 2020, Vol. 40
2020, Vol. 40文章信息
- 常海涛, 刘佳楠, 赵娟, 张静, 罗雅曦, 刘任涛, 左小安
- CHANG Haitao, LIU Jianan, ZHAO Juan, ZHANG Jing, LUO Yaxi, LIU Rentao, ZUO Xiaoan
- 乌拉特荒漠草原红砂(Reaumuria soongorica)灌丛林地地面节肢动物群落季节分布特征
- Seasonal distribution of ground-active arthropod community in Reaumuria soongorica shrubbery land in Urat desert steppe
- 生态学报. 2020, 40(9): 3121-3130
- Acta Ecologica Sinica. 2020, 40(9): 3121-3130
- http://dx.doi.org/10.5846/stxb201904150756
-
文章历史
- 收稿日期: 2019-04-15
- 网络出版日期: 2020-03-16





2. 宁夏大学农学院, 银川 750021;
3. 中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站, 兰州 730000
2. College of Agriculture, Ningxia University, Yinchuan 750021, China;
3. Urat Desert-Grassland Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China
荒漠草原是草原向荒漠过渡的草原类型, 通常位于由半干旱地区向干旱地区过渡的敏感地带, 往往由于人类活动和气候变化的影响极易发生退化和沙漠化[1-2]。在乌拉特荒漠草原, 红砂灌丛具有生态可塑性强、抗旱、耐盐和集沙等优良特性, 已成为维持该荒漠草原脆弱生态系统稳定性的重要植被类型, 关于其生态效应的研究引起越来越多的关注[3]。研究表明, 红砂灌丛属于一种低矮型耐盐性灌丛, 常常由于“生境岛”作用而导致灌丛林地空间异质性增强。并且, 随着季节更替, 降雨和温度的急剧变化, 调控着灌丛林地空间异质性的作用方向和强度, 进而将影响节肢动物的时空分布[4]。因此, 深入开展乌拉特荒漠草原红砂灌丛林地地面节肢动物的群落组成及其季节分布特征研究, 对于维持脆弱荒漠草原生态系统的稳定性、生物多样性保育及进行有效的管理均具有重要的理论与实践意义。
已有研究结果表明, 地面节肢动物的活动密度易受到季节性降水、温度和土壤环境等因子的影响, 而产生不同的季节分布格局[5-6]。关宏斌等[7]通过对科尔沁沙地土壤动物垂直分布、季节动态及群落多样性研究, 发现中小型土壤动物活动密度表现为春季>秋季>夏季;大型土壤动物幼虫为夏季>秋季>春季;成虫为秋季>春季>夏季。刘任涛等[5]通过研究宁夏荒漠草原地面节肢动物群落组成及季节动态特征, 发现随着季节变化地面节肢动物群落组成和个体数分布差异性较大, 而类群数和生物量则呈现出相对稳定的季节分布格局。刘继亮等[8-9]通过研究黑河中游沙质荒漠地面节肢动物群落组成及多样性季节变异, 发现不同地面节肢动物类群的季节变异规律不同;如拟步甲科的活动密度随季节变化表现为秋季>夏季>春季, 而蚁科则表现为夏季>秋季>春季, 即随着季节性降水及温度的动态变化, 地面节肢动物的活动密度、群落组成分布和多样性结构均会随之发生显著变化。但是, 目前对于乌拉特荒漠草原红砂灌丛林地地面节肢动物群落组成和季节动态分布尚不清楚, 以及环境因子对地面节肢动物的影响也缺乏相关的研究。
鉴于此, 本文以乌拉特荒漠草原红砂灌丛林地作为研究样地, 分别于2018年春季、夏季和秋季, 利用国际通用的陷阱诱捕法, 结合土壤理化性质, 分析红砂灌丛林地地面节肢动物的群落组成、多样性季节分布特征及其与环境因子间的关系。旨在为内蒙古乌拉特荒漠草原生态系统维持生物多样性、灌丛林利用与保护及退化生态系统的恢复与评价提供科学依据。
1 研究区概况研究区位于内蒙古巴彦淖尔市乌拉特后旗(41°25′ N、106°58′ E;海拔1650 m)。属于典型的大陆性干旱气候, 年均气温为5.3 ℃, 最高气温37 ℃, 最低气温-34.4 ℃;≥10 ℃的年积温为2000—3000 ℃。年均降雨量为180 mm, 且年际变化率大, 降水主要集中在夏季(7—8月), 占全年降雨量的70%左右;年蒸发量是降雨量的10余倍。年无霜期为130 d左右。年均风速为5 m/s, 每年风速>10 m/s的天数多达52 d。土壤类型主要以棕钙土和灰棕漠土为主[2, 10]。
