俭, 张汝民
- WU Shouguo, YU Xuejun, LI Kai, JIANG Yujian, ZHANG Rumin
- 海盐对绿竹叶片反射光谱及叶绿素荧光参数的影响
- Effects of sea salt stress on reflectance spectrum and chlorophyll fluorescence parameters in Bambusa oldhamii leaves
- 生态学报, 2014, 34(17): 4920-4930
- Acta Ecologica Sinica, 2014, 34(17): 4920-4930
- http://dx.doi.org/10.5846/stxb201309232337
-
文章历史
- 收稿日期:2013-9-23
- 修订日期:2014-7-6






2. 瑞安市林业局, 瑞安 325200
2. Forest Enterprise of Ruian City, Ruian 325200, China
沿海防护林是海岸生态系统中最基本的生物资源,由于沿海地区地下水位高,土壤含盐量高,肥力低,生态环境恶劣等特点[1],所以优良植物种选择是加快沿海优良防护林体系建设的关键。光谱和叶绿素荧光技术具有无损伤、快速、高效的优越性,已在监测植物胁迫过程中得到广泛的应用[2, 3, 4, 5]。张丽平等研究发现盐胁迫处理不同品种的黄瓜 (Cucumis sativus),其叶片在可见光区的光谱反射率明显增加,改良归一化差值指数 (modified red-edge ratio,mSR705)、改良红边比值指数 (modified red-edge normalized difference vegetation,mND705) 和光化学反射指数 (photochemical reflectance index,PRI) 与黄瓜的盐害程度及不同品种的耐盐程度有密切关系[3]。随着盐浓度增加,蓖麻子 (Ricinus communis) 叶片的光谱反射率在可见光范围内与叶绿素含量成反相关,最大荧光强度 (maximal fluorescence intensity,Fm)、光系统Ⅱ (photosystemⅡ,PSⅡ) 最大量子产率 (maximum quantum yield of PSⅡ,Fv/Fm) 和PSⅡ实际量子产量 (actual photochemical efficiency of PSⅡ,ΦPSⅡ) 等叶绿素荧光参数逐渐降低[6]。刘炳响等认为盐胁迫使白榆 (Ulmus pumila) 叶片电子传递的量子产额 (quantum yield for electron transport,ΦEo)、单位面积内反应中心的数量 (density of reaction centers,RC/CS)和叶片性能指数 (performance index of absorption basis,PIABS) 降低[7]。小麦 (Triticum aestivuml) 叶片Fv/Fm、单位反应中心吸收的光能 (absorption flux per reaction center,ABS/RC) 以及PIABS的降低来适应盐胁迫对其伤害[8]。
绿竹 (Bambusa oldhamii),禾本科绿竹属。绿竹笋是夏秋季节群众喜爱的蔬菜,俗称最美味的笋。绿竹根系发达庞大,耐水力强,能固土护岸、保持水土等。目前,绿竹的研究主要集中在光合[9]、保护酶活性[10]、基因克隆[11]、笋营养成分[12]和组织培养[13]等方面,但是利用荧光和反射光谱对其耐盐的研究较少。本研究采用Unispec-SC型单通道光纤光谱仪和非调制式叶绿素荧光仪对绿竹叶片反射光谱和叶绿素快速荧光参数进行测定,分析了绿竹反射光谱和叶绿素荧光参数对盐胁迫的响应特征,为今后绿竹的栽培应用及优良品种选育奠定基础。
1 材料与方法 1.1 实验材料及材料培养供试材料为绿竹2 年生组培苗(第1 年生长在组培室,第2 年移栽至温室大棚),缓苗期使用水培方式,选用1/2 Yoshida营养液配方[14]。选择生长健康,大小基本一致 (高80—100 cm,鲜质量90—110 g) 的绿竹苗,并用清水冲洗植物根部,在茎基部位裹海绵条,然后将苗塞进桶盖的孔中。装入10 L的红色塑料桶内 (桶盖上均匀烙3个直径1 cm左右的孔),红桶外在10 L处做好标记,桶外用黑色塑料袋封住,每桶加一个气泵,保持每天持续通气,并调节pH 5.0。每6—7 d更换1 次培养液,每天观察桶内水分情况,蒸发的水分及时补充,绿竹在水培条件下适应20 d左右后 (自然光照,湿度80%,日平均温28 ℃),待用。
1.2 人工海水的配制胁迫用的海盐按照Mocledon的人工海水配方配制 (表 1)[15]。先依照Mocledon人工海水配方配制成母液,将母液和1/4 Yoshida营养液混合,再按照所需的海盐胁迫浓度配制成模拟海盐溶液。胁迫过程的管理同缓苗期。
| 成分Composition | ||||||||
| NaCl | MgCl2 | MgSO4 | CaCl2 | NaHCO3 | KCl | NaBr | H3BO3 | |
| * 数字来源http://zhidao.baidu.com/question/26278104.htmL?fr=qrl | ||||||||
| 含量Content/(g/L) | 26.726 | 2. 226 | 3.248 | 1.153 | 0.198 | 0.721 | 0.058 | 0.058 |
实验时间为2012年8月上旬,地点选择在浙江农林大学智能楼温室大棚,自然光照,湿度控制在80%,日平均温度控制在28 ℃。待用苗采用随机分组,海盐浓度分别为0 (对照)、0.4% (Tr1)、0.8% (Tr2)、1.2% (Tr3)、1.6% (Tr4),设置5个处理,每个处理5个重复,每个重复3丛绿竹苗。胁迫处理15 d后开始各指标测定。选晴朗、无风天气,10:00—12:00进行测量。每丛选取顶端的第3或第4片光照充分生长状况良好的无病斑叶片,采样点1个/叶。
1.4 实验方法 1.4.1 叶绿素含量的测定去除叶脉后剪碎混匀后称取0.1 g,加入80%丙酮溶液5 mL,遮光室温萃取,叶片发白后测量,每个处理3个重复。用紫外-可见分光光度计UV-2500 (日本岛津)测定在470、645、663 nm波长下的吸光度值。采用Lichtenthaler[16]的公式计算叶绿素a (Ca)、叶绿素b (Cb)和类胡萝卜素含量 (Car)。公式如下:
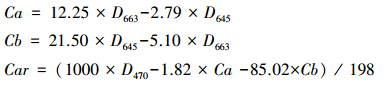
采用仪器为光谱分析仪 (UniSpec-SC,US),波段设置310—1150 nm,采样间隔1 nm,光谱分辨率1 nm。每个处理选取3个样株,每个样株选取5 片叶子,即每个处理重复15 次,取平均值作为该样品叶片光谱反射率。测量过程中及时进行标准白板校正。用Multispec 5.1数据处理软件读取反射光谱原始数据。
