 2020, Vol. 40
2020, Vol. 40文章信息
- 姜琦, 郭润泉, 宋涛涛, 陈廷廷, 陈宇辉, 贾林巧, 熊德成, 陈光水
- JIANG Qi, GUO Runquan, SONG Taotao, CHEN Tingting, CHEN Yuhui, JIA Linqiao, XIONG Decheng, CHEN Guangshui
- 增温与氮添加对不同季节杉木幼苗细根不同形态氮吸收动力学的影响
- Effects of warming and nitrogen addition on nitrogen uptake kinetics of fine roots of Cunninghamia lanceolata seedlings in different seasons
- 生态学报. 2020, 40(9): 2996-3005
- Acta Ecologica Sinica. 2020, 40(9): 2996-3005
- http://dx.doi.org/10.5846/stxb201905070924
-
文章历史
- 收稿日期: 2019-05-07
- 网络出版日期: 2020-03-16

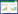




2. 福建师范大学湿润亚热带生态地理过程教育部重点实验室, 福州 350007
2. Key Laboratory for Humid Subtropical Eco-Geographical Processes of the Ministry of Education, Fujian Normal University, Fuzhou 350007, China
植物对全球变化的响应是根和茎之间协调相互作用的一系列复杂过程的产物[1]。根系作为植物吸收养分和水分的重要器官[2-3], 在森林生态系统能量和物质循环中发挥着巨大的作用[4], 是森林NPP和土壤固碳的基础[5]。植物根系可以通过调整其生理、寿命、形态和结构特征来改变其养分获取能力, 以满足地上部养分需求的变化[6]。Fransen等[7]发现一些植物根系增殖与养分吸收之间的关系并不密切, 它们主要通过细根的生理调整来获取养分资源。根系吸收养分的生理能力作为影响养分获取的调节因素之一, 它对植物环境变化的响应可能是某些物种对全球变化更为敏感的一个重要的机制[8]。离子吸收动力学是将Miehaelis-Menten方程应用到解释植物对介质中离子吸收动态过程的一种理论[9], 它是研究植物养分吸收的重要手段, 为描述根系养分吸收特性、评价不同物种对环境养分状况的适应性、鉴定并筛选具有高效吸收能力的物种提供了重要的途径[10]。
全球变暖已是不争的事实, IPCC第五次评估报告中预测21世纪末全球地表将平均增温0.3—4.8℃[11]。大量研究表明, 温度升高可能直接或间接地影响森林生态系统的地上和地下生态过程[12-14]。土壤温度升高可能对根系的生长产生负面影响[15-16], 而当根系生长受到不利影响时, 将通过增强根系吸收动力学以增加植物养分吸收的可能性, 以改善植物营养状况[8]。目前关于野外增温对根系养分吸收动力学影响的研究尚未报道, 一些通过室内培养增温的方式对植物不同形态氮吸收速率影响的研究结果却不尽相同, 有促进, 有抑制, 也有无作用的情况[17-19], 而增温引起根系养分吸收能力变化的确切机制则更不清楚。因此在全球变暖的背景下研究模拟增温对根系氮吸收速率的影响显得尤为必要。
目前, 全球氮沉降十分严重, 预计到21世纪末全球氮沉降速率会增加2—3倍[20]。尤其我国中亚热带地区已成为全球三大氮沉降区之一[21]。因此, 全球生态系统对氮沉降的响应成为国际研究热点问题[22]。生态系统对氮素的固持, 既依赖于微生物的固氮, 也依赖于植被的吸收能力[23]。目前一些研究表明, 氮的添加会降低根系吸收氮的能力[24-25], 但这种反应的程度可能在很大程度上取决于植物对氮的需求[26]。因此不同物种的氮吸收动力学对氮添加的响应还有待进一步确认。根系氮吸收动力学的变化可能会影响陆地生态系统碳和氮的负荷程度。尽管氮吸收动力学在决定碳和氮在植物和生态系统循环中起着重要作用, Bassirirad等[8]表示迫切需要在这一领域产生更多的数据, 以改进对全球环境预计变化的长期植物反应的预测, 但此后很长一段时间关于根系养分吸收能力是否或如何对全球环境变化作出反应的信息依旧很少。
土壤温度和土壤水分具有季节性变化, 从而使土壤养分矿化速率和养分有效性表现出明显的季节变化特点;同时, 一年中植物的生长亦表现出不同的生长节律[27], 导致在不同季节对土壤养分的需求不一。因而, 植物根系的养分吸收动力学特征可能亦表现出明显的季节差异。然而, 目前有关植物氮吸收速率季节性差异的研究鲜有报道, 但了解不同季节的植物的养分吸收动力学的变化, 对于理解植物养分利用策略和提供栽培管理水平等具有重要作用。
我国人工林面积占世界1/3, 亚热带地区是我国最重要的人工林基地, 而杉木(Cunninghamia lanceolata)是中国湿润亚热带区重要的造林和用材树种, 在我国人工林生产和森林碳汇中占有重要地位[28]。目前有关杉木根系氮吸收动力学特性的了解很少, 而增温与氮添加是否改变杉木氮吸收动力学特征则更不清楚。基于此, 本文研究增温和氮添加对杉木细根不同季节氮吸收动力学的影响, 这对于理解杉木养分获取策略以及对全球变化的适应性等均有重要意义。我们的目的是:(1)探讨不同季节杉木细根的氮吸收速率特性;(2)探讨杉木细根氮吸收动力学对土壤增温和氮添加的响应。