本试验选取的红砂灌丛林地位于中国科学院西北生态环境资源研究院乌拉特荒漠草原研究站的典型荒漠草原试验场内。该试验场于2010年开始围封, 面积约333.34 hm2。其中, 红砂灌丛林地的主要植被包括红砂(Reaumuria soongorica)、柠条(Caragana korshinskii)、小针茅(Stipa klemenzii)、多根葱(Allium polyrhizum)、猪毛菜(Salsola collina)和骆驼蓬(Peganum harmala)等[2, 10]。本试验调查于2018年开展, 研究区域内年降雨量为251 mm, 平均气温为7.18℃, 变异系数(CV)分别为1.88和2.02。其中, 5月份降雨量为8 mm, 平均气温为18.93℃;8月份降雨量为125.4 mm, 平均气温为21.22℃;10月份无降雨, 平均气温为5.97℃(图 1)。

|
| 图 1 2018年各月平均降雨量和气温 Fig. 1 Mean monthly rainfall and air temperature in 2018 |
于2018年5月, 在中国科学院乌拉特荒漠草原研究站的红砂灌丛林地试验场内, 设置5个重复样地。每个样地间间距为15—20 m(避免地面节肢动物活动区域重叠, 保证每个陷阱杯的相对独立性), 面积均为50 m×50 m。在每个研究样地内, 布设12个调查样点[包括灌丛内和灌丛外(指距离灌丛根部4—5 m以外空白区域)][11];每个调查样点之间距离为5—10 m, 距离样地边缘10 m以上。5个研究样地内共布设60个样点(12个样点×5个重复样地)。分别于5月(春季)、8月(夏季)和10月(秋季)进行调查。
2.2 节肢动物取样与鉴定在每个调查样点, 采用国际通用的陷阱诱捕法调查地面节肢动物群落组成与数量分布。陷阱具体布设方法:将收集器杯口齐地面(上表面直径7 cm, 下表面直径5 cm, 高度10 cm)埋入土壤中;在其内加入浓度为3 %的福尔马林溶液和少量甘油, 以增加诱捕的有效性。每次布设陷阱持续时间均为7 d, 然后将收集到的地面节肢动物标本保存于75%的酒精中, 带回实验室进行鉴定统计[9]。
动物标本分类鉴定主要依据《中国土壤动物检索图鉴》[12]、《昆虫分类》[13]和《宁夏贺兰山昆虫》[14]等工具书鉴定至科水平。然后, 将鉴定和统计后的地面节肢动物分类放在洁净的酒精中清洗表面泥土, 用纸巾或滤纸吸干其表面酒精, 用万分之一的微量天平进行称重、记录[15]。根据不同节肢动物类群在群落总个体数中所占比例将其划分为优势类群(占群落总个体数的10%以上)、常见类群(1%—10%)和稀有类群(<1%)[16]。
2.3 土壤样品采集与分析在春季、夏季和秋季的每个调查样点内, 均利用铝盒采集剖面土壤测定土壤含水量(0—10 cm)。并且用铁铲采集1个0—10 cm的表层混合土样, 带回实验室, 共取土样180个(12个样点×5个重复样地×3个季节)。然后, 在自然状态下风干, 过2 mm土壤筛, 去除其中的杂质(如草根和叶片等), 用于测定土壤pH、电导率、土壤全碳、全氮含量及土壤粒径组成。
土壤含水量(%)采用烘干称重法测定。土壤pH和电导率(μS/m)均采用1:5土水比浸提后, 分别使用PHS-3C酸度计与便携式电导率仪(雷磁DDSJ-308F)进行测定。土壤全碳(g/kg)和全氮(g/kg)采用元素分析仪(意大利DK6, UDK140分析仪)进行测定。土壤粒径组成采用Mastersizer3000激光衍射粒度分析仪进行测定。根据美国农业部(USAD)制土壤质地分级标准划分土壤质地:沙粒(50—2000 μm)、黏粉粒(0—50 μm)[17]。
2.4 数据处理与统计分析将每个样地内的12个调查样点内所采集的地面节肢动物数量进行合并计算, 以便为了保证红砂灌丛林地的样地代表性。其中, 群落指数包括Shannon-Wiener指数(H)和Simpson指数(D)。

|
(1) |

|
(2) |
式中, 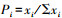
所有数据均采用SPSS 20.0软件进行统计分析。采用单因素方差分析(One-way ANOVA)和多重比较法比较不同数据组间的差异性。显著性水平为P<0.05。
通过对3个季节的地面节肢动物类群分布进行除趋势对应分析(Detrended correspondence analysis, DCA), 计算出排序轴的梯度长度(Lengths of gradient, LGA)=2.53<4, 故采用线性模型的冗余对应分析(Redundancy analysis, RDA)来研究地面节肢动物群落分布与土壤环境因子间的关系。应用国际通用排序软件CANOCO 4.5进行分析运算[18]。
3 结果与分析 3.1 土壤环境特征由表 1可知, 土壤含水量在3个季节间存在显著差异(P<0.05), 具体表现为夏季>秋季>春季。土壤pH均表现为夏季显著低于春季和秋季(P<0.05), 而后两个季节间无显著差异。土壤沙粒含量表现为春季显著高于夏季(P<0.05), 而与秋季间无显著差异, 且夏季和秋季间亦无显著差异。土壤黏粉粒含量表现为夏季显著高于春季(P<0.05), 而与秋季无显著差异, 且春季和秋季间亦无显著差异。在春季、夏季和秋季3个季节内, 土壤全碳含量表现为秋季显著高于春季和夏季(P<0.05), 而后两个季节间无显著差异。土壤全氮和土壤电导率在3个季节间均无显著差异(P>0.05)。