1.4.3 叶绿素荧光参数的测定采用非调制式叶绿素荧光仪 (Yaxin-1161型,北京雅欣理仪科技有限公司,北京) 进行叶片叶绿素荧光动力学参数的测定。叶片暗适应10 min后,选取叶肉部分用3000 μmol m-2 s-1饱和蓝闪光照射1 s,以10 μs (2 ms之前) 和1 ms (2 ms之后) 的间隔记录荧光信号,测得叶绿素荧光动力学参数。每株测定3次,计算平均值和标准误差。
1.5 数据处理 1.5.1 光谱分析将毛竹叶片反射光谱通过下列公式进行一阶微分处理得到微分光谱。
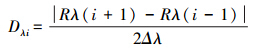
“三边”参数的计算方法:分别在505—545、560—640 nm和680—750 nm范围内确定蓝边、黄边和红边位置、幅值和面积。红边位置 (red wavelength,λred) 为红光范围内一阶导数光谱最大值所对应的波长,红边幅值 ( Dλred) 为一阶导数光谱的最大值,红边面积 (Sred) 为一阶导数光谱线所包围的面积。黄边和蓝边参数与红边参数意义类似,黄边位置 (λyellow),黄边幅值 (Dλyellow),黄边面积 (Syellow);蓝边位置 (λblue),蓝边幅值 (Dλblue),蓝边面积 (blue area,Sblue)。根据叶片特征,筛选出较能反应叶片特征结构的光谱参数及相关公式见表 2。
| 反射光谱参数 Reflectance spectrum parameters | 定义 Definition | 文献来源 Reference |
| 公式中: R代表反射,下标代表光谱波段或波长 | ||
| 红边归一化指数 Red-edge normalized difference vegetation index | 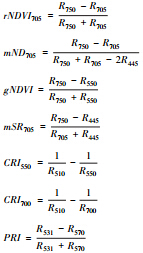 | [17] |
| 改良红边比值指数 Modified red-edge normalized difference vegetation index | [17] | |
| 绿度归一化指数 Greenness normalized index | [18] | |
| 改良归一化差值指数 Modified red-edge ratio | [17] | |
| 类胡萝卜素反射指数1 Carotenoid reflectance index Ⅰ | [19] | |
| 类胡萝卜素反射指数2 Carotenoid reflectance index Ⅱ | [19] | |
| 光化学反射指数 Photochemical reflectance index | [20] | |
参考Strasser等[21]的计算方法。下述公式中,Fm表示暗适应后的最大荧光强度;Fo表示暗适应后的最小荧光强度;F300μs 表示在暗适应后照光300 μs时的荧光强度;FJ表示在暗适应后照光2 ms时的荧光强度。2 ms时的可变荧光VJ = (FJ-Fo) / (Fm-Fo);荧光曲线的相对初始斜率Mo = 4 × (F300μs-Fo) / (Fm-Fo)。暗适应下PSⅡ最大量子产率Fv/Fm = (Fm-Fo) / Fm;单位面积反应中心数目RC / CS = (Fv / Fm) × (VJ / Mo) × Fo;单位反应中心复合体吸收的能量ABS / RC = Mo / VJ / (Fv / Fm);捕获的激子将电子传递到电子传递链中QA-下游的其他电子受体的概率Ψo = (1-VJ);电子传递的量子产额ΦEo = (1-Fo / Fm) × Ψo;用于电子传递的能量ETo / RC = Mo × (1 / VJ) × Ψo;叶片性能指数PIABS =1/ ABS / RC× [Fv/Fm / (1-Fv/Fm)] × [Ψo /(1-Ψo)]。
1.5.3 数据处理采用Origin 8.0软件进行数据统计分析和绘图。
2 结果与分析 2.1 不同海盐浓度胁迫对光合色素含量的影响海盐胁迫下了绿竹叶片光合色素含量的变化如表 3显示。从表 3可以看出,随着海盐浓度的增大,叶绿素a、叶绿素b和类胡萝卜素含量均出现先增大后下降的趋势;海盐浓度小于1.2%时,叶绿素总量和类胡萝卜素与对照无显著差异,当海盐浓度增加到1.6%时,叶绿素a、叶绿素b和类胡萝卜素含量均呈极显著的降低,与对照相比分别降低了63.2%、62.8%、47.2%。叶绿素a/b值在0.4%浓度处理下显著降低,比对照降低了12.7%。
| 海盐浓度/% Sea salt concentration | 叶绿素a/(mg/g) Chlorophyll a | 叶绿素b/(mg/g) Chlorophyll b | 叶绿素总量/(mg/g) Chlorophyll a+b | 类胡萝卜素/(mg/g) Carotenoids | 叶绿素a/b Chlorophyll a/b |
| 相同字母表示差异不显著;不同小写字母表示差异显著 (P< 0.05);不同大写字母表示差异极显著 (P < 0.01) | |||||
| 0 (CK) | 2.20±0.25Aab | 0.97±0.15Aab | 3.14±0.40ABab | 0.53±0.09Aab | 2.37±0.11Aa |
| 0.40 | 2.39±0.32Aa | 1.16±0.17Aa | 3.54±0.49Aa | 0.62±0.00Aa | 2.07±0.02Bb |
| 0.80 | 2.19±0.22Ab | 0.93±0.10Ab | 3.13±0.31ABab | 0.59±0.05Aa | 2.35±0.03Aa |
| 1.20 | 2.01±0.35Ac | 0.86±0.14Ac | 2.87±0.49Cc | 0.52±0.12Ac | 2.32±0.04Aa |
| 1.60 | 0.81±0.05Dd | 0.35±0.00Dd | 1.15±0.05Dd | 0.28±0.01Dd | 2.30±0.15Aa |
不同浓度海盐的绿竹叶片反射光谱曲线的整体变化趋势一致,均具有典型的反射光谱特征,420—500 nm蓝光谷、550 nm处绿光反射峰、600—680 nm红光谷、680—740 nm红边区的快速上升以及780—1000 nm的近红外高原平台。但是在不同浓度下反射光谱的反射率大小则存在差异,尤其是在绿光区 (525—605 nm)和近红外区(780—1000 nm)差异明显,1.6%浓度处理处理在绿光区(525—605 nm)明显高于其他处理,在556 nm和处达到最高值,相比较对照高出了30.8% (图 1)。

|
| 图 1 绿竹叶片反射光谱曲线和一阶导数曲线 Fig. 1 Reflectance spectra and first derivative of Bambusa oldhamii leaves |
