 2021, Vol. 41
2021, Vol. 41文章信息
- 孙艺, 李宏俊, 顾炎斌, 樊景凤, 郭皓
- SUN Yi, LI Hongjun, GU Yanbin, FAN Jingfeng, GUO Hao
- 利用分类充分性对辽河口大型底栖动物监测的优化研究
- Optimization of Macrobenthos monitoring strategy using taxonomic sufficiency in the Liaohe Estuary
- 生态学报. 2021, 41(4): 1645-1655
- Acta Ecologica Sinica. 2021, 41(4): 1645-1655
- http://dx.doi.org/10.5846/stxb202001230169
-
文章历史
- 收稿日期: 2020-01-23
- 网络出版日期: 2020-12-24

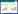




生态评价依赖于准确的生物种类鉴定技术, 在传统的环境影响生物群落研究中, 物种种级虽然是信息量最大的数据集水平, 但是存在工作量大、专业成本高、耗费时间长、鉴定错误率高等问题[1-3], 因此一些学者提出利用高分类阶元替代种级水平开展生态评价的想法。1985年, Ellis在污染评价研究中提出分类充分性(Taxonomic sufficiency, TS)概念[4], 指在任何一个项目有机体被鉴定到满足准确指示有机体生物学需要的水平(如种、属、科等), 同时又不会在评估环境质量方面丧失统计学意义。众多研究表明, 利用科级或属级的物种分类阶元代替种级水平开展多样性分析、群落组成评估和环境压力响应研究, 具有相同或类似的结果[5-8]。在环境压力响应研究中, 部分研究结果表明, 科级或属级群落对环境因子的响应结果与种级相似, 如在北大西洋石油类污染物对底栖动物群落的影响研究中, 科级水平的合并减少了单一优势种的影响, 相对于属级能清晰反应群落结构与污染因子之间的关系[9];然而也有学者不同意使用分类充分性方法, 强调高分类阶元会丢失种级水平的信息, 例如同科或同属的大型底栖动物耐污能力差异较大, 种级水平的鉴定才能全面检测到群落结构对环境变化的响应[10-12]。
近年来, 人类活动导致的生物多样性衰退问题已引起广泛关注, 世界上大多数地方没有资源和时间开展具体细致的分类调查[13], 生物多样性快捷评价成为反映生物多样性状况和变化趋势的有效工具[14], TS在海洋生物多样性快捷评价研究中也得到广泛应用[15]。波斯湾潮间带大型底栖动物α多样性研究结果表明目级、科级、属级与种级的丰富度指数显著相关[16];在厦门湾大型底栖动物分类学多样性研究中表明, 属级、科级与种级传统多样性和分类学多样性指数显著相关, 种级、属级和科级的群落组成一致性强, 属级较种级丢失的信息少于科级[3];在亚得里亚海域2个采样区域大型底栖生物β多样性研究结果表明, TS适用于物种多样性较低和物种多样性较高2种采样区域的多样性分析, 在无法准确鉴定物种情况下, 科级水平代表性最佳[17];也有研究者指出, 在区域范围较广的海域, 多毛类动物科级水平相对于种级丢失的信息最少, 适用性最强[18]。
辽河口位于辽河下游入海口, 辽河口湿地是世界第二大、中国最大的滨海芦苇湿地, 该海域沿岸辽河、大辽河、大凌河和小凌河等多条河流入海, 不断输送各种营养物质。辽河口是我国重要的海水养殖区, 养殖排污加剧区域N、P营养盐污染风险[19]。辽河口水深较浅, 水体交换能力差, 容易加剧污染物富集, 根据《2012年中国海洋环境状况公报》显示, 辽河口主要污染要素是无机氮、活性磷酸盐和石油类。前人已经开展过辽东湾大型底栖动物群落生态学研究, 结果表明辽东湾六股河口邻近海域冬季大型底栖动物丰度和生物量分布呈现远离河口增高趋势, 河口低盐、陆源污染、航道疏浚限制了大型底栖动物的生长和繁衍[20];辽东湾夏季潮间带大型底栖动物受到不同程度的干扰, 主要受底质、生境等人为因素干扰[21], 但目前关于辽河口大型底栖动物季节演替研究开展较少。本研究探讨辽河口春、夏、秋3个季节基于不同分类阶元分析大型底栖动物群落组成的相关性及其响应环境压力变化, 以期为辽河口大型底栖动物监测评价提供参考。
1 材料与方法 1.1 调查方法分别于2013年8月、10月和2014年5月在辽河口开展夏、秋、春3个航次的大型底栖动物调查, 设置25个站位开展定点采样(图 1)。使用0.05 m2的抓斗式采泥器取样3次, 并用孔径为0.5 mm的网筛分选大型底栖动物, 5%甲醛溶液固定保存, 在实验室进行形态鉴定、计数和称重, 参照《海洋监测规范》(GB/T 12763.6—2007)。每个站位采集3份水体和沉积物样本低温保存带回实验室测定, 分析的环境因子包括沉积环境因子:叶绿素a(Chla)和沉积物粒度, 海水环境因子:盐度(Sal)、水深(Depth)、悬浮物(SS)、总氮(TN)、总磷(TP)、石油类(Oil)和溶解氧(DO)。其中沉积物Chla采用湿样法[22], 沉积物粒度采用激光法分析, 参照《海洋监测规范》(GB/T12763.8—2007)。悬浮物(重量法)、总氮(过硫酸钾氧化法)、总磷(钼酸铵分光光度法)、石油类(荧光分光光度法)参照《海洋监测技术规程》(GB 17378.4—2007), 盐度、水深、溶解氧等环境因子使用YSI多参数便携式水质分析仪现场测定。

|
| 图 1 辽河口大型底栖动物样品采集点 Fig. 1 Sampling stations of macrobenthos in the Liaohe Estuary |
大型底栖动物物种名录通过《中国海洋生物种类与分布》查询和校对。本文将大型底栖动物鉴定至种级, 然后分别提高至属级、科级和目级水平, 计算不同分类阶元水平聚集率(φ), 不同分类阶元水平选用定量数据(丰度)和定性数据(有/无), 比较分析不同的处理对结果的影响, 聚集率的计算方法是高级分类数/种数[23], 不同分类水平及说明参照表 1。