| 季节 Seasons |
土壤全碳 Soil total Carbon/(g/kg) |
土壤全氮 Soil total Nitrogen/(g/kg) |
土壤含水量 Soil moisture content/% |
土壤pH Soil pH |
土壤电导率 Soil electric conductivity/(μS/m) |
土壤黏粉粒 Soil clay plus silt content/% |
土壤沙粒 Soil sand content/% |
| 春Spring | 9.88±0.39b | 0.27±0.03a | 0.79±0.00c | 7.92±0.04a | 304.29±45.37a | 54.17±2.90b | 45.83±2.90a |
| 夏Summer | 10.29±1.23b | 0.30±0.02a | 7.33±0.00a | 7.22±0.04b | 287.42±52.16a | 61.76±2.10a | 38.24±2.10b |
| 秋Autumn | 12.81±0.45a | 0.31±0.03a | 4.36±0.00b | 7.96±0.02a | 362.68±48.27a | 59.13±2.19ab | 40.86±2.20ab |
| 同列不同小写字母表示差异显著(P<0.05) | |||||||
由表 2可知, 在3个季节内, 共捕获560只地面节肢动物, 隶属于10目23科, 共有类群有8个。其中, 优势类群包括拟步甲科和蚁科, 其个体数占总个体数的63.21%;常见类群包括象甲科、天牛科、步甲科、近管蛛科、平腹蛛科、跳蛛科、蟹蛛科、长奇盲蛛科和鼠妇科共9个类群, 其个体数占总个体数的31.43%;稀有类群包括12个类群, 其个体数占总个体数的5.36%。
| 编码 Code |
科属 Familys/genus |
春Spring | 夏Summer | 秋Autumn | |||||
| 多度 Abundance/% |
生物量 Biomass/% |
多度 Abundance/% |
生物量 Biomass/% |
多度 Abundance/% |
生物量 Biomass/% |
||||
| 1 | 蜈蚣科Solopendra subspinipes | 0.98 | 1.32 | ||||||
| 2 | 木伪蝎科Neobisiidae | 0.77 | 0.00 | ||||||
| 3 | 长奇盲蛛科Phalangiidae | 0.89 | 0.10 | 1.95 | 2.27 | ||||
| 4 | 狼蛛科Lycosidae | 0.77 | 0.17 | ||||||
| 5 | 蟹蛛科Thomisidae | 1.54 | 0.03 | 1.78 | 0.26 | 3.90 | 0.25 | ||
| 6 | 管巢蛛科Clubionidae | 0.77 | 0.02 | ||||||
| 7 | 跳蛛科Salticidae | 3.85 | 0.01 | 0.45 | 0.00 | 2.44 | 0.08 | ||
| 8 | 平腹蛛科Gnaphosidae | 9.23 | 1.29 | 4.00 | 0.65 | 6.83 | 1.37 | ||
| 9 | 隆头蛛科Eresidae | 2.31 | 0.25 | ||||||
| 10 | 近管蛛科Anyphaonidae | 9.23 | 0.61 | 0.45 | 0.01 | 1.46 | 0.16 | ||
| 11 | 鼠妇科Armadillidium vulgare Latreille | 5.78 | 0.67 | 0.49 | 0.00 | ||||
| 12 | 盲蝽科Miridae | 0.76 | 0.00 | 1.33 | 0.00 | 0.49 | 0.00 | ||
| 13 | 螽蟖科Anoisotima | 0.76 | 0.03 | 0.45 | 1.70 | ||||
| 14 | 拟步甲科Tenebrionidae | 51.54 | 88.59 | 13.33 | 72.56 | 64.88 | 80.90 | ||
| 15 | 象甲科Curculionidae | 6.15 | 7.21 | 8.44 | 8.95 | 5.37 | 8.69 | ||
| 16 | 粪金龟科Geotrupidae Latreille | 0.77 | 1.26 | 0.89 | 4.20 | ||||
| 17 | 蜉金龟科Aphodiidae | 0.44 | 0.01 | ||||||
| 18 | 天牛科Cerambycidae | 8.00 | 7.26 | ||||||
| 19 | 葬甲科Silphidae | 0.98 | 0.78 | ||||||
| 20 | 步甲科Carabidae | 4.62 | 0.46 | 6.67 | 3.39 | 1.46 | 4.13 | ||
| 21 | 食蚜蝇科Syrphidae | 0.77 | 0.03 | 1.46 | 0.03 | ||||