1 研究地区与研究方法 1.1 研究区概况研究区位于福建三明森林生态系统与全球变化研究站陈大观测点, 金丝湾森林公园陈大林业采育场(26°19′N, 117°36′E)内。该区属中亚热带季风气候, 年均温19.1 ℃, 年降雨量1749 mm(多集中在3—8月), 年均蒸发量1585 mm, 相对湿度81%。本研究区地形以低山丘陵为主, 平均海拔300 m, 平均坡度25°—35°;土壤以花岗岩发育的红壤和黄壤为主。
1.2 试验设计试验采用完全随机设计, 依据IPCC第五次评估报告对21世纪末全球地表平均增温的预测[11]和本研究区氮沉降水平的背景值为36 kg N hm-2 a-1[29], 本试验设计为增温(Warming, 两水平, 不增温和增温+4℃)×氮添加(Nitrogen addition, 两水平, 不施氮和施氮+40 kg N hm-2 a-1)双因子试验, 共4个处理, 即对照(CT), 增温(Warming), 施氮(Nitrogen)和增温+施氮(Warming+Nitrogen), 每个处理6个重复。共建立24个小区, 每个小区面积为2.5 m×2.5 m, 小区内土壤取自附近杉木林(0—40 cm), 取出的土壤在去除杂物后分层充分混拌均匀, 以消除土壤异质性, 然后分层回填进实验样地中, 并采用压实法调整土壤容重与取土杉木林基本相同。

|
| 图 1 不同处理树高生长情况(平均值±标准差) Fig. 1 Height growth of trees with different treatments (Mean±SD) 不同小写字母代表不同处理间差异显著 |
土壤增温为安装加热电缆主动持续增温, 其方式是在每个小区内平行布设, 深度10 cm, 间隔20 cm, 并在最外围环绕一圈电缆, 以保证增温的均匀性。电缆布设完成后在各个样地10 cm深处各布设1个土壤温度(德国JUMO)和湿度传感器(美国Decagon)用于监测土壤温度和湿度的变化。施氮为每月月初以溶液的形式对小区进行喷洒, 全年共喷洒12次。将每个小区所需的NH4NO3溶解在800 mL去离子水中(溶液浓度约为1.67 g/L), 其方式为用手提式喷雾器在小区四周从幼苗林冠上方对小区均匀施加氮肥, 未施加氮肥的小区喷洒等量的去离子水。2015年1月在各个小区内种植5棵树高(53.0±2.2 cm)与地径(4.8±0.5 mm)相近的一年生三代杉木幼苗, 每株杉木幼苗种植在两条电缆线中间位置, 于2016年3月8日正式开始增温, 同年对施氮处理施加氮肥(NH4NO3, 分析纯), 同时定期清除小区内自然萌发的其他植物。
1.3 环境因子测定分别于2017年4月、8月和10月在每个小区取土芯(0—10 cm), 及时挑选出其中细根, 过2 mm筛用于土壤NH4+-N、NO3--N及土壤含水率的测定;取部分过2 mm筛的土在自然条件下风干后, 用于土壤pH的测定;剩余风干土过100目筛用于土壤C、N、P的测定。
| 季节 Season |
处理 Treatments |
温度 Temperature/℃ |
含水率 Moisture/% |
全碳 Total carbon/ (mg/g) | 全氮 Total nitrogen/ (mg/g) | 全磷 Total phosphorus/ (mg/g) | 铵态氮 Ammonium nitrogen/ (mg/kg) | 硝态氮 Nitrate nitrogen/ (mg/kg) |
| 春季 Spring | 对照 | 19.49±0.4b | 15.83±0.74a | 12.49±1.29a | 1.16±0.12a | 0.78±0.07 | 4.56±0.15b | 1.39±0.57b |
| 增温 | 23.21±1.97a | 14.63±0.49bc | 10.21±0.71b | 0.98±0.05b | 0.69±0.06 | 6.19±0.58a | 1.77±0.59ab | |
| 施氮 | 19.29±0.41b | 15.57±0.64ab | 12.04±1.31a | 1.12±0.11a | 0.77±0.05 | 6.00±0.86a | 2.59±0.93a | |
| 增温+施氮 | 23.06±2.3a | 14.43±0.92c | 10.99±1.14ab | 1.03±0.11ab | 0.76±0.08 | 6.20±0.42a | 1.96±0.52ab | |
| 夏季 Summer | 对照 | 27.59±0.47b | 8.00±0.9a | 9.41±1.07 | 0.92±0.11 | 0.25±0.03 | 5.55±0.33b | 1.40±0.22 |
| 增温 | 30.8±2.49a | 7.00±0.4b | 8.34±0.89 | 0.84±0.09 | 0.25±0.07 | 6.98±0.67a | 1.30±0.53 | |
| 施氮 | 27.27±0.55b | 7.00±0.8ab | 8.99±0.66 | 0.89±0.04 | 0.27±0.01 | 5.15±0.26b | 1.12±0.07 | |
| 增温+施氮 | 29.88±2.48a | 7.00±0.4b | 8.48±0.50 | 0.86±0.05 | 0.24±0.03 | 6.49±0.42a | 1.45±0.66 | |
| 秋季 Autumn | 对照 | 23.07±0.88b | 8.00±0.6ab | 9.02±1.47 | 0.83±0.13 | 0.28±0.04 | 3.48±0.11b | 1.56±0.13ab |