| 变量Variable | 定义Definition | 变量Variable | 定义Definition | |
| 种Quan | 种级定量水平(丰度数据) | 种Qual | 种级定性水平(有/无数据) | |
| 属Quan | 属级定量水平(丰度数据) | 属Qual | 属级定性水平(有/无数据) | |
| 科Quan | 科级定量水平(丰度数据) | 科Qual | 科级定性水平(有/无数据) | |
| 目Quan | 目级定量水平(丰度数据) | 目Qual | 目级定性水平(有/无数据) |
为探讨不同分类阶元群落组成一致性, 首先对各个分类阶元的丰度数据进行平方根转换和定性数据转换[24], 计算定量种级与更高分类阶元之间Bray-Curtis距离相似矩阵、定性种级与更高分类阶元之间Sørensen距离相似矩阵, 并使用RELATE程序确定种级与更高分类阶元间相似矩阵间的Spearman等级相关分析(定量数据和定性数据), 其中相关系数变化值Rho从0—1, 其中0代表两个分类阶元之间没有相关性, 1代表两个分类阶元间相关性极高。为了更直观的体现不同分类阶元间的相关性排布模式, 针对上述两种矩阵分别做Second-stage非度量多维尺度分析(NMDS)二维排序图, 对比不同分类阶元群落组成的一致性。根据NMDS排序图排序结果筛选需要的分类阶元进行Second-stage Cluster聚类树分析[25], 以确定更高分类阶元相比种级水平丢失信息的多少。
采用基于距离线性模型(DistLM)分析群落组成与环境因子响应结果, 群落数据使用定量与定性不同分类阶元数据Bray-Curtis距离、Sørensen距离相似矩阵, 环境因子进行ln(x+1)转换, 然后进行前向选择选出最佳环境因子并对DistLM模型进行R2校正(Adj. R2), 最后用Mantel test检验最佳环境因子与群落之间的相关性。
使用软件PRIMER 7.0[24]对物种定量数据转换、群落组成分析和DistLM模型分析, 使用R语言做Mental test、折线图和NMDS排序图可视化。
2 结果 2.1 辽河口大型底栖动物物种组成辽河口3个航次共采集到大型底栖动物105种(附录1), 环节动物40种为主要门类并且物种数最多, 占总数的38.10%;节肢动物30种, 占总数的28.57%;软体动物16种, 占总数15.24%;棘皮动物7种, 占总数6.67%;其余动物占总数11.43%。春季航次共采集大型底栖动物57种, 夏季航次62种, 秋季航次57种。春、夏、秋3个航次属种聚集率(表 2)分别为0.82、0.94和0.88, 科种聚集率分别为0.77、0.76和0.81, 目种聚集率0.37、0.35、0.46。
| 环节动物门Annelida | ||||
| 全刺沙蚕 | Nectoneanthes oxypoda | 乳突半突虫 | Phyllodoce papillosa | |
| 日本刺沙蚕 | Neanthes japonica | 渤海格鳞虫 | Gattyana pohaiensis | |
| 多齿围沙蚕 | Perinereis nuntia | 孟加拉还扇虫 | Pherusa cf.bengalensis | |
| 寡鳃齿吻沙蚕 | Nephtys oligobranchia | 梳鳃虫 | Terebellides stroemii | |
| 囊叶齿吻沙蚕 | Nephtys caeca | 羽鳃柿虫 | Schistocomus hiltoni | |
| 中华内卷齿沙蚕 | Aglaophamus sinensis | 那不勒斯膜帽虫 | Lagis neapolitana | |
| 中锐吻沙蚕 | Glycera rouxii | 拟蛰虫 | Amaeana trilobata | |
| 长吻沙蚕 | Glycera chirori | 不倒翁虫 | Sternaspis scutata | |
| 双小健足虫 | Micropodarke dubia | 岩虫 | Marphysa sanguinea | |
| 狭细蛇潜虫 | Ophiodromus angustifrons | 长叶索沙蚕 | Lumbrineris longiforlia | |
| 刚鳃虫 | Chaetozone setosa | 智利巢沙蚕 | Diopatra chilienis | |
| 富氏钙栅虫 | Dodecaceria fewkesi | 竹节虫 | Phasmatodea phasma | |
| 锥稚虫 | Aonides oxycephala | 寡节甘吻沙蚕 | Glycinde gurjanovae | |
| 马丁海稚虫 | Spio martinensis | 日本角吻沙蚕 | Goniada japonica | |
| 巴适钩毛虫 | Sigambra bassi | 小头虫 | Capitella capitata | |
| 背蚓虫 | Notomastus latericeus | 曲强真节虫 | Euclymene lombricoides | |
| 海蛹科 | Opheliidae | 真节虫科 | Euclymeninae | |
| 拟特须虫 | Paralacydonia paradoxa | 丝异须虫 | Heteromastus filiformis | |
| 付柿虫 | Paramphicteis angustifolia | 尖叶长手沙蚕 | Magelona cincta | |
| 中华半突虫 | Phyllodoce chinensis | 日本长手沙蚕 | Magelona japonica | |
| 节肢动物门Arthropoda | ||||
| 短角双眼钩虾 | Ampelisca brevicornis | 东北蝲蛄 | Cambaroides dauricus | |
| 日本沙钩虾 | Byblis japonicus | 河螺赢蜚 | Corophium acherusicum | |