由于光谱微分技术对光谱的噪声非常敏感,在实际分析光谱数据过程中,为了减少背景噪声的影响以提高生化参数的监测效果,常常需要对原始光谱数据进行微分变换,把实验测得光谱数据进行一阶求导 (图 1)。图中可以看出,在蓝光区505—545 nm和红光区680—750 nm出现最大峰值,黄光区560—640 nm出现最小峰值。红边是绿色植物光谱最明显的特征之一,它是指绿色植物反射光谱位于红光范围 (680—750 nm) 的光谱。不同浓度处理的绿竹叶片反射光谱值在红光区680—750 nm只有一个峰值,相比对照,随着海盐浓度的增加,λred先增大后减小,也就是说先向长波方向移动,后向短波方向移动,1.2%浓度处理下减小不显著,但是1.6%浓度处理,λred显著减小,Sred相比对照出现减小趋势 (表 4)。
| 参数 Parameter | 海盐浓度Sea salt concentration/% | ||||
| 0 (Ck) | 0.40 | 0.80 | 1.20 | 1.60 | |
| 小写字母不同表示0.05水平上的差异性 (P< 0.05) | |||||
| 蓝光波长 Blue wavelength(λblue) | 521.80±1.10a | 523.10±0.99a | 522.67±1.58a | 522.30±2.11a | 513.00±0.82b |
| 蓝光幅值Blue amplitude (Dλblue) | 0.48±0.03b | 0.50±0.05b | 0.47±0.06b | 0.31±0.06c | 0.58±0.05a |
| 红光面积 Blue area(Sblue) | 11.88±0.89b | 12.70±1.67b | 12.57±1.32b | 7.96±1.43c | 16.48±1.40a |
| 黄光波长 Yellow wavelength(λyellow) | 627.33±2.50b | 628.70±1.95a | 628.11±1.69a | 627.80±2.84ab | 626.67±0.52c |
| 黄光幅值 Yellow amplitude (Dλyellow) | -0.06±0.02b | -0.06±0.01b | -0.06±0.01b | -0.04±0.01a | -0.09±0.01c |
| 黄光面积 Yellow area(Syellow) | -13.10±1.92bc | -12.33±1.45b | -11.96±1.22b | -8.03±1.27a | -14.11±0.97c |
| 红光波长 Red wavelength (λred) | 698.50±1.64b | 701.50±0.11a | 699.67±0.87ab | 698.25±3.77b | 695.25±0.69c |
| 红光幅值Red amplitude(Dλred) | 1.32±0.07ab | 1.23±0.11b | 1.11±0.07c | 0.95±0.05d | 1.40±0.10a |
| 红光面积 Red area(Sred) | 44.66±2.74a | 25.03±2.42c | 38.93±3.87b | 40.64±0.97b | 39.92±2.74b |
光谱参数是绿色植物的光谱反射特征,是反映植物生长状况的最常用光谱变量。当海盐浓度为1.2%时,叶绿素参数rNDVI705和gNDVI显著下降 (P<0.05),与对照相比分别降低了27.3%和23.3%;当海盐浓度达到1.6%时,rNDVI705、mND705、gNDVI和mSR705呈极显著的降低 (P< 0.01),与对照相比分别下降了42.4%、43.9%、32.6%、21.5%。在1.2%海盐浓度处理下,类胡萝卜素参数CRI700和PRI显著降低 (P< 0.05),与对照相比分别降低了19.5%和43.9%;当海盐浓度增加到1.6%时,CRI550、CRI700和PRI相比较对照分别下降了47.2%、49.9%和58.5% (表 5)。
| 参数 Parameter | 海盐浓度Sea salt concentration/% | ||||
| 0 (Ck) | 0.40 | 0.80 | 1.20 | 1.60 | |
| 小写字母不同表示0.05水平上的差异性 (P< 0.05) | |||||
| 红边归一化指数 (rNDVI705) Red-edge normalized difference vegetation index | 0.33±0.12ab | 0.37±0.25a | 0.28±0.19b | 0.24±0.21b | 0.19±0.11c |
| 改良红边比值指数 (mND705) Modified red-edge normalized difference vegetation index | 0.41±0.2ab | 0.46±0.14a | 0.36±0.22b | 0.35±0.19b | 0.23±0.09c |
| 绿度归一化指数 (gNDVI) Greenness normalized index | 0.43±0.15a | 0.45±0.28a | 0.36±0.18b | 0.33±0.26b | 0.29±0.16c |
| 改良归一化差值指数 (mSR705) Modified red-edge ratio | 1.35±0.48ab | 1.43±0.65a | 1.20±0.65b | 1.19±0.59b | 1.06±0.54c |
| 类胡萝卜素反射指数1 (CRI550) Carotenoid reflectance index I | 6.72±1.56a | 6.91±2.45a | 6.82±2.69b | 5.89±3.10c | 3.55±2.46d |
| 类胡萝卜素反射指数2 (CRI700) Carotenoid reflectance index Ⅱ | 6.55±2.05a | 6.63±1.96a | 6.58±2.98ab | 5.27±3.45b | 3.28±2.58c |
| 光化学反射指数 (PRI) Photochemical reflectance index | 0.41±0.03a | 0.38± 0.01a | 0.32±0.07b | 0.23± 0.06b | 0.17± 0.06c |
相关分析表明 (表 6),绿竹叶片叶绿素a、叶绿素b、叶绿素a+b及类胡萝卜素含量与反射光谱参数都存在显著的相关性 (P < 0.05)。叶绿素a、叶绿素b和叶绿素a+b含量与rNDVI、mND、gNDVI、mSR705、CRI550、CRI700、PRI等呈极显著 (P < 0.01) 正相关,其中与CRI700相关系数最小,与mND的相关系数最大。类胡萝卜素含量与rNDVI、mND、gNDVI、mSR705等呈极显著 (P < 0.01) 正相关,与CRI550、CRI700、PRI等呈显著 (P < 0.01) 正相关,其中与mSR705相关系数最小,与mND的相关系数最大。
| 光谱参数 Spectral parameters | 叶绿素a Chlorophyll a | 叶绿素b Chlorophyll b | 叶绿素a+b Chlorophyll a+b | 类胡萝卜素 Carotinoid |
| * 表示差异显著 (P < 0.05),** 表示差异极显著 (P < 0.01) | ||||