| 增温 | 27.44±1.15a | 7.00±1.5b | 9.04±0.84 | 0.86±0.07 | 0.29±0.04 | 5.35±1.97a | 1.45±0.37b | |
| 施氮 | 23.02±0.65b | 9.00±1.3a | 9.12±0.45 | 0.84±0.03 | 0.29±0.02 | 3.82±0.11b | 1.64±0.31ab | |
| 增温+施氮 | 26.52±2.27a | 7.00±1.4b | 9.54±0.69 | 0.89±0.05 | 0.23±0.03 | 3.94±0.55ab | 1.92±0.36a | |
| 同列不同小写字母表示同一季节处理间差异显著(P < 0.05) | ||||||||
本研究采用15N标记法[30]测定细根氮吸收速率。分别于2017年春季、夏季、秋季在每个小区挖取表层(0—10 cm)细根, 随后洗净挑出活细根(0—1 mm), 取11份0.1 g鲜重的细根, 用0.5 mmol的CaCl2溶液将细根表面的土壤颗粒洗去, 以保证细胞膜的完整性[31]。然后将10份细根浸泡于装有0.5 mmol CaCl2溶液(20℃)的烧杯中, 再将细根分别浸泡于10、50、100、250、500 μmol的15NH4Cl溶液与K15NO3溶液(100 mL, 20℃)中15 min(两种溶液各5份细根)。随后用5 mmol KCl和0.5 mmol CaCl2溶液将浸泡后的细根分别冲洗3次(每次100 mL左右), 以洗去细根表面所吸附的15N。冲洗后的细根置于70℃烘箱中烘干(24 h), 再用球磨仪对烘干后的细根进行研磨, 最后用同位素质谱仪(Finnigan MAT-253, Thermo Electron)测定细根15N含量, 并用元素分析仪(vario EL III Element Analyzer, 德国)测定样品全N含量。剩余的1份细根在烘干磨碎后用来测定自然丰度下细根中的15N。NH4+-N吸收速率通过15NH4Cl溶液浸泡后细根中总15N含量减去空白对照中细根的15N获得(NO3--N吸收速率通过同样方法获得), 即公式为:

|
(1) |
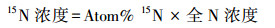
|
(2) |
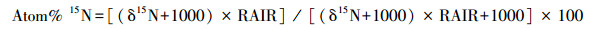
|
(3) |
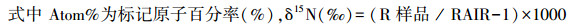
|
(4) |
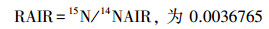
|
(5) |
Michaelis-Menten动力学参数通过采用Lineweaver & Burke双倒数法[31]变换计算获得, 其公式为:
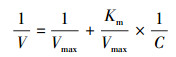
|
式中, V为离子吸收速率(μmol g-1h-1), C为离子浓度(μmol/L), Vmax为最大吸收速率(μmol g-1h-1), Km为半饱和常数。
1.5 数据分析利用SPSS 21.0软件对数据进行统计分析。采用混合线性模型分析季节、增温与氮添加对细根不同形态氮吸收动力学参数的影响, 采用双因素方差分析检验不同季节下增温与氮添加对细根不同形态氮吸收动力学参数的影响。利用Origin 9.0软件作图, 图表中数据为平均值±标准差。
2 结果与分析 2.1 不同季节不同处理下杉木细根的氮吸收速率图 2表明在三个季节中, 四个处理的杉木离体根对不同浓度下硝态氮的吸收速率基本呈现出春季较高, 夏秋季较低的态势, 而其对不同浓度下铵态氮的吸收速率基本呈现出夏秋季较高, 春季较低的态势。不同季节四个处理的离体根对不同浓度下铵态氮的吸收均遵循米氏-曼氏动力学方程(R2均大于0.90, P < 0.01), 铵态氮的吸收速率呈现出随着铵态氮溶液浓度的增加而提高的趋势。三个季节相同溶液浓度下, 铵态氮的吸收要比硝态氮吸收快。四个处理的离体根对不同浓度下硝态氮的吸收并不完全遵循米氏方程, 杉木的离体根在约100μmol处表现出初始饱和。在10—50 μmol浓度下呈上升趋势, 而在50—100 μmol浓度下呈下降趋势, 随后又转为上升趋势。由于本研究区土壤硝态氮浓度远不及100 μmol溶液中硝态氮浓度, 且无法估计其Vmax和Km, 因此本文采用100 μmol时的硝态氮吸收速率作为我们的硝态氮吸收能力的指标进行分析。

|
| 图 2 不同季节不同处理下细根硝态氮和铵态氮吸收动力学曲线(平均值±标准差) Fig. 2 Fine roots kinetics of nitrate uptake curve of different seasons and treatments (Mean±SD) |
由表 2可得, 季节对Vmax-NH4+、Km-NH4+以及V100-NO3-吸收均有极显著影响(P < 0.01)。土壤增温对Vmax-NH4+有显著影响(P < 0.05), 但对Km-NH4+和V100-NO3-吸收都没有显著影响。氮添加对Vmax-NH4+有极显著影响(P < 0.01), 但对Km-NH4+和V100-NO3-吸收都没有显著影响。土壤增温与氮添加间的交互作用对Km-NH4+有显著影响(P < 0.05)。然而, 季节、土壤增温与氮添加间对Vmax-NH4+和V100-NO3-吸收均没有显著的交互作用。
| 变异来源 Source of variance |
Vmax-NH4+ | Km-NH4+ | V100-NO3- | |||||
| F | P | F | P | F | P | |||