| 博氏双眼钩虾 | Ampelisca bocki | 日本关公蟹 | Dorippe japonica | |
| 中型三强蟹 | Tritodynamia intermedia | 狭颚新绒螯蟹 | Neoeriocheir leptognathus | |
| 霍氏三强蟹 | Tritodynamia horvathi | 脊尾白虾 | Exopalaemon carinicauda | |
| 短齿大眼蟹 | Macrophthalmus brevis | 日本拟钩虾属 | Gammaropsis sp. | |
| 小头弹钩虾 | Orchomene breviceps | 大螯蜚属 | Grandidierell sp. | |
| 日本大螯蜚 | Grandidierella japonica | 寄居蟹属 | Pagurus sp. | |
| 平掌拟钩虾 | Gammaropsis laevipalmata | 豆形拳蟹 | Philyra pisum | |
| 中国毛虾 | Acetes chinensis | 中华豆蟹 | Pinnotherinae sinensis | |
| 毛盲蟹 | Typhlocarcinus villosus | 鹰爪虾 | Trachysalambria curvirostris | |
| 颗粒拟关公蟹 | Paradorippe granulata | 日本拟背尾水虱 | Paranthura japonica | |
| 细螯虾 | Leptonchela gracilis | 三叶针尾涟虫 | Diastylis tricincta | |
| 日本游泳水虱 | Natatolana japonensis | 口虾蛄 | Oratosquilla oratoria | |
| 伊予双眼钩虾 | Ampelisca iyoensis | 锯齿利尔钩虾 | Liljeborgia serrata | |
| 软体动物门Mollusca | ||||
| 饼干镜蛤 | Dosinia biscocta | 泥螺 | Bullacta ecarata | |
| 文蛤 | Meretrix meretrix | 毛蚶 | Scapharca kagoshimensis | |
| 小莢蛏 | Siliqua minima | 中国蛤蜊 | Mactra chinensis | |
| 小刀蛏 | Cultellus attenuatus | 扁玉螺 | Glossaulax didyma | |
| 纵肋织纹螺 | Nassarius variciferus | 光滑河篮蛤 | Potamocorbula laevis | |
| 红带织纹螺 | Nassarius succinctus | 彩虹明樱蛤 | Moerella iridescens | |
| 圆筒原盒螺 | Eocylichna braunsi | 白带三角口螺 | Trigonaphera bocageana | |
| 脆壳肌蛤 | Musculus perfragilis | 焦河篮蛤 | Potamocorbula ustulata | |
| 棘皮动物门Echinodermata | ||||
| 细雕刻肋海胆 | Temnopleurus toreumatica | 倍棘尾蛇属 | Amphioplus sp. | |
| 哈氏刻肋海胆 | Temnopleurus hardwickii | 棘刺锚参 | Protankyra bidentata | |
| 浅水萨氏真蛇尾 | Ophiura sarsii | 马氏刺蛇尾 | Ophiothrix marenzelleri | |
| 金氏真蛇尾 | Ophiura kinbergi | |||
| 其他门类Others | ||||
| 小头副孔鰕虎 | Paratrypauchen microcephalus | 苔藓动物门 | Bryozoa | |
| 孔鰕虎 | Trypauchen vagina | 纵条肌海葵 | Haliplanella luciae | |
| 斑尾刺鰕虎 | Synechogobius ommaturus | 短吻螠 | Listriolobus brevirostris | |
| 红狼牙鰕虎 | Odontamblyopus rubicundus | 纽形动物门 | Nemertea | |
| 哈氏仙人掌海鳃 | Cavernularia habereri | 涡虫纲 | Turbellaria | |
| 黄侧花海葵 | Anthopleura xanthogrammica | 星虫动物门 | Sipuncula | |
| 季节 Season |
目级 Order |
科级 Family |
属级 Genes |
种级 Species |
优势种(≥2%) Dominant Species |
| 春季Spring | 21 | 44 | 47 | 57 | 博氏双眼钩虾Ampelisca bocki |
| 聚集率φ | 0.37 | 0.77 | 0.82 | 河螺赢蜚Corophium acherusicum | |
| Aggregation ratio | 日本游泳水虱Natatolana japonensis | ||||
| 饼干镜蛤Dosinia biscocta | |||||
| 夏季Summer | 22 | 47 | 58 | 62 | 日本拟背尾水虱Paranthura japonica |
| 聚集率φ | 0.35 | 0.76 | 0.94 | 短角双眼钩虾Ampelisca brevicornis | |
| Aggregation ratio | 三叶针尾涟虫Diastylis tricincta | ||||
| 巴适钩毛虫Sigambra bassi | |||||
| 日本角吻沙蚕Goniada japonica | |||||
| 富氏钙栅虫Dodecaceria fewkesi | |||||
| 不倒翁虫Sternaspis scutata | |||||