| 红边归一化指数 (rNDVI705) Red-edge normalized difference vegetation index | 0.883* * | 0.911* * | 0.895* * | 0.785* * |
| 改良红边比值指数 (mND705) Modified red-edge normalized difference vegetation index | 0.901* * | 0.909* * | 0.917* * | 0.870* * |
| 绿度归一化指数 (gNDVI) Greenness normalized index | 0.877* * | 0.927* * | 0.874* * | 0.702* * |
| 改良归一化差值指数 (mSR705) Modified red-edge ratio | 0.856* * | 0.901* * | 0.868* * | 0.761* * |
| 类胡萝卜素反射指数1 (CRI550) Carotenoid reflectance index I | 0.778* * | 0.846* * | 0.795* * | 0.642* |
| 类胡萝卜素反射指数2 (CRI700) Carotenoid reflectance index Ⅱ | 0.736* * | 0.814* * | 0.755* * | 0.699* |
| 光化学反射指数 (PRI) Photochemical reflectance index | 0.818* * | 0.836* * | 0.805* * | 0.671* |
绿竹叶片经过暗适应后转入饱和脉冲光强照射,叶绿素荧光迅速上升,经过O、J、I、P各点后逐渐平稳。海盐浓度0.8%处理的O点和P点的荧光强度上升幅度较大,1.6%浓度处理出现了K点 (图 2A)。1.2%和1.6%处理的J点最高,然后骤然下降,达到I点时最低然后又上升。J点的相对可变荧光强度在1.2%和1.6%浓度处理明显下降,1.6%下降最明显之后随时间而上升 (图 2B)。在J、I点各处理间的相对可变荧光强度差值明显增加,其中1.2%和1.6%浓度处理差值增加最为明显。随着时间推移,处理间的相对可变荧光强度恢复到同一水平 (图 2C)。
2.5 不同浓度海盐对叶片叶绿素荧光动力学参数的影响基于生物膜能量流动理论[21]计算损伤后各时间点的荧光动力学参数,可以观察到不同海盐浓度处理后叶片叶绿素荧光动力学参数发生明显变化 (表 7)。
| 参数 Parameter | 海盐浓度Sea salt concentration/% | ||||
| 0(对照) | 0.4 | 0.8 | 1.2 | 1.6 | |
| 小写字母不同表示0.05水平上的差异性 (P< 0.05) | |||||
| 光系统Ⅱ最大量子产率(Fv/Fm) Maximum quantum yield of PSⅡ | 0.59±0.08b | 0.70±0.01a | 0.62±0.06ab | 0.59±0.02b | 0.29±0.04c |
| 电子传递的量子产额 (ΦEo) Quantum yield for electron transport | 0.21±0.05bc | 0.32±0.03a | 0.25±0.04b | 0.16±0.03c | 0.07±0.01d |
| 单位反应中心吸收的能量 (ABS/RC) Absorption flux per PSⅡ reaction center | 3.38±0.43b | 2.81±0.06c | 3.61±0.35b | 3.83±0.42b | 7.54±0.83a |
| 用于电子传递的能量 (ETo/RC) Electron transport per reaction center | 0.69±0.11b | 0.91±0.06a | 0.90±0.08a | 0.49±0.06c | 0.54±0.06c |
| 单位面积反应中心数目 (RC/CS) Density of reaction centers | 3.80±0.21a | 3.65±0.27a | 3.70±0.44a | 3.41±0.43a | 1.82±0.31b |
| 叶片性能指数 (PIABS) Performance index of absorption basis | 0.26±0.15b | 0.73±0.12a | 0.34±0.14b | 0.18±0.07bc | 0.02±0.01c |
|
|
| 图 2 不同海盐浓度对绿竹叶片荧光动力学曲线的变化 Fig. 2 Variation of chlorophyll fluorescence transients different concentration of sea salt stressed leaves of Bambusa oldhamii A: 不同海盐浓度对绿竹叶片O-J-I-P荧光诱导曲线 (3次重复的平均值); B: Fo(minimal fluorescence intensity) 与Fm(maximal fluorescence intensity) 间相对可变荧光强度 (relative variable fluorescence, Vt) 随时间的变化,Vt = (Ft -Fo)/(Fm-Fo),Ft (description fluorescence at time t) 表示t时的荧光强度,Fo表示暗适应后的最小荧光强度,Fm表示暗适应后的最大荧光强度; C: Vt与对照的差值△Vt,△Vt = Vt(处理) -Vt (对照);O、J、I、P各点含义详见李鹏民等[22] |
由表 7可以看出:随着盐胁迫强度的增加,ABS/RC呈现先下降后上升的趋势,在1.6%浓度处理处达到最大值,比对照上升了1.23 倍 (P< 0.05);ETo/RC呈现先上升后下降的趋势,相比较对照下降了21.7% (P< 0.05)。RC/CS随着浓度的升高呈现下降趋势,1.6%浓度处理比对照下降了52.1% (P< 0.05);ΦEo呈现先增加后下降的趋势,比对照下降了28.6% (P< 0.05)。Fv/Fm和PIABS呈现先升高后下降的趋势,相比对照,1.2%浓度处理下降,但不显著,1.6%浓度处理下降显著,分别下降了50.8%和92.3% (P< 0.05)(表 7)。
2.6 不同浓度海盐的光谱色素参数与PRI、Fv/Fm、ΦEo、PIABS的相关性由表 8可以看出,PRI与光谱色素参数在各个处理中具有相关性,尤其是与对照具有极显著性相关,0.4%浓度处理下的mNDVI705、mND705、gNDVI、mSR705、CRI550显著相关,0.8%浓度处理下的gNDVI、mSR705出现显著相关,CRI550和CRI700极显著性相关,1.2%浓度与1.6%浓度下的呈现显著相关性。
| 参数Parameters | mNDVI705 | mND705 | gNDVI | mSR705 | CRI550 | CRI700 | |