| S | 17.509 | < 0.01* | 28.575 | < 0.01* | 12.43 | < 0.01* | ||
| W | 5.194 | 0.026* | 0.664 | 0.418 | 0.757 | 0.388 | ||
| N | 8.533 | 0.005* | 0.643 | 0.426 | 0.615 | 0.436 | ||
| S×W | 0.278 | 0.758 | 2.337 | 0.105 | 3.815 | 0.028 | ||
| S×N | 1.318 | 0.275 | 2.635 | 0.08 | 1.668 | 0.197 | ||
| W×N | 0.004 | 0.947 | 5.535 | 0.022* | 0.719 | 0.4 | ||
| S×W×N | 0.39 | 0.679 | 0.894 | 0.414 | 0.536 | 0.588 | ||
| S:季节因子Season factor; W:增温因子Warming factor; N:氮添加因子Nitrogen addition factor; *P < 0.05 | ||||||||
由图 3可得, 春季, 氮添加对Vmax-NH4+有极显著影响(P < 0.01), 但对Km-NH4+和V100-NO3-吸收都没有显著影响;与没有氮添加相比, 氮添加使Vmax-NH4+提高了36%(图 3)。增温及其与氮添加的交互作用对Vmax-NH4+、Km-NH4+以及V100-NO3-吸收均没有显著影响。

|
| 图 3 不同季节下不同处理细根Vmax-NH4+、Km-NH4+与V100-NO3-吸收值(平均值±标准差) Fig. 3 Fine roots Vmax-NH4+, Km-NH4+ and V100-NO3- uptake values in different seasons and treatments (Mean ± SD) W:增温因子Warming factor; N:氮添加因子Nitrogen addition factor; *表示显著性(*P < 0.05), ns表示无显著性 |
夏季, 增温对Vmax-NH4+有显著影响(P < 0.05), 但对Km-NH4+和V100-NO3-吸收都没有显著影响;与没有增温相比, 增温使Vmax-NH4+提高了16%(图 3)。氮添加对Vmax-NH4+、Km-NH4+以及V100-NO3-吸收均没有显著影响。氮添加与土壤增温的交互作用对Vmax-NH4+、Km-NH4+及V100-NO3-吸收均没有显著影响。
秋季, 增温对Km-NH4+有显著影响(P < 0.05), 但对Vmax-NH4+和V100-NO3-吸收都没有显著影响;与没有增温相比, 增温使Km-NH4+降低了17%(图 3)。氮添加对Km-NH4+有显著影响(P < 0.05), 但对Vmax-NH4+和V100-NO3-吸收都没有显著影响;与没有氮添加相比, 氮添加使Km-NH4+提高了19%(图 3b)。增温与氮添加的交互作用对Vmax-NH4+、Km-NH4+以及V100-NO3-吸收均没有显著影响。
3 讨论 3.1 杉木细根的不同形态氮吸收速率特性本研究中, 季节对铵态氮吸收动力学参数及硝态氮吸收速率均有极显著的影响, 不同季节下四个处理杉木离体根的对应浓度硝态氮吸收速率基本呈现出春季较高, 夏秋季较低的态势。一方面, 冬季作为亚热带林木的非生长季, 其对氮素的需求较少, 因此经过冬天氮素的积累, 有效氮大量富集于春季土壤中[32]。另一方面, 本研究区春季一般为雨季, 春季硝态氮的淋溶损失可能增加。因此细根可能对环境产生适应性, 从而提高其在春季对硝态氮的吸收速率。但本研究中杉木对不同季节铵态氮的吸收速率态势则相反, 为夏秋季较高, 春季较低。这可能因为夏季的高温加速了土壤氮矿化速率(特别是铵化速率), 增加了土壤铵态氮的浓度[32], 并且植物需要获取大量养分以满足其生长需要(图 1)。此外, 由于植物需要消耗更多的能量去获取硝态氮[33], 因此植物一般会更偏爱铵态氮。从而杉木可能为适应土壤养分环境, 在夏秋季更多地同化铵态氮。
不同季节杉木离体根对铵态氮的吸收速率均符合Michaelis-Menten动力学方程, 对硝态氮的吸收速率均不符合Michaelis-Menten动力学方程。这与Rothstein等[30]的研究结果相似, 他们在密歇根州对毛白杨(Populus tremuloides)细根的氮吸收速率研究发现, 毛白杨离体根对铵态氮的吸收符合Michaelis-Menten动力学, 而对硝酸盐的吸收速率在约100 μmol · L-1处表现出初始饱和, 并不符合此方程。但周晓红等[34]对蕹菜(Ipomoea aquatica)的研究发现, 其不同形态氮素吸收动力学吸附曲线均符合Michaelis-Menten动力学方程。另外, Sato等[35]对裙带菜(Undaria pinnatifida)不同形态氮素吸收动力学特性进行了研究研究发现, NO3-和NH4+浓度与裙带菜氮吸收速率之间均不符合Michaelis-Menten动力学方程, 而呈双曲线回归关系。这可能因为高等植物对硝酸盐的吸收通常表现为双相动力学, 在较高浓度时, 高亲和力转运体达到初始饱和, 然后是线性反应[36]。另外, 已确定在浓度为0—1000 μmol · L-1的条件下, 植物根系吸收NO3-具有三个传输系统, 前两个系统表现出可饱和的行为, 而第三个系统是线性的, 第二个可饱和运输系统和线性系统被称为低亲和力运输系统(LATS), 在相对较高的离子浓度下对总的吸收有很大的贡献[37]。