| 锥稚虫Aonides oxycephala | |||||
| 泥螺Bullacta ecarata | |||||
| 秋季Autumn | 26 | 46 | 50 | 57 | 河螺赢蜚Corophium acherusicum |
| 聚集率φ | 0.46 | 0.81 | 0.88 | 羽鳃柿虫Schistocomus hiltoni | |
| Aggregation ratio | 梳鳃虫Terebellides stroemii | ||||
| 饼干镜蛤Dosinia biscocta | |||||
| 彩虹明樱蛤Moerella iridescens | |||||
| 扁玉螺Glossaulax didyma |
3个季节中群落组成分析表明, 大型底栖动物群落定量与定性数据Bray-Curtis相似矩阵和Sørensen距离相似矩阵的Spearman等级一致性的分析表明(图 2), 种级与更高分类阶元相似矩阵间都具有显著相关性(P < 0.001), 但相似矩阵之间的相关性系数随着分类阶元的增加而降低, 种级与属级矩阵之间的相关性极高(Rho>0.95), 科级与种级矩阵间的相关性也很强(Rho>0.88)但略低于属级。

|
| 图 2 3个季节不同分类阶元相似矩阵间Spearman相关性变化趋势 Fig. 2 Spearman correlation coefficient trends between different taxonomic resolution similarity matrices among three seasons S: Species, G: Genus, F: Family, O: Order |
Second-stage NMDS排序图结果如图 3所示, 图中每一个符号代表一个相似矩阵, 符号间的距离表示他们之间的相似度。3个季节定量和定性数据结果显示, 属级与种级相似性最高, 与目级的相似性最低, 选用属级、科级定量数据和种级定性数据进行Second-stage Cluster聚类树分析。根据Cluster聚类树(图 4)可知, 3个季节中定量与定性种级之间丢失信息约2%, 在不同分类阶元定量数据方面, 与种级相比, 属级丢失信息少于3%, 科级丢失的信息虽然多于属级, 但是丢失的信息也较少(< 6%), 这可能与辽河口大型底栖动物物种聚集率高有关, Cluster聚类树图结果与NMDS排序图结果相吻合, 在群落组成鉴定时, 可将大型底栖动物鉴定至属级或科级。

|
| 图 3 3个季节不同分类阶元Second-stage NMDS排序图 Fig. 3 Second-stage NMDS plot based on different taxonomic resolution similarity matrices among three seasons |

|
| 图 4 3个季节不同分类阶元Second-stage聚类树状图 Fig. 4 Second-stage Cluster plot based on different taxonomic resolution similarity matrices among three seasons |
群落组成对环境因子响应在不同数据类型、分类水平和季节间略有不同(表 3), 以选出最佳环境因子和Adj. R2为参考依据, 不同数据类型群落组成与环境因子之间的关系有明显的季节差异, 总体而言3个季节属级、科级对环境因子的响应结果均与种级相似, 定量科级数据对环境因子的响应略高于属级和种级, 夏、秋2季科级选择的环境因子多于属级和种级。而定性数据, 春、秋2季科级对环境因子响应最高, 但秋季科级选择的环境因子与种级略有不同, 夏季科级对环境因子的响应最低。
| 变量Variable | Adj. R2 | P | 环境变量Environmental variables |
| 春季Spring | |||
| 种Quan | 0.209 | 0.012 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 属Quan | 0.245 | 0.016 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 科Quan | 0.254 | 0.034 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 种Qual | 0.231 | 0.021 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 属Qual | 0.272 | 0.025 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 科Qual | 0.311 | 0.017 | 石油、总氮、总磷、悬浮物、叶绿素、分选系数、水深 |
| 夏Summer | |||
| 种Quan | 0.241 | 0.017 | 叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 属Quan | 0.241 | 0.017 | 叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 科Quan | 0.258 | 0.003 | 粉砂(%)、叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 种Qual | 0.293 | 0.003 | 粉砂(%)、叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 属Qual | 0.293 | 0.003 | 粉砂(%)、叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 科Qual | 0.269 | 0.002 | 叶绿素、盐度、悬浮物、中值粒径、分选系数、总氮、总磷、石油 |