| 22 | 蚁科Formicidae | 6.15 | 0.01 | 45.78 | 0.08 | 6.34 | 0.01 | ||
| 23 | 蚁蜂科Mutillidae | 1.33 | 0.17 | ||||||
根据个体数量划分, 地面节肢动物的优势类群春季为拟步甲科, 夏季为拟步甲科和蚁科, 秋季为拟步甲科, 个体数分别占总个体数的51.54%、59.11%和64.88%, 共有的优势类群为拟步甲科。常见类群春季和夏季均有8类, 秋季有10类, 其个体数分别占总个体数的43.08%, 37.33%和32.68%, 共有的常见类群为象甲科、平腹蛛科和蟹蛛科。稀有类群春季有7类, 夏季有6类, 秋季有3类, 其个体数分别占总个体数的5.38%、3.56%和2.44%, 共有的稀有类群为盲蝽科。
根据生物量划分, 3个季节内生物量所占比例超过10%的地面节肢动物类群均为拟步甲科, 分别占群落总生物量的88.59%、72.56%和80.90%。所占比例处于1%—10%的地面节肢动物类群, 春季有象甲科、粪金龟科和平腹蛛科3个类群, 占群落总生物量的9.76%;夏季有象甲科、粪金龟科、天牛科、步甲科和螽斯科5个类群, 占群落总生物量的25.51%;秋季有象甲科、步甲科、平腹蛛科、长奇盲蛛科和蜈蚣科5个类群, 占群落总生物量的17.78%。所占比例小于1%的地面节肢动物类群, 春季有12类, 占群落总生物量的1.62%;夏季有10类, 占群落总生物量的1.94%;秋季有8类, 占群落总生物量的1.32%。
3.3 节肢动物群落结构季节分布由图 2可知, 地面节肢动物的群落结构随季节更替发生显著变化。其中, 地面节肢动物密度表现为夏季显著高于春季(P<0.05), 而春季和夏季均与秋季间无显著差异。土壤动物类群数和Shannon-Wiener指数的水平分布变化, 均表现为夏季显著高于春季和秋季(P<0.05), 而后两个季节间无显著差异。Simpson指数则表现为夏季显著高于秋季(P<0.05), 而春季和秋季均与夏季间无显著差异。

|
| 图 2 荒漠草原地面节肢动物群落指数季节分布 Fig. 2 Seasonal community index of ground-active arthropods in desert steppe 小写字母表示不同季节间存在显著差异性(P<0.05) |
地面节肢动物个体数分布与环境因子间关系的RDA排序分析表明(图 3)。第1型轴(F=9.47, P=0.016)和所有典型轴(F=3.10, P=0.030)在统计学上均达到了显著水平, 说明排序轴能够较好的反映地面节肢动物个体数分布与环境因子的关系。并且, 前两个排序轴累积解释了77.60%的地面节肢动物群落变异。从图 3可以看出, 与第1排序轴相关性较大的是气温、降雨量、土壤含水量、pH和电导率, 其中气温、降雨量、土壤含水量与排序轴1呈显著正相关(相关系数分别为0.723、0.840和0.601), 土壤pH和电导率与排序轴1呈显著负相关(相关系数分别为-0.898和-0.503)。与第2排序轴相关性较大的是土壤全碳含量, 与排序轴2呈显著负相关(相关系数为-0.497)。

|
| 图 3 地面节肢动物群落分布与环境因子关系的RDA二维排序图 Fig. 3 RDA two-dimensional diagram of the relationship between ground-active arthropods and soil factors TC:土壤全碳Soil total carbon; TN:土壤全氮Soil total nitrogen; SM:土壤含水量Soil moisture; pH:土壤pH Soil pH; EC:电导率Electrical conductivity; CS:土壤黏粉粒Soil clay plus silt content; SS:土壤沙粒Soil sand content; AT:气温Air temperature; R:降水量Rainfall; 图中数字表示地面节肢动物的类群编码, 具体内容见表 2 |
由图 3和表 3可知, 天牛科、步甲科、蚁科、蚁蜂科和鼠妇科等类群与排序轴1呈显著正相关。其中, 天牛科个体数分布与气温、降雨量和土壤含水量呈显著正相关关系(P<0.01), 与pH呈显著负相关关系(P<0.01), 与土壤黏粉粒含量呈正相关关系(P<0.05), 与沙粒含量呈负相关关系(P<0.05);蚁科和鼠妇科个体数分布与降雨量呈正相关关系(P<0.05), 与土壤含水量呈显著正相关关系(P<0.01), 与pH呈显著负相关关系(P<0.01), 且蚁科个体数分布与土壤黏粉粒含量呈正相关关系(P<0.05), 与沙粒含量呈负相关关系(P<0.05);步甲科个体数分布与土壤含水量呈正相关关系(P<0.05), 与pH呈显著负相关关系(P<0.01);蚁蜂科个体数分布与土壤pH呈负相关关系(P<0.05)。拟步甲科、葬甲科、平腹蛛科等3个类群与排序轴1呈显著负相关。其中, 拟步甲科个体数分布与气温和降雨量呈显著负相关关系(P<0.01), 与土壤pH呈正相关关系(P<0.05);葬甲科和平腹蛛科个体数分布均与土壤电导率呈正相关关系(P<0.05)。近管蛛科与排序轴2均呈显著正相关, 其个体数分布与土壤含水量及土壤黏粉粒含量呈显著负相关关系(P<0.01), 与沙粒含量呈显著正相关关系(P<0.01)。蜈蚣科与排序轴2呈显著负相关关系, 其个体数分布与气温呈负相关关系(P<0.05), 与pH呈正相关关系(P>0.05)。