杉木离体根对铵态氮的吸收速率符合Michaelis-Menten动力学方程, 但铵态氮吸收速率仍未趋于平缓。这与许多采取离体根吸收的试验方法的研究结果相似, 其铵态氮吸收速率曲线随溶液氮浓度的提高并未呈现出趋于平缓的态势[38-39], 但不少通过营养液培养的方法研究氮吸收速率的试验结果均呈现出氮吸收速率曲线随营养液氮浓度的提高而趋于平缓的规律[40-41], 这可能与试验方法上的差异有关, 离体根有切除伤口, 其伤口直接暴露在外, 更易于离子的渗入, 从而间接的提高其吸收能力, 使得氮吸收速率曲线不易趋于平缓。另外, 本研究中杉木细根对铵态氮的吸收速率在溶液氮浓度达到500 μmol仍未趋于平缓, 魏旭等[42]在中国东北地区对长白落叶松(Larix olgensis)氮素吸收的研究发现, 长白落叶松的铵态氮吸收速率曲线在营养液氮浓度达到2000 μmol时亦未趋于平缓;而Wijk等[43]在对道格拉斯冷杉(Abies fabri)的研究结果表明, 其铵态氮吸收速率曲线在营养液氮浓度达到200 μmol时却已趋于平缓。这表明不同物种对NH4+离子的吸收水平不尽相同, 因此在实际林业生产中应根据当地养分状况合理选择树种进行培育。
杉木离体根对铵态氮的吸收速率明显远高于对硝态氮的吸收速率。Wijk等[43]对道格拉斯冷杉(Abies fabri)幼树的研究同样发现铵态氮的吸收速率要高于硝态氮的吸收速率。张熙灵等[9]对香蒲(Typha orientalis)、芦苇(Phragmites australis)与藨草(Scirpus triqueter)的研究也发现, 三种植物对NH4+的吸收速率均高于NO3-的吸收速率, NH4+是这三种植物主要吸收的无机氮形态。树木对NH4+的吸收能力通常大于吸收NO3-的能力[44-45], 但在草本植物中可能存在不同情况, 如何明洁等[46]对烟草的研究发现, 其对NH4+的吸收能力和速率均劣于对NO3-的吸收, 他们认为烟草吸收的主要氮形态可能是NO3-。这表明植株对不同形态氮吸收速率的快慢可能与植物种类有关。
3.2 增温与氮添加对杉木细根氮吸收动力学的影响夏季土壤增温使得杉木细根对铵态氮的最大吸收速率显著增加, 但对硝态氮的吸收速率无显著影响。这与一些通过室内培养增温试验的研究结果相似, 如Cruz等[17]对角豆树(Ceratonia siliqua)的研究发现, 当温度从15℃增到25℃时, 角豆树增加了对铵态氮的吸收速率, 而对硝态氮的吸收速率无显著影响。Marschner等[47]对挪威的云杉研究发现, 当温度从5℃增到15℃时, 云杉对铵态氮的吸收速率随温度的增加而增加, 但对硝态氮的吸收速率无显著影响。Gessler等[19]对山毛榉(Fagus sylvatica)的研究发现, 当温度从20℃增到25℃时, 山毛榉对铵态氮的吸收速率显著增加, 而对硝态氮的吸收速率无显著影响。但亦有一些不同的报道。如Gessler等[19]发现当温度从20℃增到25℃时, 云杉(Picea abies)硝态氮的吸收速率显著增加, 而对铵态氮的吸收速率无显著影响。Bassirirad等[18]采用铺设塑料薄膜增温的方式对沙生冰草(Agropyron desertorum)的研究发现, 增温降低了冰草对硝态氮的最大吸收速率。目前温度引起植物养分吸收能力变化的确切机制尚不清楚。一方面可能因为温度的变化可以通过改变NH4+[44]、NO3-[18]的根系传输特性而直接影响植物养分的获取。另一方面, 根区温度也可能通过改变根系质膜中脂肪酸的流动性而影响养分吸收能力[48]。在本研究中, 可能因为夏季增温提高了土壤养分矿化速率, 促进了细根氮吸收, 提高了细根氮含量[49], 并提高了根内氮同化酶的数量, 从而提高了铵态氮的最大吸收速率。此外, 植物更偏爱铵态氮, 会更多的同化利用铵态氮, 因为植物可能需要更多的能量吸收利用硝态氮[33]。Km表示载体对离子的亲和力, Km越小, 载体对离子的亲和力越高, 与载体数目及其在运转过程中的效率有关[50]。本研究中秋季土壤增温下的Km-NH4+显著降低, 这可能是因为土壤增温延长了树木的生长期, 植物对养分吸收需求更大。杉木养分吸收的亲和力提高, 对其来说是更好的吸收利用养分的策略。
春季氮添加使得杉木细根对铵态氮的最大吸收速率显著增加, 但对硝态氮的吸收速率无显著影响。Bassirirad等[39]对美国北卡罗来纳州的火炬松(Pinus taeda)研究发现, 氮添加显著降低了其对铵态氮的吸收能力, 对硝态氮的吸收能力无显著影响。Rothstein等[30]对毛白杨(Populus tremuloides)的研究发现, 增加土壤氮的有效性显著降低其对NH4+和NO3-的吸收能力。Jones等[51]人在瑞典北部研究发现, 施氮后的樟子松(Pinus sylvestris)根系的NH4+吸收速率仅在一个生长季节就比未施肥的对照植物减少了近四倍。Högberg等[52]测量了瑞典北部至法国南部若干森林物种对NO3-和NH4+的吸收能力, 发现较高的N沉降与植物NH4+吸收速率呈负相关。这可能解释了氮沉降对NH4+吸收的抑制作用, 因为高浓度的根系N被认为是氮吸收的反馈因素[53]。根系吸收氮的动力学及其对增施氮的反应在不同物种之间有很大差异[25], 并往往还与物种的生长速度有很强的相关性[54]。本研究中, 施氮显著增加了杉木细根对铵态氮的最大吸收速率, 这可能因为杉木处在幼树阶段, 建构树冠需要大量的氮, 并且春季为生长期, 对养分的需求大, 从而提高了其对铵态氮的吸收能力。另外, 本研究中秋季氮添加使得杉木细根的Km-NH4+显著提高, 表明杉木细根对NH4+的亲和力降低, 这可能因为秋季不是植物的旺盛生长期, 氮添加反而降低了杉木对NH4+的吸收能力。本研究中增温与氮添加对细根V100-NO3-无显著交互作用, 一方面可能因为硝态氮本身移动性较强, 易吸收。另一方面硝态氮可能主要通过植物蒸腾作用随水分被吸收, 导致增温与氮添加的交互作用对硝态氮吸收的影响不显著。