| *代表显著相关 (P< 0.05);**代表极显著相关 (P< 0.01); rNDVI705:红边归一化指数; mND705: 改良红边比值指数; gNDVI: 绿度归一化指数; mSR705: 改良归一化差值指数; CRI550: 类胡萝卜素反射指数1; CRI700: 类胡萝卜素反射指数2; PRI: 光化学反射指数; ΦEo,电子传递的量子产额; Fv/Fm,PSⅡ最大量子产率; PIABS,叶片性能指数; * indicates correlation significant at 0.05 level,** indicates correlation significant at 0.01 level. rNDVI705: red-edge normalized difference vegetation index;mND705: modified red-edge normalized difference vegetation index;gNDVI: greenness normalized index; mSR705: modified red-edge ratio; CRI550: carotenoid reflectance index I; CRI700: carotenoid reflectance index Ⅱ; PRI: photochemical reflectance index; ΦEo,quantum yield for electron transport; Fv/Fm,maximum quantum yield of PSⅡ; PIABS,performance index of leaf | |||||||
| 0 (Ck) | PRI | 0.934* * | 0.925* * | 0.98* * | 0.94* * | 0.972* * | 0.902* * |
| Fv/Fm | 0.776* | 0.779* | 0.851* * | 0.771* | 0.632 | 0.996* * | |
| ΦEo | 0.668 | 0.692 | 0.776* | 0.681 | 0.526 | 0.976* * | |
| PIABS | 0.802* | 0.805* | 0.872* * | 0.796* | 0.664 | 0.999* * | |
| 0.40% | PRI | 0.767* | 0.739* | 0.867* * | 0.809* | 0.864* * | 0.696 |
| Fv/Fm | 0.264 | 0.256 | 0.331 | 0.298 | 0.587 | 0.682 | |
| ΦEo | 0.874* * | 0.878* * | 0.838* | 0.856* * | 0.926* * | 0.873* * | |
| PIABS | 0.900* * | 0.903* * | 0.867* * | 0.883* * | 0.904* * | 0.844 | |
| 0.80% | PRI | -0.52 | -0.36 | -0.719* | -0.682* | -0.910* * | -0.905* * |
| Fv/Fm | 0.834* | 0.822* | 0.887* * | 0.824* | 0.884* * | 0.768* | |
| ΦEo | 0.837* | 0.825* | 0.902* * | 0.824* | 0.911* * | 0.801* | |
| PIABS | 0.838* | 0.846* | 0.687 | 0.654 | 0.711* | 0.313 | |
| 1.20% | PRI | 0.724* | 0.734* | 0.842* | 0.716* | 0.135 | 0.148 |
| Fv/Fm | 0.943* * | 0.915* * | 0.925* * | 0.984* * | 0.411 | 0.176 | |
| ΦEo | 0.994* * | 0.998* * | 0.998* * | 0.966* * | 0.763* | 0.528 | |
| PIABS | 0.979* * | 0.989* * | 0.999* * | 0.981* * | 0.718* | 0.526 | |
| 1.60% | PRI | 0.742* | 0.727* | 0.595 | 0.758* | 0.446 | 0.323 |
| Fv/Fm | 0.986* * | 0.983* * | 0.992* * | 0.996* * | 0.999* * | 0.999* * | |
| ΦEo | 0.968* * | 0.964* * | 0.978* * | 0.985* * | 0.992* * | 0.995* * | |
| PIABS | 0.914* * | 0.908* * | 0.930* * | 0.943* * | 0.957* * | 0.965* * | |
由表 8可以看出,在对照水平,Fv/Fm与mNDVI705、mND705、gNDVI、mSR705、CRI550具有相关性,0.4%浓度下无相关性,其他3个处理都具有良好的相关性;在对照水平,ΦEo与gNDVI和CRI700显著相关,在其他处理水平,与mNDVI705、mND705、gNDVI、mSR705、CRI550都具有显著相关性;在对照、0.4%、1.2%和1.6%浓度水平上,PIABS与叶绿素光谱参数具有显著相关性,0.8%浓度处理水平与mNDVI705、mND705、CRI550显著相关。
3 讨论叶绿素是植物叶片中吸收和传递光能的主要色素分子,直接影响光合作用的效能与水平,是植物活力的重要体现;同时,叶绿素含量水平也是反映植物营养状况和生长发育进程的重要指标[23, 24]。有研究表明,盐胁迫使黄瓜 (Cucumis sativus) 叶绿体大部分与细胞壁脱离,基粒片层数减少,叶片光合色素含量下降,引起植株光合能力降低[25]。随着NaCl处理浓度的加大,白柠条 (Caragana korshinskii) 和骆驼刺 (Alhagi sparsifolia) 的叶绿素a含量呈先升高后降低趋势[26]。NaCl胁迫后,黄瓜叶片叶绿素a、叶绿素b 和叶绿素总含量均显著降低[26]。本研究表明:随着海盐浓度的增大,光合色素呈现先升高后降低的趋势 (表 3),说明在低盐胁迫下,绿竹可能通过提高光合色素合成速度对低盐的适应[27];高盐胁迫下,可能导致绿竹叶片光合色素合成速度降低,或降解速度加快 (表 3)。
光谱分析是一种快速、准确判断物体所含成分及含成分多少的有效方法。在植物生理监测过程中,反射光谱因其无损、快速、高效的优越性,成为获取农田生物环境信息的重要手段,在精确农业发展中发挥着重要作用。在可见光区域影响光谱反射的主导因子是叶绿素和类胡萝卜素[17],叶绿素在可见光区有很强的光吸收能力。有研究表明,盐处理明显增加了黄瓜叶片在可见光区的光谱反射率[3]。Peuelas 等[28]在大麦 (Horduem vulgare) 上也做过类似研究,结果显示近红外反射随盐浓度升高而降低,而可见光反射随盐浓度升高而增加。随着海盐浓度的升高,光谱参数降低 (表 5),色素含量降低 (表 3) 说明两者具有一定的相关性 (表 6),这与张丽平[3]和Peňuelas[28]研究结果相似。
