 2020, Vol. 40
2020, Vol. 40文章信息
- 殷正, 范秀华
- YIN Zheng, FAN Xiuhua
- 长白山不同演替阶段温带森林林下草本植物对乔木幼苗的影响
- Effects of herbs on tree seedlings in different succession stages of temperate forests in Changbai Mountain, China
- 生态学报. 2020, 40(7): 2194-2204
- Acta Ecologica Sinica. 2020, 40(7): 2194-2204
- http://dx.doi.org/10.5846/stxb201901180148
-
文章历史
- 收稿日期: 2019-01-18
- 网络出版日期: 2019-12-26

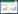




森林更新是森林群落动态的重要组成部分, 也是森林生态系统自我繁衍恢复的重要手段[1-2]。幼苗的生长、发育和定居作为植物整个生活史中最脆弱、对外界环境最敏感的时期[3-4], 会在时空上发生明显变化并间接影响森林群落的结构和稳定性, 对植物群落的更新起着至关重要的作用。对森林更新的研究有助于我们深入了解内部机制, 完善已有的森林生态系统经营的方式[5-6]。
林下幼苗更新的研究是国内外森林生态学研究的重要内容, 对幼苗更新影响因素的分析更是得到广泛关注。影响幼苗存活和更新的因素主要可分为非生物因素和生物因素两大类。非生物因素主要是指光照、水分、土壤、温度、自然灾害等[7-9];生物因素主要是指幼苗自身植物属性、邻体、植食性动物及有害微生物等[10-12]。目前国内学者对幼苗更新的研究多集中在对幼苗的长期监测, 对幼苗的数量组成和空间格局[13-15]、月际动态和年际动态[16-17]、幼苗存活影响因素[18]等方面进行探讨。其中在对幼苗存活影响因素的研究中关注的生物因素主要为树木邻体[19], 而草本植物对更新早期幼苗也存在着相互作用, 在不同生长条件下草本植物对乔木幼苗生长的影响也不同, 目前关于草本植物对更新幼苗影响的研究存在多种结论。有研究表明, 物种丰富的林下草本层更有利于森林的恢复[20];对华山松人工林天然更新的研究中发现灌草层盖度对幼苗数量起正相互作用, 认为较大的灌草盖度易形成适宜的小气候环境[21];在分析了幼苗密度与林下植被之间的关系后发现大多数物种与幼苗密度之间呈正相关关系[22], 而Caldeira等发现幼苗存活率与草本植物生物量间表现出负相关[23]。以上研究在温带森林进行的有限, 需要进一步探究不同演替阶段温带森林林下草本植物对幼苗生长及存活的影响以及产生这种影响可能的机制。
长白山覆盖着大面积温带原始森林, 过去是重要木材产地, 但受20世纪初以来大规模采伐的影响, 大面积原始阔叶红松林被次生林或人工林所取代, 次生针阔混交林与原始阔叶红松林在物种组成、种群结构上均存在着较大差异, 主导幼苗生长与存活的因子也可能不同。有关长白山不同演替阶段幼苗更新的研究已有一些[8, 15, 24], 而草本植物对乔木更新幼苗直接作用的研究较少。因此, 本文以长白山阔叶红松林和次生针阔混交林固定监测样地为平台, 将草本植物对乔木幼苗的直接影响加以分析, 旨在通过探讨:(1)探讨草本植物对不同演替阶段下乔木幼苗高度生长的影响;(2)探讨草本植物对不同演替阶段下乔木幼苗生长的影响在不同年龄级是否存在差异;(3)探讨幼苗存活率与草本植物盖度、多度和草本物种数量之间的相关性, 揭示草本植物与乔木幼苗在不同演替阶段温带森林中的相互关系, 为温带森林多样性维持和可持续经营提供理论依据和参考。
1 研究区概况研究区位于吉林省白河林业局光明林场(次生针阔混交林样地)和长白山北坡自然保护区(阔叶红松林样地)(图 1)。该地区气候类型为受季风影响的温带大陆性山地气候, 年均气温3.6℃, 年均降水量707 mm, 最热月7月平均气温19.6℃, 最冷月1月平均气温-15.4℃, 土壤类型为山地暗棕色森林土, 土层厚20—100 cm。次生针阔混交林是由阔叶红松林采伐后形成, 林龄60—80年, 林下生境均一稳定, 群落中木本植物主要包括紫椴(Tilia amurensis)、臭松(Abies nephrolepis)、假色槭(Acer pseudo-sieboldianum)、色木槭(Acer mono)、白桦(Betula platyphylla)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、大青杨(Populus ussuriensis)等[24]。阔叶红松林为原始林, 人为干扰少, 环境均质稳定, 其中主要木本植物包括红松(Pinus koraiensis)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、蒙古栎(Quercus mongolica)、色木槭(Acer mono)等[25]。