4 结论四个处理的杉木细根对不同浓度下硝态氮的吸收速率在春季较高, 夏秋季较低, 而其对不同浓度下铵态氮的吸收速率却在夏秋季较高, 春季较低。不同季节下杉木细根对不同浓度铵态氮的吸收均遵循米氏-曼氏动力学方程, 而对硝态氮的吸收并不完全遵循米氏-曼氏动力学, 表现为双相动力学。并且三个季节相同溶液浓度下, 铵态氮的吸收要比硝态氮吸收快。氮添加提高了杉木幼苗春季的Vmax-NH4+与秋季的Km-NH4+, 增温提高了夏季的Vmax-NH4+, 降低了秋季的Km-NH4+, 但增温与氮添加对不同季节的V100-NO3-都无显著影响。三个季节下, 增温与氮添加对Vmax-NH4+、Km-NH4+和V100-NO3-都没有显著的交互作用。本文的研究结论是基于杉木幼树氮吸收动力学的测定结果, 不同物种的不同形态氮吸收动力学对增温与氮添加的响应可能不同, 未来应拓宽增温与氮添加对氮吸收动力学研究的物种范围, 以进一步揭示植物应对全球变化的养分利用策略。
| [1] |
Farrar J F, Jones D L. The control of carbon acquisition by roots. New Phytologist, 2000, 147(1): 43-53. DOI:10.1046/j.1469-8137.2000.00688.x |
| [2] |
Kochsiek A, Tan S, Russo S E. Fine root dynamics in relation to nutrients in oligotrophic Bornean rain forest soils. Plant Ecology, 2013, 214(6): 869-882. DOI:10.1007/s11258-013-0215-9 |
| [3] |
Pinno B D, Wilson S D. Fine root response to soil resource heterogeneity differs between grassland and forest. Plant Ecology, 2013, 214(6): 821-829. DOI:10.1007/s11258-013-0210-1 |
| [4] |
Solly E F, Schöning I, Boch S, Kandeler E, Marhan S, Michalzik B, Müller J, Zscheischler J, Trumbore S E, Schrumpf M. Factors controlling decomposition rates of fine root litter in temperate forests and grasslands. Plant and Soil, 2014, 382(1/2): 203-218. |
| [5] |
Kou L, Jiang L, Fu X L, Dai X Q, Wang H M, Li S G. Nitrogen deposition increases root production and turnover but slows root decomposition in Pinus elliottii plantations. New Phytologist, 2018, 218(4): 1450-1461. DOI:10.1111/nph.15066 |
| [6] |
Wang G L, Fahey T J, Xue S, Liu F. Root morphology and architecture respond to N addition in Pinus tabuliformis, West China. Oecologia, 2013, 171(2): 583-590. DOI:10.1007/s00442-012-2441-6 |
| [7] |
Fransen B, de Kroon H, Berendse F. Root morphological plasticity and nutrient acquisition of perennial grass species from habitats of different nutrient availability. Oecologia, 1998, 115(3): 351-358. DOI:10.1007/s004420050527 |
| [8] |
Bassirirad H. Kinetics of nutrient uptake by roots:responses to global change. New Phytologist, 2000, 147(1): 155-169. DOI:10.1046/j.1469-8137.2000.00682.x |
| [9] |
张熙灵, 王立新, 刘华民, 清华, 刘东伟, 王炜, 梁存柱, Recknagel F. 芦苇、香蒲和藨草3种挺水植物的养分吸收动力学. 生态学报, 2014, 34(9): 2238-2245. |
| [10] |
余勤, 邝炎华. 根系养分吸收动力学及应用. 华南农业大学学报, 1997, 18(2): 105-110. |
| [11] |
IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013.