叶绿素荧光具有反应内在性的特点,是研究植物光合生理与逆境胁迫最好的无损检测技术[21]。张蕾等[29]对4种珍稀观赏竹光合作用特性研究认为Fv/Fm都随光强增大而逐步下降。张秋英等[30]研究了水分胁迫对小麦 (Triticum aestivuml) 旗叶叶绿素a荧光动力学参数的影响,发现旗叶的T l/2值减少,旗叶Fv/Fm和潜在活性 (potential activity, Fv/Fo) 降低。应叶青等[31]研究认为毛竹 (Phyllostachys edulis) 幼苗叶片初始荧光表现出随干旱胁迫程度的加剧,不断增加的趋势,而Fm、ΦPSⅡ、Fv/Fm呈逐渐降低的趋势。当盐胁迫浓度增加到1.6%时,绿竹叶片叶绿素荧光动力学曲线出现明显的K点 (图 2),表明绿竹叶片光合作用放氧系统受到严重的抑制,以及QA供体侧受损[21],导致ETo/RC、ΦEo、Fv/Fm和PIABS降低,反应中心降解或失活[7]。PRI被认为是与叶绿素含量高度相关[32],Rahimzadeh-Bajgiran等[33]认为在植物叶片生长的各个阶段,PRI以及叶绿素荧光参数和植物色素指数在短期密切相关。在海盐胁迫下,PRI、Fv/Fm、ΦEo和PIABS与光谱色素参数具有良好的相关性 (表 8)。
综上所述,高盐胁迫 (1.6%) 条件下,抑制绿竹叶片光合色素的合成,降低光能的吸收,导致叶片发生光抑制,PSⅡ的受体侧受到伤害,PSⅡ反应中心降解或失活,直接抑制绿竹的生长发育。低盐胁迫下,绿竹通过提高叶片光合色素含量和光能的吸收,增加Fv/Fm、ΦEo、ETo/RC、RC/CS和PIABS,提高绿竹的耐盐能力。从本研究的结果来看,当盐胁迫增加到1.2%时,绿竹叶片保持较高的色素含量和光能吸收能力,以及Fv/Fm、ΦEo、ETo/RC、RC/CS和PIABS,说明绿竹具有一定的耐盐性。
| [1] | Hu H B, Zhang J C, Lu X Z. Study on environmental benefits of coastal windbreak system in China. World Forestry Research, 2001, 14(5): 37-43. |
| [2] | Shi Z, Shi S Q, Zhang Z X, Gao R F, Xiao W F. Effects of drought and excess salt stress on characteristics of chlorophyll fluorescence of leaves in Haloxylon ammodendron. Journal of Beijing Forestry University, 2012, 34(3): 20-25. |
| [3] | Zhang L P, Zhang G Y, Shi Q H, Ding F, Wang X F, Feng J P. Effects of NaCl stress on damage and reflectance characteristics of cucumber cultivars with different salt-tolerance. Plant Nutrition and Fertilizer Science, 2008, 14(4): 761-765. |
| [4] | Eklundh L, Johansson T, Solberg S. Mapping insect defoliation in Scots pine with MODIS time-series data. Remote Sensing of Environment, 2009, 113(7): 1566-1573. |
| [5] | Fang X W, Turner N C, Li F M, Guo X S. Caragana korshinskii seedlings maintain positive photosynthesis during short-term severe drought stress. Photosynthetica, 2011, 49(4): 603-609. |
| [6] | Li G, Wan S W, Zhou J, Yang Z Y, Qin P. Leaf chlorophyll fluorescence, hyperspectral reflectance, pigments content, malondialdehyde and proline accumulation responses of castor bean (Ricinus communis L.) seedlings to salt stress levels. Industrial Crops and Products, 2010, 31(1): 13-19. |
| [7] | Liu B X, Wang Z G, Yang M S, Liang H Y. Effects of simulated salt stress on seed germination, seedling emergence and growth of Ulmus pumila . Acta Prataculturae Sinica, 2012, 21(5): 39-46. |
| [8] | Mehta P, Jajoo A, Mathur S, Bharti S. Chlorophyll a fluorescence study revealing effects of high salt stress on Photosystem Ⅱ in wheat leaves. Plant Physiology and Biochemistry, 2010, 48(1): 16-20. |
| [9] | Zheng R M, Zheng Y S, Zhang M, Li Q R, Chen X D, Chen L G. Study on Impact of Photosynthesis and Chlorophyll of Dendrocalamopsis oldhami Grown in the Coastal Areas Under Salt Stress. Journal of Bamboo Research, 2002, 21(4): 76-80. |
| [10] | Zheng Y S, Zheng R M, Yao Q D. Effects of salt stress on physiology and biochemistry in the leaves of Dendrocalamus oldhami on the coast. Journal of Fujian Agriculture and Forestry University, 2003, 32(4): 478-481. |