|
| 图 1 研究区位置与样地分布 Fig. 1 Location of study area and distribution of two plots Ⅰ:次生针阔混交林样地;Ⅱ:阔叶红松林样地 |
参照美国CTFS (Center for Tropical Forest Science)样地建设标准和操作规范, 于2005年和2014年分别选择典型的次生针阔混交林和阔叶红松林建立固定监测样地。次生针阔混交林样地面积5.2 hm2(260 m×200 m), 样地坐标42°20′N, 128°07′E, 平均海拔748 m。阔叶红松林样地面积40 hm2(800 m×500 m), 样地坐标42°20′N, 127°54′E, 平均海拔997 m。将每个样地划分为20 m×20 m的连续样方。
2.2 幼苗、草本植物调查及除草处理2018年, 在次生针阔混交林样地和阔叶红松林样地内已有长期固定幼苗监测样方的基础上分别设置72组和51组1 m×1 m样方(包括对照组(CK)和处理组(HR)), 共计246个, 样方设置如图 2所示。6月, 对样方内所有胸径(DBH, Diameter of Breast Height)小于1 cm的乔木个体进行调查, 记录幼苗种类、高度、数量、年龄等并挂牌标记, 同时调查样方中草本植物物种、数量、盖度。在草本植物调查完毕后去除处理组样方中所有草本植物, 为避免拔除草本时影响乔木幼苗, 较大草本使用工具从根部剪除, 较小草本直接拔除[26], 去除草本过程中保证其他条件不变。8月再次进行幼苗调查, 两次调查期间无新生乔木幼苗, 及时去除样方中新生草本植物。

|
| 图 2 幼苗样方设置图 Fig. 2 Seedling quadrats layout at two forest type CK:对照Control;HR:草本去除Herb removal |
(1) 存活率=第二次调查到的非新生幼苗个体数/第一次调查幼苗总个体数
(2) 幼苗密度=幼苗个体数/样方面积(m2)
2.3.2 土壤含水量2018年8月第二次进行幼苗调查前, 使用手持土壤水分测定仪(HH2 Moisture Meter, UK)测定幼苗样方对角线上3个点的表层土壤含水率, 取三点平均值作为该幼苗样方土壤含水量(表 1)。
| 林型Forest types | 对照组CK | 处理组HR |
| 次生针阔混交林Secondary conifer and broad-leaved mixed forests | 18.89±12.61a | 18.00±11.45a |
| 阔叶红松林Broad-leaved Korean pine forests | 22.95±11.62b | 23.73±11.71b |
| CK:对照Control;HR:草本去除Herb removal;表中不同字母代表处理与对照间差异显著(P < 0.05) | ||
采用246个1 m×1 m样方中两次幼苗调查数据, 草本植物调查数据以及土壤含水量指标进行数据分析。用Wilcoxon秩和检验(Mann-Whitney U检验)比较在次生针阔混交林和阔叶红松林中群落水平和不同年龄级水平下去除草本对幼苗高度生长的影响。利用普通最小二乘(OLS)回归法分析幼苗存活率与草本植物盖度, 多度和物种数之间的相关关系。本文中所有数据分析和制图均采用Microsoft Excel 2016和国际通用软件R3.5.1完成。
3 结果与分析 3.1 林下草本组成及幼苗数量特征2018年6月在次生针阔混交林样方内共调查到63种草本植物, 隶属于31科57属, 主要包括唇形科、酢浆草科、莎草科、五福花科、紫草科等;阔叶红松林样方内共调查到62种草本植物, 分属30科55属, 主要包括伞形科、凤仙花科、唇形科、茜草科、百合科等。2018年6月和8月两次调查期间, 次生针阔混交林样方内共调查到13种乔木幼苗, 隶属于7科8属;阔叶红松林样方内共调查到11种乔木幼苗, 隶属于5科5属(表 2)。两个演替阶段森林中幼苗都是以水曲柳、假色槭为主, 水曲柳幼苗在两个林分下都具有明显优势地位;簇毛槭、青楷槭和紫椴幼苗在次生针阔混交林中出现较多, 而红松幼苗在阔叶红松林中出现更多。
| 林型 Forest type |
物种 Species |
幼苗数量 No. of seedlings |
出现样方数 No. of stations occupied |
幼苗密度 Seedling density(株/m2) |
||||||||||||||
| 对照组 Control |
处理组 Herb moval |
对照组 Control |
处理组 Herb moval |
对照组 Control |
处理组 Herb moval |
|||||||||||||
| 6月 June |
8月 August |
6月 June |
8月 August |
6月 June |
8月 August |
6月 June |
8月 August |
6月 June |