| 科属Familys/genus | AT | R | TN | TC | SM | pH | EC | CS | SS |
| 蜈蚣科Solopendra subspinipes | -0.590* | -0.345 | -0.341 | 0.454 | 0.045 | 0.569* | -0.045 | 0.182 | -0.182 |
| 木伪蝎科Neobisiidae | -0.124 | -0.235 | 0.124 | 0.000 | -0.371 | -0.124 | -0.371 | -0.247 | 0.247 |
| 长奇盲蛛科Phalangiidae | 0.166 | 0.140 | 0.046 | 0.120 | 0.314 | -0.058 | 0.258 | 0.217 | -0.217 |
| 狼蛛科Lycosidae | 0.186 | 0.201 | -0.186 | -0.186 | -0.309 | 0.155 | 0.062 | -0.433 | 0.433 |
| 蟹蛛科Thomisidae | -0.025 | -0.189 | -0.029 | -0.143 | 0.246 | 0.220 | 0.365 | 0.231 | -0.231 |
| 管巢蛛科Clubionidae | 0.062 | -0.235 | 0.031 | 0.062 | -0.433 | -0.031 | 0.186 | -0.371 | 0.371 |
| 跳蛛科Salticidae | -0.400 | -0.398 | 0.451 | 0.087 | -0.299 | 0.367 | 0.119 | -0.216 | 0.216 |
| 平腹蛛科Gnaphosidae | -0.050 | -0.257 | 0.223 | 0.341 | -0.327 | 0.143 | 0.615* | -0.286 | 0.286 |
| 隆头蛛科Eresidae | 0.127 | -0.046 | -0.321 | -0.368 | -0.356 | 0.082 | 0.139 | -0.441 | 0.441 |
| 近管蛛科Anyphaonidae | 0.206 | -0.082 | -0.067 | -0.208 | -0.676*** | 0.186 | 0.023 | -0.783*** | 0.783*** |
| 鼠妇科Armadillidium vulgare Latreille | 0.358 | 0.610* | 0.141 | 0.179 | 0.646*** | -0.737*** | -0.230 | 0.505 | -0.505 |
| 盲蝽科Miridae | -0.099 | -0.183 | 0.060 | -0.058 | 0.194 | 0.050 | -0.140 | 0.227 | -0.227 |
| 螽蟖科Anoisotima | -0.045 | 0.123 | 0.136 | -0.318 | 0.136 | -0.341 | -0.045 | 0.182 | -0.182 |
| 拟步甲科Tenebrionidae | -0.671*** | -0.782*** | 0.016 | 0.436 | -0.471 | 0.603* | 0.436 | -0.304 | 0.304 |
| 象甲科Curculionidae | 0.229 | 0.122 | -0.498 | 0.283 | 0.381 | -0.147 | -0.444 | 0.346 | -0.346 |
| 粪金龟科Geotrupidae Latreille | 0.205 | 0.101 | 0.036 | -0.338 | -0.060 | -0.230 | -0.350 | -0.072 | 0.072 |
| 蜉金龟科Aphodiidae | -0.062 | 0.403 | 0.434 | -0.124 | 0.371 | -0.434 | -0.186 | 0.433 | -0.433 |
| 天牛科Cerambycidae | 0.713*** | 0.831*** | -0.181 | -0.162 | 0.805*** | -0.798*** | -0.479 | 0.571* | -0.571* |
| 葬甲科Silphidae | -0.363 | -0.345 | 0.500 | 0.091 | -0.136 | 0.159 | 0.545* | -0.136 | 0.136 |