| 秋季Autumn | |||
| 种Quan | 0.223 | 0.030 | 叶绿素、盐度、溶解氧、分选系数、水深 |
| 属Quan | 0.222 | 0.027 | 叶绿素、盐度、溶解氧、分选系数、水深 |
| 科Quan | 0.232 | 0.025 | 叶绿素、盐度、溶解氧、分选系数、水深、石油 |
| 种Qual | 0.233 | 0.029 | 叶绿素、盐度、溶解氧、分选系数、水深、石油 |
| 属Qual | 0.230 | 0.027 | 叶绿素、盐度、溶解氧、分选系数、水深 |
| 科Qual | 0.306 | 0.024 | 叶绿素、悬浮物、溶解氧、分选系数、石油、总氮、总磷 |
| (Adj. R2为校正后环境因子解释量, P<0.05为显著相关) | |||
本研究针对辽河口3个航次大型底栖动物进行分析, 共采集到大型底栖动物105种, 环节动物为主要类群, 其次是节肢动物、软体动物和棘皮动物, 主要类群组成与辽东湾已开展调查结论相似[26-28], 在2007年至2014年间, 辽东湾采集到的物种数介于78—85, 说明辽河口与近7年辽东湾大型底栖动物整体数量大致相同。
本研究利用定量数据(丰度)和定性数据(有/无)验证分类充分性在群落组成分析中的有效性, 从统计结果来看, 将种级定量和定性数据鉴定到更高分类阶元(属级、科级)都是有效评估生物群落组成方法。3个季节种级与更高分类阶元Bray-Curtis距离和Sørensen距离相似矩阵之间均呈显著相关性(P < 0.001), 无论是定量数据还是定性数据的不同分类阶元间相似矩阵相关性系数均变化一致, 种级与属级的相关性(Rho=0.969, P < 0.001)高于种级与科级(Rho=0.888, P < 0.001)、目级(Rho=0.635, P < 0.001)的相关性, 说明属级和科级数据的群落组成与种级一致性更强。有研究者提出不同分类阶元群落组成的一致性取决于更高分类阶元与种级之间的聚集率, 随着种级的属和属级的科增加, 会导致属级(或科级)/种级聚集率升高, 增加了种级与更高分类阶元的一致性概率, 形成群落组成趋同效应[29]。本文3个季节的属种聚集率范围为0.8—0.95, 科种聚集率范围为0.75—0.85, 与科级相比, 属种聚集率相对较高且聚集率都大于0.4, 因此辽河口区域物种辐射度低, 使用属级或者科级丢失信息较少, 此结果与国外研究底栖动物的结果相似[30-31]。运用Second-stage NMDS排序图进一步分析不同分类阶元的定量与定性数据的群落矩阵相对的位置关系, 结果表明相对位置越近, 其群落组成相关性越强。其中3个季节定量属级与种级的距离要近于种级与科级的距离, 因此种级与属级群落组成的一致更高;根据NMDS结果进一步做Cluster聚类树分析, 3个季节中, 在定量数据方面属级丢失信息最少(少于3%), 科级丢失信息少于6%, 这一结果与NMDS排序图吻合, 进而说明属级与种级群落组成的一致性最高, 这优于厦门湾和苏北地区底栖动物属级丢失8%和小于5%, 科级丢失20%和7%结果[2-3], 由此说明辽河口定量数据中, 科级的群落组成与种级群落组成相似性也很高, 丢失信息少, 可代替性强。
在群落组成与环境因子响应研究中, 定量科级数据被环境因子解释量略高于属级和种级数据, 根据“Hierarchical-response-to-stress”准则[32], 大型底栖动物不同分类阶元区分群落的改变主要依赖于人类活动干扰强度。普遍认为自然环境的变化是通过迫使物种替换进而影响群落结构, 人类活动的干扰则会促进更高分类阶元的物种分类聚集。在污染严重区域, 耐污种会聚集, 不耐污物种将会分散分布, 已有研究表明较高分类阶元尤其是科级水平很好地反映人为干扰对种级水平群落变化[33-35], 辽河口污染严重, 科级水平对环境响应高于种级水平的结果验证了此观点。此外, 更高分类阶元与种级聚集率高时, 同属级或同科级的物种没有明显的辐射, 大型底栖动物扩散能力差, 耐污染能力强, 可能具有相同的生态位, 因此这些物种对环境变化反应趋同, 辽河口科级聚集率φ>0.4[23], 说明在环境受到污染时, 耐污染物种敏感性低, 在科级水平高密度聚集, 因此科级对环境因子响应高于种级。
不同分类阶元对环境因子响应结果在不同季节存在差异, 因为不同季节的解释变量变化范围不一致, 春季和夏季是辽河口的雨季[36], 秋季是旱季, 辽河口上游有稻田和养殖区[37], 春季和夏季有大量的施肥, 会导致辽河口的氮盐、磷酸盐等浓度升高[19], 导致春、夏2季节的人为活动干扰是影响河口水质为主要因素, 而秋季海洋自然环境是河口水质的主要影响因素, 因此, 不同季节的大型底栖动物群落组成不同, 在污染的环境中, 耐污种会聚集, 因此每个季节的分类变量对环境的响应不一致。辽河口多毛类(Polychaeta)在不同季节占比不同, 春季占31.58%, 夏季占48.39%, 秋季占29.82%。研究者指出, Polychaeta被认为是可以代替整个大型底栖动物对生态系统健康状况评价的一类生物类群, 对栖息环境污染耐受性高同时对环境变化敏感性低的一类大型底栖动物, 在环境受到扰动时, 可以高密度聚集, 当群落中Polychaeta为主要类群, 则可以采用定性或者科级Polychaeta数据代替种级数据[38-39]。
关于群落组成与环境响应的另一个研究是关于定量与定性数据的相关应用对比, 通常定量数据优于定性数据, 因为前者包含生物体对环境响应更多信息, 相比定量丰度数据, 定性数据可以更容易、更有效地收集[40], 定性数据中物种Pielou′s evenness为1, 不存在优势种的干扰, 但在分类充分性的研究背景下定性数据不同分类阶元也会对环境响应提供另一种简便的研究思路, 尤其是对于广泛的地理范围研究, 种级定性数据在环境指示群落方面与种级定量数据具有同样的效用[41]。如果区域中种级定量数据的丰度、均匀度的差异大, 会导致群落组成被环境因子解释的结果有很大的不同, 得到的信息也会比定性数据清晰[5, 42]。本文只有春季不同分类阶元定性数据对环境响应变化与定量数据一致(Adj. R2和环境变量), 因此定性数据不能推广到所有应用中[43]。