8月 August |
6月 June |
8月 August |
|||||||
| 次生针阔混交林 | 春榆Ulmus davidiana var. japonica | 1 | 0 | 3 | 3 | 1 | 0 | 2 | 2 | 1.000 | 0.000 | 1.500 | 1.500 | |||||
| Secondary conifer and | 簇毛槭Acer barbinerve | 28 | 15 | 21 | 10 | 15 | 11 | 14 | 5 | 1.867 | 1.364 | 1.500 | 2.000 | |||||
| broad-leaved mixed | 红松Pinus koraiensis | 2 | 1 | 4 | 2 | 2 | 1 | 2 | 2 | 1.000 | 1.000 | 2.000 | 1.000 | |||||
| forests | 怀槐Maackia amurensis | 12 | 10 | 10 | 8 | 8 | 7 | 6 | 6 | 1.500 | 1.429 | 1.667 | 1.333 | |||||
| 黄檗Phellodendron amurense | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1.000 | 1.000 | 0.000 | 0.000 | ||||||
| 假色槭Acer pseudosieboldianum | 36 | 23 | 33 | 26 | 16 | 15 | 9 | 6 | 2.250 | 1.533 | 3.667 | 4.333 | ||||||
| 蒙古栎Quercus mongolica | 6 | 3 | 4 | 2 | 6 | 3 | 4 | 2 | 1.000 | 1.000 | 1.000 | 1.000 | ||||||
| 青楷槭Acer tegmentosum | 22 | 18 | 26 | 21 | 12 | 11 | 11 | 11 | 1.833 | 1.636 | 2.364 | 1.909 | ||||||
| 色木槭Acer mono | 8 | 7 | 14 | 8 | 7 | 6 | 12 | 6 | 1.143 | 1.167 | 1.167 | 1.333 | ||||||
| 水曲柳Fraxinus mandschurica | 348 | 213 | 422 | 256 | 48 | 41 | 52 | 45 | 7.250 | 5.195 | 8.115 | 5.689 | ||||||
| 小楷槭Acer komarovii | 7 | 6 | 4 | 3 | 6 | 4 | 4 | 3 | 1.167 | 1.500 | 1.000 | 1.000 | ||||||
| 紫椴Tilia amurensis | 32 | 10 | 22 | 10 | 18 | 7 | 15 | 9 | 1.778 | 1.429 | 1.467 | 1.111 | ||||||
| 臭松Abies nephrolepis | 0 | 0 | 2 | 2 | 0 | 0 | 2 | 2 | 0.000 | 0.000 | 1.000 | 1.000 | ||||||
| 总计Total | 503 | 307 | 565 | 351 | 22.787 | 18.252 | 26.446 | 23.209 | ||||||||||
| 阔叶红松林 | 白牛槭Acer mandshuricum | 1 | 1 | 3 | 3 | 1 | 1 | 3 | 3 | 1.000 | 1.000 | 1.000 | 1.000 | |||||
| Broad-leaved Korean | 簇毛槭Acer barbinerve | 10 | 6 | 12 | 9 | 4 | 2 | 6 | 6 | 2.500 | 3.000 | 2.000 | 1.500 | |||||
| pine forests | 风桦Betula costata | 4 | 2 | 2 | 2 | 4 | 2 | 2 | 2 | 1.000 | 1.000 | 1.000 | 1.000 | |||||
| 红松Pinus koraiensis | 16 | 16 | 5 | 4 | 3 | 3 | 3 | 2 | 5.333 | 5.333 | 1.667 | 2.000 | ||||||
| 假色槭Acer pseudosieboldianum | 20 | 14 | 17 | 12 | 13 | 11 | 12 | 10 | 1.538 | 1.273 | 1.417 | 1.200 | ||||||
| 蒙古栎Quercus mongolica | 2 | 2 | 2 | 1 | 1 | 1 | 2 | 1 | 2.000 | 2.000 | 1.000 | 1.000 | ||||||