| 步甲科Carabidae | 0.426 | 0.434 | 0.092 | -0.116 | 0.534* | -0.700*** | -0.333 | 0.474 | -0.474 |
| 食蚜蝇科Syrphidae | -0.340 | -0.184 | -0.240 | 0.246 | -0.064 | 0.473 | 0.207 | 0.102 | -0.102 |
| 蚁科Formicidae | 0.361 | 0.547* | -0.011 | 0.104 | 0.702*** | -0.725*** | -0.458 | 0.583* | -0.583* |
| 蚁蜂科Mutillidae | 0.121 | 0.498 | 0.508 | -0.012 | 0.417 | -0.551* | 0.024 | 0.465 | -0.465 |
| AT:气温Air temperature;R:降水量Rainfall;TN:土壤全氮Soil total nitrogen;TC:土壤全碳Soil total carbon;SM:土壤含水量Soil moisture;pH:土壤pH Soil pH;EC:电导率Electrical conductivity;CS:土壤黏粉粒Soil clay plus silt content;SS:土壤沙粒Soil sand content;*表示在0.05水平(双侧)上相关性显著;***表示在0.01水平(双侧)上相关性显著 | |||||||||
在乌拉特荒漠草原生态系统中, 气候及降雨量随季节变化而发生变化, 显著影响到该区域的土壤理化性质及其养分元素等环境条件, 并且对于土壤生态系统也产生了深刻影响[19]。本研究中, 秋季土壤全碳含量显著高于春季和夏季, 这与吴永胜等[20]的研究结果相似。而土壤全氮受到土壤缓冲性能的影响, 促使土壤系统内部的氮元素处于动态平衡状态[21], 导致红砂灌丛林地内的土壤全氮在3个季节内无显著差异性。本研究中, 土壤含水量和黏粉粒含量均表现为夏季最高, 秋季次之, 春季最低;而土壤沙粒含量则与黏粉粒含量相反。乌拉特荒漠草原区春季风力较大, 土壤表层风蚀作用强, 较低的地表植被覆盖度不能起到保护地表的作用, 易导致地表土壤水分损失, 沙粒含量上升[21-22]。夏季降水量占全年降水量的70%左右, 处于降水入渗补给阶段, 土壤表层含水量增加, 促进了红砂灌丛林地植被的生长, 有利于减少地表径流, 为降尘等细粒物质的截存提供了有利条件[23]。随着季节变化, 秋季降水量下降, 温度降低, 植被开始枯黄并产生枯落物, 导致地表风蚀作用加强, 土壤含水量下降, 细粒物质逐渐损失, 而出现沙粒含量累积的现象[24]。土壤酸碱度是土壤基本属性的重要指标之一。本研究中, 夏季土壤pH显著低于春季和秋季, 说明生长季植被发育过程中根系分泌的有机酸及其与微生物的相互作用降低了表层土壤的pH值[23]。
在乌拉特荒漠草原生态系统中, 土壤理化性质在随季节变化的同时, 地面节肢动物群落结构分布特征也随之发生显著变化。本研究中, 从类群组成上看, 3个季节内, 拟步甲科为共有优势类群, 蚁科在夏季亦为优势类群。这与刘继亮等[9]在黑河中游、Sackmann和Flores[25]在南美荒漠草原的研究结果相似, 即在荒漠草原生态系统中拟步甲科是地面节肢动物群落的主要类群。通过分析其个体数分布与环境因子间的关系发现, 拟步甲科动物的活动主要受到大气温度及降雨量的影响(表 3)。有研究表明, 干旱少雨的环境会对拟步甲科动物的活动和繁殖过程产生显著影响[9]。而蚁科的活动主要受到土壤含水量和酸碱度的影响(表 3)。说明土壤理化性质会直接影响蚁科的群落结构分布特征[26]。常见类群和稀有类群的数量分布特征随季节变化亦存在显著变化。说明地面节肢动物的季节分布格局受到了降雨、气温及土壤环境因子等因素的影响。不仅影响地面节肢动物的生长、发育和繁殖等生活史过程, 同时也影响到了地面节肢动物个体数量分布, 这反映了地面节肢动物对降雨、气温和土壤环境因子改变而产生的不同适应性、选择性和敏感性[6]。
生物量的大小是节肢动群落中内在所固有的功能特征之一, 也是种群数量、年龄大小、死亡率以及能值等生物指标的综合反映, 它既表示了群落的结构特征, 也反映了群落的功能(生物量)特征[27-28]。本研究中, 地面节肢动物生物量的分布格局随季节变化而发生变化。这反映了乌拉特荒漠草原地面节肢动物生物量季节分布的一种基本特征[27]。生物量超过1%的节肢动物类群中, 拟步甲科、象甲科和平腹蛛科在3个季节内均有活动分布;而粪金龟科主要活动在春/夏季, 步甲科主要活动在夏/秋季, 天牛科主要活动在夏季。研究区域内, 春季, 灌丛下积累的枯落物能够吸引食碎屑的拟步甲科前来定居, 可以有效促进枯落物的分解, 提高土壤中腐殖质的含量, 为粪金龟科昆虫的生长提供所需食物[5];夏季为植被生长季, 丰富的食物资源会吸引大量的植食性昆虫(如象甲科、天牛科等)至灌丛下取食, 随着植食性地面节肢动物类群的增加, 捕食性地面节肢动物(步甲科、平腹蛛科等)的数量也会随之增加, 从而导致灌丛下产生较多的动植物残体, 更进一步促进了粪金龟科和步甲科的生长[29];秋季植被产生新的枯落物又导致更多的食碎屑动物前来取食, 这些均有利于拟步甲科、象甲科、平腹蛛科、步甲科、粪金龟科和天牛科等维持较高的生物量水平[30]。