TS是用来监测海洋生物的一种简化方法, 目的是用更高分类阶元或者定性数据来获得跟种级定量数据一样的结果以减少鉴定的时间和人力成本[44], 有研究者指出在鉴定至属级、科级水平时只用了77%和55%的种级鉴定开支, 并且属级相比于科级缺失的信息较少[45]。鉴定到种级对海洋生物的完整性发育程度要求很高, 将其鉴定至更高分类水平会减少鉴定失误率, 在有些区域会出现单一优势种, 这会对后续多样性分析造成干扰, 但鉴定至更高分类阶元会合并优势物种, 提高多样性研究的准确性, 这使得TS在海洋监测中广泛应用, 尤其以保护为导向的研究应优先关注物种级别数据, 而快速评估研究如果能够很好地描述受胁迫的生态系统的生态状况, 则可能会利用较高的分类阶元。根据本文结果, 在人力、物力等条件有限情况下开展辽河口大型底栖动物监测, 根据实际需求可将大型底栖生物鉴定放宽至属级或科级。
| [1] |
Marshall J C, Steward A L, Harch B D. Taxonomic resolution and quantification of freshwater macroinvertebrate samples from an Australian dryland river: the benefits and costs of using species abundance data. Hydrobiologia, 2006, 572(1): 171-194. DOI:10.1007/s10750-005-9007-0 |
| [2] |
吴东浩, 徐兆安, 王备新, 张咏, 厉以强, 沈燕飞. 大型底栖无脊椎动物不同分类阶元对水生态评价的影响. 环境科学研究, 2012, 25(1): 24-29. |
| [3] |
朱晓芬, 陈彬, 俞炜炜, 林俊辉, 黄雅琴, 廖建基. 厦门湾大型底栖动物分类学多样性指数及分类充分性. 生态学报, 2018, 38(15): 5554-5565. |
| [4] |
Ellis D. Taxonomic sufficiency in pollution assessment. Marine Pollution Bulletin, 1985, 16(12): 459. DOI:10.1016/0025-326X(85)90362-5 |
| [5] |
Terlizzi A, Anderson M J, Bevilacqua S, Fraschetti S, Włodarska-Kowalczuk M, Ellingsen K E. Beta diversity and taxonomic sufficiency: do higher-level taxa reflect heterogeneity in species composition?. Diversity and Distributions, 2009, 15(3): 450-458. DOI:10.1111/j.1472-4642.2008.00551.x |
| [6] |
Landeiro V L, Bini L M, Costa F R C, Franklin E, Nogueira A, De Souza J L P, Moraes J, Magnusson W E. How far can we go in simplifying biomonitoring assessments? An integrated analysis of taxonomic surrogacy, taxonomic sufficiency and numerical resolution in a megadiverse region. Ecological Indicators, 2012, 23: 366-373. DOI:10.1016/j.ecolind.2012.04.023 |
| [7] |
Aguado-Giménez F, Gairín J I, Martinez-Garcia E, Fernandez-Gonzalez V, Ballester Moltó M, Cerezo-Valverde J, Sanchez-Jerez P. Application of "taxocene surrogation" and "taxonomic sufficiency" concepts to fish farming environmental monitoring. Comparison of BOPA index versus polychaete assemblage structure. Marine Environmental Research, 2015, 103: 27-35. DOI:10.1016/j.marenvres.2014.10.006 |
| [8] |
Xiong W, Yang Y Z, Zhan A B. Reliability of simplifying strategies for rapid biodiversity assessment in studying community-environment interactions. Ecological Indicators, 2018, 85: 861-868. DOI:10.1016/j.ecolind.2017.11.050 |
| [9] |
Olsgard F, Somerfield P J. Surrogates in marine benthic investigations-which taxonomic unit to target?. Journal of Aquatic Ecosystem Stress and Recovery, 2000, 7(1): 25-42. DOI:10.1023/A:1009967313147 |