| 青楷槭Acer tegmentosum | 6 | 5 | 4 | 3 | 3 | 2 | 2 | 2 | 2.000 | 2.500 | 2.000 | 1.500 | ||||||
| 色木槭Acer mono | 1 | 1 | 11 | 6 | 1 | 1 | 3 | 3 | 1.000 | 1.000 | 3.667 | 2.000 | ||||||
| 水曲柳Fraxinus mandschurica | 441 | 286 | 486 | 319 | 44 | 40 | 43 | 40 | 10.023 | 7.150 | 11.302 | 7.975 | ||||||
| 小楷槭Acer komarovii | 4 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2.000 | 1.500 | 1.000 | 1.000 | ||||||
| 紫椴Tilia amurensis | 12 | 6 | 12 | 9 | 10 | 5 | 10 | 8 | 1.200 | 1.200 | 1.200 | 1.125 | ||||||
| 总计Total | 517 | 342 | 556 | 370 | 29.595 | 26.956 | 27.252 | 21.300 | ||||||||||
由图 3可知, 在次生针阔混交林(Ⅰ)和阔叶红松林(Ⅱ)下HR处理对幼苗高度生长的影响不同。在次生针阔混交林中, HR处理前后均显示CK和HR组高度存在显著差异, 而HR处理后幼苗高度增长量显著高于CK。在阔叶红松林中, HR处理前组间高度无显著差异, HR处理后幼苗高度显著高于CK并且处理组幼苗高度增长量显著高于CK。比较两演替阶段幼苗高度生长对HR处理的响应, 阔叶红松林中幼苗高度增长显著高于次生针阔混交林。这表明在在两种林型中HR处理在群落水平上均表现出有助于幼苗的高度生长;且在阔叶红松林中幼苗高度生长显著高于次生针阔混交林。

|
| 图 3 除草处理下不同演替阶段森林林下幼苗高度生长比较 Fig. 3 Comparison of seedling height growth in different succession stages under herb removal treatment SCBF:次生针阔混交林Secondary conifer and broad-leaved mixed forests;BKF:阔叶红松林Broad-leaved Korean pine forests;图中不同字母代表处理与对照间差异显著(P < 0.05) |
由图 4可知, 在次生针阔混交林中HR处理对一到三年生幼苗的高度生长影响不显著, CK与HR在处理前后相对高度未产生显著变化;与CK相比, HR处理对幼苗高度生长量未产生显著影响。四年生及以上幼苗在HR处理前两组高度无显著差异, HR处理后高度较CK出现显著差异;HR处理后的幼苗高度增长量也显著高于CK。这表明在次生针阔混交林中, 草本植物对一到三年生幼苗高度生长的影响较小, 除草并未对幼苗高度生长产生显著影响;多年生幼苗对HR处理有较强响应, 除草后高度生长显著加快, HR处理有助于多年生幼苗的生长。

|
| 图 4 除草处理下次生针阔混交林下不同年龄级水平幼苗高度生长比较 Fig. 4 Comparison of height growth of seedlings at different age classes under herb removal treatment in secondary coniferous and broad-leaved mixed forests |
由图 5可知, 在阔叶红松林中HR处理对一至三年生幼苗产生显著影响, CK和HR处理组前后高度都未出现差异, 但HR处理后幼苗的高度增长量显著高于CK;相对于一至三年生幼苗, 阔叶红松林中多年生幼苗对HR处理响应较弱, HR处理后高度较CK未产生显著差异, 高度增长量也未有显著差异。这表明在阔叶红松林中, 去除草本植物有助于一至三年生幼苗高度生长;HR处理对多年生幼苗生长产生的影响有限, 未达显著水平。

|
| 图 5 除草处理阔叶红松林下不同年龄级水平幼苗高度生长比较 Fig. 5 Comparison of height growth of seedlings at different age classes under herb removal treatment in broad-leaved Korean pine forests |
将在次生针阔混交林和阔叶红松林中都占据优势的水曲柳和假色槭幼苗在HR处理前后的高度生长量进行比较。由图 6, 占据绝对优势的水曲柳幼苗在两种林型下HR处理后高度生长量较CK均显著升高, 且在阔叶红松林中HR对水曲柳高度生长的作用更为显著;对于假色槭, HR处理后只在次生针阔混交林中高度生长量显著地高于CK, 在阔叶红松林中HR处理组与CK未发现显著差异。其他幼苗在次生针阔混交林中HR处理对高度生长未产生影响, 在阔叶红松林中HR处理显著促进了其他幼苗的高度生长。