|
| [12] |
Xiong D C, Yang Z J, Chen G S, Liu X F, Lin W S, Huang J X, Bowles F P, Lin C F, Xie J S, Li Y Q, Yang Y S. Interactive effects of warming and nitrogen addition on fine root dynamics of a young subtropical plantation. Soil Biology and Biochemistry, 2018, 123: 180-189. DOI:10.1016/j.soilbio.2018.05.009 |
| [13] |
宋涛涛, 陈光水, 史顺增, 郭润泉, 郑欣, 熊德成, 陈望远, 陈廷廷. 土壤增温对杉木幼苗细根呼吸和非结构性碳的影响. 应用生态学报, 2018, 29(3): 705-712. |
| [14] |
Leppälammi-Kujansuu J, Salemaa M, Kleja D B, Linder S, Helmisaari H S. Fine root turnover and litter production of Norway spruce in a long-term temperature and nutrient manipulation experiment. Plant and Soil, 2014, 374(1/2): 73-88. |
| [15] |
陈望远, 熊德成, 史顺增, 宋涛涛, 蔡瑛莹, 郭润泉, 陈廷廷, 郑欣, 陈光水. 土壤增温对杉木幼苗细根生长量及形态特征的影响. 生态学报, 2018, 38(15): 5305-5314. |
| [16] |
陈云玉, 熊德成, 邓飞, 许辰森, 冯建新, 史顺增, 钟波元, 陈光水. 土壤增温与氮添加对杉木幼苗细根径级根长分布的影响. 应用生态学报, 2016, 27(4): 1009-1014. |
| [17] |
Cruz C, Lips S H, Martins-Loução M A. The effect of nitrogen source on photosynthesis of carob at high CO2 concentrations. Physiologia Plantarum, 1993, 89(3): 552-556. DOI:10.1111/j.1399-3054.1993.tb05212.x |
| [18] |
Bassirirad H, Caldwell M M, Bilbrough C. Effects of soil temperature and nitrogen status on kinetics of 15NO3- uptake by roots of field-grown Agropyron desertorum (Fisch. ex Link) Schult. New Phytologist, 1993, 123(3): 485-489. DOI:10.1111/j.1469-8137.1993.tb03760.x |
| [19] |
Gessler A, Schneider S, von Sengbusch D, Weber P, Hanemann U, Huber C, Rothe A, Kreutzer K, Rennenberg H. Field and laboratory experiments on net uptake of nitrate and ammonium by the roots of spruce (Picea abies) and beech (Fagus sylvatica) trees. New Phytologist, 1998, 138(2): 275-285. DOI:10.1046/j.1469-8137.1998.00107.x |
| [20] |
史顺增, 熊德成, 邓飞, 冯建新, 许辰森, 钟波元, 陈云玉, 陈光水, 杨玉盛. 土壤增温、氮添加及其交互作用对杉木幼苗细根生产的影响. 植物生态学报, 2017, 41(2): 186-195. |
| [21] |
Liu X J, Duan L, Mo J M, Du E Z, Shen J L, Lu X K, Zhang Y, Zhou X B, He C E, Zhang F S. Nitrogen deposition and its ecological impact in China:an overview. Environmental Pollution, 2011, 159(10): 2251-2264. DOI:10.1016/j.envpol.2010.08.002 |
| [22] |
郭润泉, 熊德成, 宋涛涛, 蔡瑛莹, 陈廷廷, 陈望远, 郑欣, 陈光水. 模拟氮沉降对杉木幼苗细根化学计量学特征的影响. 生态学报, 2018, 38(17): 6101-6110. |
| [23] |
Nadelhoffer K J, Downs M R, Fry B, Aber J D, Magill A H, Melillo J M. The fate of 15N-labelled nitrate additions to a northern hardwood forest in eastern Maine, USA. Oecologia, 1995, 103(3): 292-301. DOI:10.1007/BF00328617 |
| [24] |
Clarkson D T. Factors affecting mineral nutrient acquisition by plants. Annual Review of Plant Physiology, 1985, 36: 77-115. DOI:10.1146/annurev.pp.36.060185.000453 |
| [25] |
Berry W L. Plant nutrition. An introduction to current concepts. A. D. M. Glass. The Quarterly Review of Biology, 1989, 64(4): 499-499. |
| [26] |
Forde B G, Clarkson D T. Nitrate and ammonium nutrition of plants:physiological and molecular perspectives. Advances in Botanical Research, 1999, 30: 1-90. DOI:10.1016/S0065-2296(08)60226-8 |
| [27] |
Huang J X, Chen G S, Yang Z J, Xiong D C, Guo J F, Xie J S, Robinson D, Yang Y S. Understory fine roots are more ephemeral than those of trees in subtropical Chinese fir (Cunninghamia lanceolata (Lamb.) Hook) stands. Annals of Forest Science, 2016, 73(3): 657-667. DOI:10.1007/s13595-016-0551-8 |
| [28] |
Piao S L, Ciais P, Friedlingstein P, Peylin P, Reichstein M, Luyssaert S, Margolis H, Fang J Y, Barr A, Chen A P, Grelle A, Hollinger D Y, Laurila T, Lindroth A, Richardson A D, Vesala T. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature, 2008, 451(7174): 49-52. DOI:10.1038/nature06444 |
| [29] |
史顺增, 熊德成, 冯建新, 许辰森, 钟波元, 邓飞, 陈云玉, 陈光水, 杨玉盛. 模拟氮沉降对杉木幼苗细根的生理生态影响. 生态学报, 2017, 37(1): 74-83. |
| [30] |
Rothstein D E, Zak D R, Pregitzer K S, Curtis P S. Kinetics of nitrogen uptake by Populus tremuloides in relation to atmospheric CO2 and soil nitrogen availability. Tree Physiology, 2000, 20(4): 265-270. |
| [31] |
王梦柳, 樊卫国. 刺梨实生苗对硝态氮、铵态氮的吸收与利用差异分析. 果树学报, 2017, 34(6): 682-691. |
| [32] |
刘志江, 林伟盛, 杨舟然, 林廷武, 刘小飞, 陈岳民, 杨玉盛. 模拟增温和氮沉降对中亚热带杉木幼林土壤有效氮的影响. 生态学报, 2017, 37(1): 44-53. DOI:10.3969/j.issn.1673-1182.2017.01.009 |
| [33] |
韩兴国, 李凌浩, 黄建辉.生物地球化学概论.北京: 高等教育出版社, 1999.