| [10] |
Resh V H, Unzicker J D. Water quality monitoring and aquatic organisms: the importance of species identification. Journal (Water Pollution Control Federation), 1975, 47(1): 9-19. |
| [11] |
Maurer D. The dark side of Taxonomic Sufficiency (TS). Marine Pollution Bulletin, 2000, 40(2): 98-101. DOI:10.1016/S0025-326X(99)00235-0 |
| [12] |
Elbrecht V, Vamos E E, Meissner K, Aroviita J, Leese F. Assessing strengths and weaknesses of DNA metabarcoding-based macroinvertebrate identification for routine stream monitoring. Methods in Ecology and Evolution, 2017, 8(10): 1265-1275. DOI:10.1111/2041-210X.12789 |
| [13] |
李巧. 生物多样性研究中的分类充分性研究. 云南农业大学学报, 2015, 30(4): 629-633. |
| [14] |
Moreno C E, Sanchez-Rojas G, Pineda E, Escobar F. Shortcuts for biodiversity evaluation: a review of terminology and recommendations for the use of target groups, bioindicators and surrogates. International Journal of Environment and Health, 2007, 1(1): 71-86. DOI:10.1504/IJENVH.2007.012225 |
| [15] |
Terlizzi A, Bevilacqua S, Fraschetti S, Boero F. Taxonomic sufficiency and the increasing insufficiency of taxonomic expertise. Marine Pollution Bulletin, 2003, 46(5): 556-561. DOI:10.1016/S0025-326X(03)00066-3 |
| [16] |
Yekta F A, Izadi S, Asgari M, Aghajan-Pour F, Shokri M. Higher taxa as surrogates for species richness in intertidal habitats of Qeshm Island in the Persian Gulf. Marine Biodiversity, 2017, 1-8. |
| [17] |
Bacci T, Trabucco B, Marzialetti S, Marusso V, Lomiri S, Vani D, Virno Lamberti C. Taxonomic sufficiency in two case studies: where does it work better?. Marine Ecology, 2009, 30(S1): 13-19. |
| [18] |
Domínguez-Castanedo N, Rojas-López R, Solís-Weiss V, Hernández-Alcántara P, Granados-Barba A. The use of higher taxa to assess the benthic conditions in the southern Gulf of Mexico. Marine Ecology, 2007, 28(S1): 161-168. |
| [19] |
罗西玲, 邢磊, 徐宾铎, 张美昭, 任一平. 辽河口芦苇湿地河蟹养殖区水体N、P营养盐和COD的变化. 海洋湖沼通报, 2016(3): 20-27. |
| [20] |
李少月, 隋吉星, 曲方圆, 王振钟, 赵宁, 于子山. 辽东湾六股河口邻近海域冬季大型底栖动物生态学特征. 海洋科学进展, 2016, 34(1): 95-105. |
| [21] |
李轶平, 王小林, 郭栋, 刘修泽, 付杰, 董婧. 辽东湾潮间带夏季大型底栖动物群落结构研究. 海洋环境科学, 2016, 35(2): 214-220. |
| [22] |
尹桂金, 严岩. 大亚湾冬季沉积物中叶绿素含量与分布. 生态学杂志, 2012, 31(11): 2834-2840. |
| [23] |
Bevilacqua S, Terlizzi A, Claudet J, Fraschetti S, Boero F. Taxonomic relatedness does not matter for species surrogacy in the assessment of community responses to environmental drivers. Journal of Applied Ecology, 2012, 49(2): 357-366. DOI:10.1111/j.1365-2664.2011.02096.x |
| [24] |
Clarke K R, Gorley R N. PRIMER V7: User Manual/Tutorial. Plymouth: PRIMER-E, Plymouth Ltd, 2015.