|
| 图 6 除草处理下不同演替阶段森林林下不同物种幼苗高度生长量比较 Fig. 6 Comparison of seedling height growth of different species in different succession stages under herb removal treatment |
对次生针阔混交林和阔叶红松林中处理与对照组在试验期间的幼苗存活率进行对比, 两群落中处理与对照组间幼苗存活率均未产生显著差异(P > 0.05);且次生针阔混交林和阔叶红松林中处理组与对照组幼苗存活率均趋于一致(图 7)。

|
| 图 7 不同群落幼苗存活率比较 Fig. 7 Comparison of seedlings survival in different communities |
由图 8, 幼苗存活率与草本植物各项指标之间相关关系不同, 次生针阔混交林中幼苗存活率随着草本植物多度和草本植物物种数的升高而升高, 存在着显著正相关关系, 而幼苗存活率与草本盖度之间不存在相关关系。这表明, 随着草本植物多度和物种数的增加有助于次生针阔混交林下幼苗的存活, 与草本盖度无直接关系。

|
| 图 8 次生针阔混交林中幼苗存活率与草本植物间关系 Fig. 8 Relationship between seedling survival and herbaceous plants in secondary coniferous and broad-leaved mixed forests |
与次生针阔混交林相比, 阔叶红松林中幼苗存活率与草本植物多度之间未发现显著相关关系;幼苗存活率与草本盖度和草本物种数之间的关系与次生针阔混交林中结果一致(图 9)。其中, 幼苗存活率与草本盖度之间未发现显著相关关系, 幼苗存活率与草本物种之间存在着显著的正相关关系。这表明, 草本物种数在两种林型中都表现出了对幼苗存活率的显著正相关关系, 较高的草本物种数更有助于幼苗的存活;而对于草本植物盖度在两种林型中都未表现出和幼苗存活率的相关关系。

|
| 图 9 阔叶红松林中幼苗存活率与草本植物间关系 Fig. 9 Relationship between seedling survival and herbaceous plants in broad-leaved Korean pine forests |
幼苗作为整个森林更新过程中最脆弱、变化最大的阶段, 对更新成功以及群落未来组成具有重要作用, 因而受到人们的广泛关注[3]。在2018年6月和8月对长白山不同演替阶段温带森林样地内246个幼苗样方的调查分析发现, 次生针阔混交林和阔叶红松林中幼苗物种都是群落中乔木层主要物种, 与样地中乔木大树物种具有一定的一致性[24]。其中, 在两个演替阶段温带森林中水曲柳幼苗几乎均占据调查幼苗总体的80%以上, 占据优势地位, 这可能与乔木树种在不同年份间种子雨强度有很大差异有关, 如刘双等[27]研究发现, 一些树种如水曲柳在某一年份结实量很小或几乎不结实, 而红松受人为干扰较大。
4.2 草本植物对幼苗高度生长的影响在乔木幼苗早期生长阶段, 草本植物作为生物邻体与幼苗生长有着密切关系, 为探讨不同演替阶段林下草本植物对乔木幼苗的影响, 本试验通过样方中草本植物去除的对照试验表明在群落水平上次生针阔混交林与阔叶红松林下幼苗高度生长受到草本植物的抑制作用。一些研究也得出了类似的结论, Denslow等[28]的研究结果表明, 草本植物的存在在大多数情况下会显著降低林下幼苗更新, 去除草本植物后幼苗的发生显著升高;韩文娟等[29]通过野外播种和室内控制试验分析去除草本对幼苗萌发和生长的研究发现, 清除草本有助于提高种子萌发;Duncan等[26]在灌木和其他地上植被对树木更新影响的研究中表明, 去除地上植被促进了幼苗高度生长。这可能是由于乔木幼苗阶段对环境的变化反应敏感, 去除草本植物后获得了更好的环境条件, 更有利于幼苗生长。也有研究得出的结果不同, Masaki等[30]的研究中发现, 幼苗生长与草本盖度呈正相关关系, 这可能与该实验研究地点中林隙条件和物种的生长特性有关。去除草本植物后, HR处理与CK土壤水分未发生显著变化(表 1), 表明去除草本并非通过改变土壤水分影响幼苗生长, 可能是草本去除后对光照条件有所提升导致的。阔叶红松林中幼苗的高度生长量在对照和处理组都显著高于次生针阔混交林, 这可能是因为阔叶红松林处于演替的后期阶段, 林分结构相对稳定, 且土壤含水量在阔叶红松林中也显著高于次生针阔混交林(表 1), 相对较高的土壤含水量有助于幼苗的生长。