|
| [34] |
周晓红, 王国祥, 杨飞, 何伟, 杨周. 空心菜对不同形态氮吸收动力学特性研究. 水土保持研究, 2008, 15(5): 84-87. |
| [35] |
Sato Y, Hirano T, Niwa K, Suzuki T, Fukunishi N, Abe T, Kawano S. Phenotypic differentiation in the morphology and nutrient uptake kinetics among Undaria pinnatifida cultivated at six sites in Japan. Journal of Applied Phycology, 2016, 28(6): 3447-3458. DOI:10.1007/s10811-016-0857-9 |
| [36] |
Pilbeam D J, Kirkby E A. The Physiology of Nitrate Uptake Nitrogen in Higher Plants. New York: Y P Abrol John Wiley and Sons, 1990: 39-64.
|
| [37] |
Glass A D M, Siddiqi M Y. Nitrogen absorption by plant roots//Srivastava H S, Singh R P, eds. Nitrogen Nutrition in Higher Plants. New Delhi, India: Associated Publishing Co, 1995: 21-56.
|
| [38] |
Bassirirad H, Griffin K L, Strain B R, Reynolds J F. Effects of CO2 enrichment on growth and root uptake rate of loblolly pine and ponderosa pine seedlings. Tree Physiology, 1996, 16(11/12): 957-962. |
| [39] |
Bassirirad H, Thomas R B, Reynolds J F, Strain B R. Differential responses of root uptake kinetics of NH4+ and NO3- to enriched atmospheric CO2 concentration in field-grown loblolly pine. Plant, Cell & Environment, 1996, 19(3): 367-371. |
| [40] |
任军, 徐程扬, 林玉梅, 周睿智, 张爱花. 水曲柳幼苗根系吸收不同形态氮的动力学特征. 植物生理学通讯, 2008, 44(5): 919-922. |
| [41] |
段英华, 张亚丽, 沈其荣. 增硝营养对不同基因型水稻苗期吸铵和生长的影响. 土壤学报, 2005, 42(2): 260-265. DOI:10.3321/j.issn:0564-3929.2005.02.013 |
| [42] |
魏红旭, 徐程扬, 马履一, 江俐妮, 李雪莲, 杨卓. 长白落叶松幼苗对铵态氮和硝态氮吸收的动力学特征. 植物营养与肥料学报, 2010, 16(2): 407-412. |
| [43] |
Wijk C K V, Prins H B A. The kinetics of NH4+ and NO3- uptake by Douglas fir from single N-solutions and from solutions containing both NH4+ and NO3-. Plant and Soil, 1993, 151(1): 91-96. DOI:10.1007/BF00010789 |
| [44] |
Chapin III F S, van Cleve K, Tryon P R. Relationship of ion absorption to growth rate in taiga trees. Oecologia, 1986, 69(2): 238-242. DOI:10.1007/BF00377628 |
| [45] |
Wallenda T, Read D J. Kinetics of amino acid uptake by ectomycorrhizal roots. Plant, Cell & Environment, 1999, 22(2): 179-187. |
| [46] |
何明洁, 范腾飞, 杨超, 陈益银, 李迪秦, 李常军, 刘来华. 15N短时标记法鉴定烟草'K326'和'红花大金元'吸收NO3-和NH4+的生理动力学特征. 植物生理学报, 2017, 53(4): 572-580. |
| [47] |
Marschner H, Häussling M, George E. Ammonium and nitrate uptake rates and rhizosphere pH in non-mycorrhizal roots of Norway spruce[Picea abies (L.) Karst.]. Trees, 1991, 5(1): 14-21. |
| [48] |
Clarkson D T, Earnshaw M J, White P J, Cooper H D. Temperature dependent factors influencing nutrient uptake:an analysis of responses at different levels of organization. Symposia of the Society for Experimental Biology, 1988, 42: 281-309. |
| [49] |
冯建新, 熊德成, 史顺增, 许辰森, 钟波元, 邓飞, 陈云玉, 陈光水, 杨玉盛. 土壤增温对杉木幼苗细根生理生态性质的影响. 生态学报, 2017, 37(1): 35-43. |
| [50] |
张亚娟, 刘存歧, 李洪波, 吴亦红, 王亚斌. 蕹菜对富营养化水体的氮磷去除及吸收动力学研究. 环境工程学报, 2011, 5(5): 1057-1061. |
| [51] |
Jones H E, Högberg P, Ohlsson H. Nutritional assessment of a forest fertilisation experiment in northern Sweden by root bioassays. Forest Ecology and Management, 1994, 64(1): 59-69. DOI:10.1016/0378-1127(94)90127-9 |
| [52] |
Högberg P, Högbom L, Schinkel H. Nitrogen-related root variables of trees along an N-deposition gradient in Europe. Tree Physiology, 1998, 18(12): 823-828. DOI:10.1093/treephys/18.12.823 |
| [53] |
Siddiqi M Y, Glass A D M, Ruth T J, Rufty T W. Studies of the uptake of nitrate in barley. I. Kinetics of 13NO3- influx. Plant Physiology, 1990, 93(4): 1426-1432. |
| [54] |
Schenk M K. Regulation of nitrogen uptake on the whole plant level. Plant and Soil, 1996, 181(1): 131-137. DOI:10.1007/BF00011299 |