地面节肢动物对于环境条件季节变化具有不同的生理生态适应能力, 导致地面节肢动物在荒漠草原生态系统中的群落多样性分布存在明显的季节变化[8]。本研究中, 地面节肢动物的密度、类群数、Simpson指数和Shannon-Wiener指数均表现为夏季最高, 春季和秋季相对较低, 具有相似的季节变异规律。乌拉特荒漠草原区, 春季和秋季由于受到干旱、低温等环境条件的限制, 会直接影响地面节肢动物的繁殖、发育等生活史过程, 导致地面节肢动物的群落结构分布及其多样性受到限制[5]。夏季雨热同期, 一方面能有效促进植被群落的生长发育, 为地面节肢动物提供丰富的食物资源, 吸引更多的土壤动物进行取食活动[8];另一方面大量的地面节肢动物幼虫开始滋生繁衍, 此时地面节肢动物数量及多样性均呈显著增加的趋势[31]。
RDA分析结果表明, 优势类群和常见类群的群落结构更容易受到降雨量、气温及土壤环境因子的影响。说明地面节肢动物的序列分布与环境因子间关系密切, 即地面节肢动物的群落分布可能会受到土壤因子之间相互作用的影响[32]。通过研究相关环境因子与地面节肢动物群落结构分布的相关性, 发现不同地面节肢动物类群的分布受到气温、降雨量、土壤含水量、pH、电导率、黏粉粒含量和沙粒含量等因子的影响不同。说明不同类群地面节肢动物的个体大小、活动时间、活动方式和发育、繁殖等生物学特性存在一定的差异性, 导致地面节肢动物的个体数分布对非生物环境因子变化的响应模式不同[33-34]。
5 结论(1) 随着季节变化, 土壤TC含量、含水量、pH、黏粉粒含量和沙粒含量均具有显著差异性。(2)拟步甲科和蚁科是乌拉特荒漠草原生态系统地面节肢动物群落最重要的组成部分。(3)夏季地面节肢动物的类群数、密度和多样性等均表现为夏季显著高于春季秋季。(4)地面节肢动物的个体数分布特征受到气温、降雨量、土壤含水量、pH、电导率、黏粉粒含量和沙粒含量等环境因子的影响较大, 而受到土壤养分含量的影响较小。研究表明, 乌拉特荒漠草原红砂灌丛群落内地面节肢动物个体数、类群数和生物量随季节变化分布格局差异性较大, 夏季地面节肢动物多样性较高, 这将有助于理解乌拉特荒漠草原地面节肢动物响应气候变化的适应规律。
| [1] |
万敏, 郭永平, 郭映光, 祁俊华. 乌拉特草原植被恢复主要途径的研究. 内蒙古草业, 2004, 16(4): 49-51. |
| [2] |
王少昆, 赵学勇, 贾昆峰, 高宝兰, 曲浩, 毛伟, 连杰, 陈敏, 朱阳春. 乌拉特荒漠草原小针茅(Stipa klemenzii)群落土壤细菌多样性及垂直分布特征. 中国沙漠, 2016, 36(6): 1564-1570. |
| [3] |
朱恭, 李正平, 王万鹏, 钟芳. 红砂属植物研究进展. 甘肃林业科技, 2004, 29(3): 1-6. DOI:10.3969/j.issn.1006-0960.2004.03.001 |
| [4] |
赵哈林, 郭轶瑞, 周瑞莲. 灌丛对沙质草地土壤结皮形成发育的影响及其作用机制. 中国沙漠, 2011, 31(5): 1105-1111. |
| [5] |
刘任涛, 郗伟华, 朱凡. 宁夏荒漠草原地面节肢动物群落组成及季节动态特征. 草业学报, 2016, 25(6): 126-135. |
| [6] |
刘任涛, 赵哈林, 赵学勇. 半干旱区草地土壤动物多样性的季节变化及其与温湿度的关系. 干旱区资源与环境, 2013, 27(1): 97-101. |
| [7] |
关宏斌, 郭砺, 刘永江. 科尔沁沙地土壤动物垂直分布、季节动态及群落多样性探讨. 中国沙漠, 1999, 19(S1): 111-115. |
| [8] |
刘继亮, 赵文智, 李锋瑞. 黑河中游沙质荒漠地面节肢动物群落组成及多样性季节变异. 干旱区资源与环境, 2015, 29(6): 98-103. |
| [9] |
刘继亮, 李锋瑞, 刘七军, 牛瑞雪. 黑河中游干旱荒漠地面节肢动物群落季节变异规律. 草业学报, 2010, 19(5): 161-169. |
| [10] |
曲浩, 赵学勇, 王少昆, 黄文达, 毛伟. 乌拉特荒漠草原不同植被群落对土壤碳、氮的影响. 草业科学, 2014, 31(3): 355-360. |
| [11] |
刘任涛, 郗伟华, 赵娟, 刘佳楠. 不同立地条件柠条灌丛内外地面节肢动物群落结构分布特征. 干旱区研究, 2018, 35(2): 354-362. |
| [12] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [13] |
郑乐怡, 归鸿. 昆虫分类. 南京: 南京师范大学出版社, 1999.
|
| [14] |
王新谱, 杨贵军. 宁夏贺兰山昆虫. 宁夏: 宁夏人民出版社, 2010.