此外, 去除草本植物对不同年龄级幼苗高度生长产生的影响也不同。在次生针阔混交林中草本去除对一至三年生幼苗高度生长的影响较小, 对四年生及以上幼苗影响较大, 这与孟令君等[31]研究发现邻体作用对当年生幼苗存活影响更加显著的结果不同, 这可能是由于本试验所在的次生针阔混交林样地中草本植物大多较为低矮, 该次生林中生境因子较生物邻体对幼苗影响更大, 去除处理未对一至三年生幼苗生长环境产生显著影响;而对于多年生幼苗, 其受到来自草本植物对土壤养分等生境因子的竞争更为强烈, 草本去除有助于释放有限资源, 利于其高度生长;阔叶红松林中草本去除对一至三年生幼苗影响显著, 对四年生及以上幼苗影响较小, 可能的原因是阔叶红松林样方中草本更为密集和高大, 去除草本极大的提升了一至三年生幼苗的生长条件, 且阔叶红松林样地中调查到多年生幼苗数量较少且树龄较大, 枝干木质化程度增加, 对病原体、捕食者或非生物胁迫作用的耐受能力增强, 受到草本邻体的影响较小, 且随着幼苗年龄增大, 高度生长也逐渐减弱[30]。
去除草本对乔木幼苗高度生长的影响存在着明显的种间差异。在物种水平上, 水曲柳幼苗对去除草本植物的响应在次生针阔混交林和阔叶红松林中相似, 草本去除均显著增加了水曲柳幼苗的高度生长;此外在阔叶红松林中表现出显著高于次生针阔混交林的高度增长量, 这与群落水平上得出的结果相似, 韩有志等[32]对水曲柳与林分光照关系的研究中发现的水曲柳更新与光照空间异质性紧密相关可能是其中一个原因, 草本去除后对光环境的改善有助于水曲柳幼苗生长。假色槭幼苗对去除草本的响应在不同演替阶段森林中不同, 在次生针阔混交林中去除草本对假色槭幼苗产生显著影响, 而在阔叶红松林中假色槭幼苗在除草处理前后未表现出显著差异, 这是由于阔叶红松林中假色槭幼苗年龄较大, 除草并未影响其高度生长[30]。在对其他幼苗的分析发现, 其他幼苗只在阔叶红松林中对除草处理产生响应, 去草有助于幼苗高度生长, 这可能与次生针阔混交林中草本植物低矮和其他幼苗的物种组成有关。
4.3 草本植物对幼苗存活率的影响样方处理组与对照组间幼苗存活率在次生针阔混交林和阔叶红松林中差异均不显著, 表明去除草本植物对幼苗存活未产生显著影响, 这也可能与当地2018年7月少有的长达一个月的高温干旱有关, 长时间降水不足与高温使得水分成为限制幼苗存活的主要因素, 草本生物邻体对幼苗存活的影响成为次要因素。
草本植物与乔木幼苗存活率间关系在次生针阔混交林和阔叶红松林中存在差异。次生针阔混交林中幼苗存活率与草本多度以及草本物种数呈正相关关系, 这与Thomaes等的研究结果一致, 认为在森林恢复过程中较高的草本层物种多样性有助于减少竞争性排斥对主要树种幼苗的影响[20], 高的草本多度与物种多样性在干旱等胁迫条件下有助于维持相对稳定的微生境[33]。幼苗存活率与草本盖度之间未发现显著相关关系, 这可能是由于地表光照条件基本能满足幼苗的生长存活需求, 草本植物盖度对光照条件的影响并非制约幼苗存活的主导因素[34]。阔叶红松林中幼苗存活率只和草本物种数存在正相关关系, 与草本多度以及草本盖度无显著相关关系, 这可能是由于阔叶红松林中调查到的幼苗多为一年生, 多年生幼苗数量较少[35], 而水分是幼苗存活主导因素, 高的草本多样性在一定程度上有助于在干旱条件下保持土壤水分, 且草本植物多样性高代表着良好的生境条件, 因此幼苗存活率随草本多样性升高。也有研究表明, 草本植物对乔木幼苗间的竞争不利于幼苗自然更新, 当草本盖度较低时, 幼苗更新更为良好[36];Van等[37]在灌草对胖大海幼树幼苗影响的研究中表明, 草本盖度增加不利于胖大海幼树幼苗的更新;Tsvuura等[38]的研究发现, 草本植物主要通过对光照条件的削弱抑制了乔木幼苗的生长与存活。
5 结论在次生针阔混交林和阔叶红松林中, 草本植物均对乔木幼苗生长存活产生影响, 去除草本促进了幼苗的高度生长, 进而促进了森林更新;去除草本对幼苗生长的影响在年龄级间和物种间存在差异, 随着幼苗年龄增加, 生境因素的影响加大, 草本邻体对幼苗影响减弱;草本植物可能在一定程度上有助于改善微生境条件, 但未显著促进幼苗存活率的提高。
| [1] |
徐振邦, 代力民, 陈吉泉, 王战, 戴洪才, 李昕. 长白山红松阔叶混交林森林天然更新条件的研究. 生态学报, 2001, 21(9): 1413-1420. DOI:10.3321/j.issn:1000-0933.2001.09.003 |
| [2] |
Harms K E, Wright S J, Calderón O, Hernández A, Herre E A. Pervasive density-dependent recruitment enhances seedling diversity in a tropical forest. Nature, 2000, 404(6777): 493-495. DOI:10.1038/35006630 |
| [3] |
Clark D A, Clark D B. Spacing dynamics of a tropical rain forest tree:evaluation of the Janzen-Connell model. The American Naturalist, 1984, 124(6): 769-788. DOI:10.1086/284316 |
| [4] |