| [11] | Gao Z M, Liu Q, Mu S H, Li X P, Hu T. Cloning and primary functional analysis of a zinc finger protein gene of BoBZF from Bambusa oldhamii. Scientia Silvae Sinicae, 2012, 48(10): 49-54. |
| [12] | Zhu Y, Luo C G. Analysis of nutrient components in Dendrocalamopsis oldhami bamboo shoot. Nonwood Forest Research, 2012, 30(3): 103-105. |
| [13] | Pei H Y, Fang W, Lin X C, Gui R Y, Huang L C. Micropropagation of Bambusa oldhamii f. variegata by tissue culture. Journal of Zhejiang Forestry College, 2010, 27(1): 149-154. |
| [14] | Shao J F, Gui R Y, Ji H B, Li G D, Fang W. A preliminary study on establishment of hydroponic culture system for Phyllostachys pubescens seedlings. Journal of Zhejiang A & F University, 2011, 28(1): 86-94. |
| [15] | Xiao J, Xiao Y K, Liu C Q, Zhao Z Q, He M Y, Liang C S. Boron isotopic fractionation during incorporation of boron into Mg(OH)2. Chinese Science Bulletin, 2009, 54(17): 2363-2371. |
| [16] | Lichtenthaler H K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods in Enzymology, 1987, 148: 350-382. |
| [17] | Sims D A, Gamon J A. Relationships between leaf pigment content and spectral reflectance across a wide range of species leaf structures and developmental stages. Remote Sensing of Environment, 2002, 81(2/3): 337-354. |
| [18] | Gitelson A A, Merzlyak M N. Signature analysis of leaf reflectance spectra: algorithm development for remote sensing of chlorophyll. Journal of Plant Physiology, 1996, 148(3): 494-500. |
| [19] | Gitelson A A, Zur Y, Chivkunova O B, Merzlyak M N. Assessing carotenoid content in plant leaves with reflectance Spectroscopy. Photochemistry and Photobiology, 2002, 75(3): 272-281. |
| [20] | Gamon J A, Penuelas J, Field C B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sensing of Environment, 1992, 41(1): 35-44. |
| [21] | Strasser R J, Srivastava A, Tsimilli-Michael M. 2000. The fluorescence transient as a tool to characterize and screen photosynthetic samples//Yunus M, Pathre U, Mohanty P, eds. Probing Photosynthesis: Mechanisms, Regulation and Adaptation. London: Taylor and Francis, 445-483. |
| [22] | Li P M, Gao H Y, Strasser R J. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study. Journal of Plant Physiology and Molecular Biology, 2005, 31(6): 559-566. |
| [23] | Gitelson A A, Gitzt Y, Merzlyak M N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. Journal of Plant Physiology, 2003, 160(3): 271-282. |
| [24] | Moorthy I, Miller J R, Noland T L. Estimating chlorophyll concentration in conifer needles with hyperspectral data: An assessment at the needle and canopy level. Remote Sensing of Environment, 2008, 112(6): 2824-2838. |
| [25] | Tong H, Zhang Z X, Li B, Wang J W, Guo S R. Effects of Iso-osmotic Ca(NO3)2 and NaCl stress on chloroplast Ultra-structure and photosynthesis in cucumber leaves. China Vegetables, 2012, (18): 160-165. |
| [26] | Han Z X, Li L, Xu X W, Lü X F, Yue H X, Bian Z, Li L Z. Effect of NaCl on physiological features of 4 legume seedlings in desert areas of Xinjiang, China. Chinese Bulletin of Botany, 2012, 47(5): 491-499. |