|
| [25] |
徐勇, 李新正, 王洪法, 张宝琳, 帅莲梅. 长江口邻近海域丰水季大型底栖动物群落特征. 生物多样性, 2016, 24(7): 811-819. |
| [26] |
刘录三, 孟伟, 郑丙辉, 李新正, 雷坤, 李中宇. 辽东湾北部海域大型底栖动物研究: I. 种类组成与数量分布. 环境科学研究, 2008, 21(6): 118-123. |
| [27] |
王振钟, 隋吉星, 曲方圆, 季相星, 赵宁, 于子山. 春季辽东湾西部海域大型底栖动物生态学研究. 海洋湖沼通报, 2013(1): 113-119. |
| [28] |
李轶平, 李玉龙, 付杰, 于旭光, 邹存杰. 辽东湾西海岸潮间带大型底栖动物群落生态位特征. 海洋科学, 2019, 43(3): 32-39. |
| [29] |
Bowman M F, Bailey R C. Does taxonomic resolution affect the multivariate description of the structure of freshwater benthic macroinvertebrate communities?. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(8): 1802-1807. |
| [30] |
Sánchez-Moyano J E, Fa D A, Estacio F J, García-Gómez J C. Monitoring of marine benthic communities and taxonomic resolution: an approach through diverse habitats and substrates along the Southern Iberian coastline. Helgoland Marine Research, 2006, 60(4): 243-255. |
| [31] |
Heino J, Soininen J. Are higher taxa adequate surrogates for species-level assemblage patterns and species richness in stream organisms?. Biological Conservation, 2007, 137(1): 78-89. |
| [32] |
Dauvin J C, Gomez Gesteira J L, Salvande Fraga M. Taxonomic sufficiency: an overview of its use in the monitoring of sublittoral benthic communities after oil spills. Marine Pollution Bulletin, 2003, 46(5): 552-555. |
| [33] |
Vijapure T, Sukumaran S. Optimization of the taxonomic resolution of an indicator taxon for cost-effective ecological monitoring: Perspectives from a heterogeneous tropical coastline. Journal of Environmental Management, 2019, 247: 474-483. |
| [34] |
Bevilacqua S, Claudet J, Terlizzi A. Best practicable aggregation of species: a step forward for species surrogacy in environmental assessment and monitoring, 2013, Ecology and Ecolution, 3: 3780-3793.
|
| [35] |
Soares-Gomes A, Mendes C L T, Tavares M, Santi L. Taxonomic sufficiency of polychaete taxocenes for estuary monitoring. Ecological Indicators, 2012, 15(1): 149-156. |
| [36] |
Zheng L, Liu Z T, Yan Z G, Zhang Y H, Yi X L, Zhang J, Zheng X, Zhou J L, Zhu Y. pH-dependent ecological risk assessment of pentachlorophenol in Taihu Lake and Liaohe River. Ecotoxicology and Environmental Safety, 2017, 135: 216-224. |
| [37] |
Qin R G, Wu Y Q, Xu Z G, Xie D, Zhang C. Assessing the impact of natural and anthropogenic activities on groundwater quality in coastal alluvial aquifers of the lower Liaohe River Plain, NE China. Applied Geochemistry, 2013, 31: 142-158. |
| [38] |
Checon H H, Amaral A C Z. Taxonomic sufficiency and the influence of rare species on variation partitioning analysis of a polychaete community. Marine Ecology, 2017, 38(1): e12384. |
| [39] |
Nóbrega-Silva C, Patrício J, Marques J C, Dos Santos Olímpio M, Farias J N B, Molozzi J. Is polychaete family-level sufficient to assess impact on tropical estuarine gradients?. Acta Oecologica, 2016, 77: 50-58. |
| [40] |
Lenat D R, Resh V H. Taxonomy and stream ecology-the benefits of genus- and species-level identifications. Journal of the North American Benthological Society, 2001, 20(2): 287-298. |
| [41] |
Melo A S. Effects of taxonomic and numeric resolution on the ability to detect ecological patterns at a local scale using stream macroinvertebrates. Archiv für Hydrobiologie, 2005, 164(3): 309-323. |
| [42] |
Cushman S A, McGarigal K. Patterns in the species-environment relationship depend on both scale and choice of response variables. Oikos, 2004, 105(1): 117-124. |
| [43] |
Heino J. Taxonomic surrogacy, numerical resolution and responses of stream macroinvertebrate communities to ecological gradients: are the inferences transferable among regions?. Ecological Indicators, 2014, 36: 186-194. |
| [44] |
Jones F C. Taxonomic sufficiency: the influence of taxonomic resolution on freshwater bioassessments using benthic macroinvertebrates. Environmental Reviews, 2008, 16: 45-69. |
| [45] |
Ferraro S P, Cole F A. Taxonomic level sufficient for assessing pollution impacts on the southern california bight macrobenthos-revisited. Environmental Toxicology and Chemistry, 1995, 14(6): 1031-1040. |