Wright S J, Muller-Landau H C, Calderón O, Hernandéz A. Annual and spatial variation in seedfall and seedling recruitment in a neotropical forest. Ecology, 2005, 86(4): 848-860. DOI:10.1890/03-0750 |
| [5] |
彭闪江, 黄忠良, 彭少麟, 欧阳学军, 徐国良. 植物天然更新过程中种子和幼苗死亡的影响因素. 广西植物, 2004, 24(2): 113-121, 124-124. DOI:10.3969/j.issn.1000-3142.2004.02.004 |
| [6] |
Muller-Landau H C, Wright S J, Calderón O, Condit R, Hubbell S P. Interspecific variation in primary seed dispersal in a tropical forest. Journal of Ecology, 2008, 96(4): 653-667. DOI:10.1111/j.1365-2745.2008.01399.x |
| [7] |
Padilla F M, Pugnaire F I. Rooting depth and soil moisture control Mediterranean woody seedling survival during drought. Functional Ecology, 2007, 21(3): 489-495. DOI:10.1111/j.1365-2435.2007.01267.x |
| [8] |
刘帅, 廖嘉星, 肖翠, 范秀华. 长白山次生针阔混交林乔木幼苗存活的影响因素分析. 植物生态学报, 2016, 40(7): 711-722. |
| [9] |
Johnstone J F, Chapin III F S. Effects of soil burn severity on post-fire tree recruitment in boreal forest. Ecosystems, 2006, 9(1): 14-31. DOI:10.1007/s10021-004-0042-x |
| [10] |
Klopčič M, Simončič T, Bončina A. Comparison of regeneration and recruitment of shade-tolerant and light-demanding tree species in mixed uneven-aged forests:experiences from the Dinaric region. Forestry, 2015, 88(5): 552-563. DOI:10.1093/forestry/cpv021 |
| [11] |
Comita L S, Hubbell S P. Local neighborhood and species' shade tolerance influence survival in a diverse seedling bank. Ecology, 2009, 90(2): 328-334. DOI:10.1890/08-0451.1 |
| [12] |
Jansen P A, Visser M D, Joseph Wright S, Rutten G, Muller-Landau H C. Negative density dependence of seed dispersal and seedling recruitment in a Neotropical palm. Ecology Letters, 2014, 17(9): 1111-1120. DOI:10.1111/ele.12317 |
| [13] |
李晓亮, 王洪, 郑征, 林露湘, 邓晓保, 曹敏. 西双版纳热带森林树种幼苗的组成、空间分布和旱季存活. 植物生态学报, 2009, 33(4): 658-671. DOI:10.3773/j.issn.1005-264x.2009.04.004 |
| [14] |
宾粤, 叶万辉, 曹洪麟, 黄忠良, 练琚愉. 鼎湖山南亚热带常绿阔叶林20公顷样地幼苗的分布. 生物多样性, 2011, 19(2): 127-133. |
| [15] |
陈贝贝, 王凯, 倪瑞强, 程艳霞. 长白山针阔混交林乔木幼苗组成与空间分布. 北京林业大学学报, 2018, 40(2): 68-75. |
| [16] |
姚杰, 闫琰, 张春雨, 邳田辉, 赵秀海. 吉林蛟河针阔混交林乔木幼苗组成与月际动态. 植物生态学报, 2015, 39(7): 717-725. |
| [17] |