| [27] | Matoh T, Murata S. Sodium stimulates growth of Panicum coloratlzm through enhanced photosynthesis. Plant Physiology, 1990, 92(4): 1169-1173. |
| [28] | Peñuelas J, Isla R, Fillela I, Araus J L. Visible and near-infrared reflectance assessment of salinity effects on barley. Crop Science, 1997, 37(1): 198-202. |
| [29] | Zhang L, Jin S H, Fang W. Characteristics of photosynthesis in four rare ornamental bamboo species. Journal of Zhejiang University, 2009, 35(2): 165-172. |
| [30] | Zhang Q Y, Li F D, Liu M Y, Dong B D. Effect of water stress on the photosynthesis of flag leaf of winter wheat. Agricultural Research in the Arid Areas, 2002, 20(3): 80-84. |
| [31] | Ying Y Q, Guo J, Wei J F, Zou Y Q, Hu D C, Fang W. Photosynthetic and chlorophyll fluorescent responses of Phyllostachys pubescens seedlings to water deficiency stress. Journal of Beijing Forestry University, 2009, 31(6): 128-133. |
| [32] | Garbulsky M F, Peñuelas J, Gamon J, Inoue Y, Filella I. The photochemical reflectance index (PRI) and the remote sensing of leaf, canopy and ecosystem radiation use efficiencies-a review and meta-analysis. Remote Sensing of Environment, 2011, 115(2): 281-297. |
| [33] | Rahimzadeh-Bajgiran P, Munehiro M, Omasa K. Relationships between the photochemical reflectance index (PRI) and chlorophyll fluorescence parameters and plant pigment indices at different leaf growth stages. Photosynthesis Research, 2012, 113(1/3): 261-271. |
| [1] | 胡海波, 张金池, 鲁小珍. 我国沿海防护林体系环境效应的研究. 世界林业研究, 2001, 14(5): 37-43. |
| [2] | 施征, 史胜青, 张志翔, 高荣孚, 肖文发. 干旱与高盐对梭梭叶绿素荧光特性的影响. 北京林业大学学报, 2012, 34(3): 20-25. |
| [3] | 张丽平, 张格英, 史庆华, 丁飞, 王秀峰, 冯建鹏. 氯化钠胁迫对不同耐盐黄瓜品种的损伤效应及反射光谱特性的影响. 植物营养与肥料学报, 2008, 14(4): 761-765. |
| [7] | 刘炳响, 王志刚, 杨敏生, 梁海永. 模拟盐胁迫对白榆种子发芽、出苗及幼苗生长的影响. 草业学报, 2012, 21(5): 39-46. |
| [9] | 郑容妹, 郑郁善, 张梅, 李庆荣, 陈羡德, 陈礼光. 盐分胁迫对沿海绿竹光合作用及叶绿素的影响. 竹子研究汇刊, 2002, 21(4): 76-80. |
| [10] | 郑郁善, 郑容妹, 姚庆端. 盐分胁迫对沿海沙地绿竹生理生化的效应. 福建农林大学学报, 2003, 32(4): 478-481. |
| [11] | 高志民, 刘青, 牟少华, 李雪平, 胡陶. 绿竹锌指蛋白基因BoBZF克隆及功能初步分析. 林业科学, 2012, 48(10): 49-54. |
| [12] | 朱勇, 罗朝光. 绿竹笋营养成分的测定与分析. 经济林研究, 2012, 30(3): 103-105. |
| [13] | 裴海燕, 方伟, 林新春, 桂仁意, 黄丽春. 花叶花秆绿竹的试管快繁研究. 浙江林学院学报, 2010, 27(1): 149-154. |
| [14] | 邵继峰, 桂仁意, 季海宝, 李国栋, 方伟. 毛竹实生苗水培体系初步建立. 浙江农林大学学报, 2011, 28(1): 86-94. |
| [15] | 肖军, 肖应凯, 刘丛强, 赵志琦, 贺茂勇, 梁重山. 硼掺入 Mg(OH)2过程中的硼同位素分馏. 科学通报, 2009, 54(16): 2363-2371. |
| [22] | 李鹏民, 高辉远, Strasser R J. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用. 植物生理与分子生物学学报, 2005, 31(6): 559-566. |
| [25] | 童辉, 张振兴, 李斌, 王军伟, 郭世荣. 等渗Ca(NO3)2和NaCl胁迫对黄瓜叶片叶绿体超微结构及光合特性的影响. 中国蔬菜, 2012, (18): 160-165. |
| [26] | 韩张雄, 李利, 徐新文, 吕湘芳, 岳红霞, 边振, 李力争. NaCl对4种荒漠豆科植物幼苗生理参数的影响. 植物学报, 2012, 47(5): 491-499. |
| [29] | 张蕾, 金松恒, 方伟. 4种珍稀观赏竹光合作用特性研究. 浙江大学学报, 2009, 35(2): 165-172. |
| [30] | 张秋英, 李发东, 刘孟雨, 董宝娣. 水分胁迫对小麦旗叶叶绿素a荧光参数和光合速率的影响. 干旱地区农业研究, 2002, 20(3): 80-84. |
| [31] | 应叶青, 郭璟, 魏建芬, 邹奕巧, 胡冬春, 方伟. 水分胁迫下毛竹幼苗光合及叶绿素荧光特性的响应. 北京林业大学学报, 2009, 31(6): 128-133. |