|
| [15] |
张雪萍, 崔国发, 陈鹏. 人工落叶松林土壤动物生物量的研究. 应用生态学报, 1996, 7(2): 150-154. |
| [16] |
刘任涛, 郗伟华, 刘佳楠, 赵娟, 常海涛. 沙地柠条(Caragana)灌丛微生境节肢动物群落特征. 中国沙漠, 2018, 38(1): 117-125. |
| [17] |
Su Y Z, Zhao H L, Zhao W Z, Zhang T H. Fractal features of soil particle size distribution and the implication for indicating desertification. Geoderma, 2004, 122(1): 43-49. DOI:10.1016/j.geoderma.2003.12.003 |
| [18] |
Liu J L, Zhao W Z, Li F R. The effects of the identity of shrub species on the distribution and diversity of ground arthropods in a sandy desert ecosystem of northwestern China. Sciences in Cold and Arid Regions, 2014, 6(6): 587-596. |
| [19] |
刘任涛. 荒漠草原土壤动物与降雨关系研究现状. 生态学杂志, 2012, 31(3): 760-765. |
| [20] |
吴永胜, 马万里, 李浩, 吕桂芬, 卢萍. 内蒙古退化荒漠草原土壤有机碳和微生物生物量碳含量的季节变化. 应用生态学报, 2010, 21(2): 312-316. |
| [21] |
高英志, 汪诗平, 韩兴国, 陈全胜, 王艳芬, 周志勇, 张淑敏, 杨晶. 退化草地恢复过程中土壤氮素状况以及与植被地上绿色生物量形成关系的研究. 植物生态学报, 2004, 28(3): 285-293. DOI:10.3321/j.issn:1005-264X.2004.03.001 |
| [22] |
张蕊, 赵学勇, 左小安, 刘新平, 曲浩, 马旭君, 刘良旭, 陈娟丽, 刘立平. 荒漠草原沙生针茅(Stipa glareosa)群落物种多样性和地上生物量对降雨量的响应. 中国沙漠, 2019, 39(2): 45-52. |
| [23] |
常海涛, 刘任涛, 刘佳楠, 赵娟, 罗雅曦, 张静, 马继. 草方格造林固沙过程中土壤性质变化及分形特征——以腾格里沙漠东南缘为例. 水土保持学报, 2018, 32(6): 58-65, 165-165. |
| [24] |
Su Y Z, Zhang T H, Li Y L, Wang F. Changes in soil properties after establishment of Artemisia halodendron and Caragana microphylla on shifting sand dunes in semiarid Horqin Sandy Land, northern China. Environmental Management, 2005, 36(2): 272-281. DOI:10.1007/s00267-004-4083-x |
| [25] |
Sackmann P, Flores G E. Temporal and spatial patterns of tenebrionid beetle diversity in NW Patagonia, Argentina. Journal of Arid Environments, 2009, 73(12): 1095-1102. DOI:10.1016/j.jaridenv.2009.05.007 |
| [26] |
刘任涛, 赵哈林, 赵学勇. 科尔沁沙地流动沙丘掘穴蚁蚁丘分布及影响因素. 应用生态学报, 2009, 20(2): 376-380. |
| [27] |
师光禄, 席银宝, 王海香, 李捷, 赵飞. 枣园节肢动物群落的数量与生物量多样性特征分析. 林业科学, 2004, 40(2): 107-112. |
| [28] |
Yang Y, Liu B R. Impact factors and relationships between biodiversity and biomass of different communities in the desert steppe of Ningxia, China. Acta Prataculturae Sinica, 2015, 24(10): 48-57. |
| [29] |
李婷, 苏杰, 许照强, 李兴龙, 韩国栋, 张建萍. 古尔班通古特沙漠梭梭林象甲科昆虫分布及其动态研究. 石河子大学学报:自然科学版, 2017, 35(2): 201-206. |
| [30] |
Li F R, Liu J L, Liu C A, Liu Q J, Niu R X. Shrubs and species identity effects on the distribution and diversity of ground-dwelling arthropods in a Gobi desert. Journal of Insect Conservation, 2013, 17(2): 319-331. DOI:10.1007/s10841-012-9512-1 |
| [31] |
武海涛, 吕宪国, 姜明, 朱宝光. 三江平原典型湿地土壤动物群落结构及季节变化. 湿地科学, 2008, 6(4): 459-465. |
| [32] |
刘继亮, 殷秀琴, 邱丽丽. 左家自然保护区大型土壤动物与土壤因子关系研究. 土壤学报, 2008, 45(1): 130-136. DOI:10.3321/j.issn:0564-3929.2008.01.017 |
| [33] |
Seastedt T R, Crossley D A Jr. The influence of arthropods on ecosystems. BioScience, 1984, 34(3): 157-161. DOI:10.2307/1309750 |
| [34] |
任国栋, 于有志, 侯文君. 中国荒漠半荒漠地区拟步甲的组成和分布特点. 河北大学学报:自然科学版, 1999, 19(2): 176-183. |