闫琰, 姚杰, 张新娜, 张春雨, 赵秀海. 吉林蛟河不同演替阶段针阔混交林木本植物幼苗空间分布与年际动态. 生态学报, 2016, 36(23): 7644-7654. |
| [18] |
刘帅, 侯嫚嫚, 廖嘉星, 王均伟, 肖翠, 范秀华. 生物邻体和生境异质性共同驱动乔木幼苗存活动态. 应用与环境生物学报, 2016, 22(4): 639-645. |
| [19] |
肖翠, 刘帅, 黄珍, 樊莹, 王均伟, 赵秀海, 唐景毅. 长白山阔叶红松林中影响乔木幼苗存活的关键因素. 生态学报, 2015, 35(19): 6557-6565. |
| [20] |
Thomaes A, De Keersmaeker L, Verschelde P, Vandekerkhove K, Verheyen K. Tree species determine the colonisation success of forest herbs in post-agricultural forests:Results from a 9 yr introduction experiment. Biological Conservation, 2014, 169: 238-247. DOI:10.1016/j.biocon.2013.10.024 |
| [21] |
张小鹏, 王得祥, 杜江涛, 张宋智. 微生境对华山松人工林天然更新的影响. 西北农林科技大学学报:自然科学版, 2018, 46(1): 39-45. |
| [22] |
Dyderski M K, Gazda A, Hachułka M, Horodecki P, Kałucka I L, Kamczyc J, Malicki M, Pielech R, Smoczyk M, Skorupski M, Wierzcholska S, Jagodziński A M. Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests. Annals of Forest Science, 2018, 75(4): 91. DOI:10.1007/s13595-018-0775-x |
| [23] |
Caldeira M C, Ibáñez I, Nogueira C, Bugalho M N, Lecomte X, Moreira A, Pereira J S. Direct and indirect effects of tree canopy facilitation in the recruitment of Mediterranean oaks. Journal of Applied Ecology, 2014, 51(2): 349-358. DOI:10.1111/1365-2664.12189 |
| [24] |
张春雨, 赵秀海, 赵亚洲. 长白山温带森林不同演替阶段群落结构特征. 植物生态学报, 2009, 33(6): 1090-1100. DOI:10.3773/j.issn.1005-264x.2009.06.009 |
| [25] |
Cao J, Zhang C Y, Zhao B, Li X Y, Hou M M, Zhao X H. Seedling density dependence regulated by population density and habitat filtering:evidence from a mixed primary broad-leaved Korean pine forest in Northeastern China. Annals of Forest Science, 2018, 75(1): 25. DOI:10.1007/s13595-018-0706-x |
| [26] |
Duncan R S, Chapman C A. Tree-shrub interactions during early secondary forest succession in Uganda. Restoration Ecology, 2003, 11(2): 198-207. DOI:10.1046/j.1526-100X.2003.00153.x |
| [27] |
刘双, 金光泽. 小兴安岭阔叶红松(Pinus koraiensis)林种子雨的时空动态. 生态学报, 2008, 28(11): 5731-5740. DOI:10.3321/j.issn:1000-0933.2008.11.060 |
| [28] |
Denslow J S, Uowolo A L, Hughes R F. Limitations to seedling establishment in a mesic Hawaiian forest. Oecologia, 2006, 148(1): 118-128. DOI:10.1007/s00442-005-0342-7 |
| [29] |
韩文娟, 曹旭平, 张文辉. 地被物对油松幼苗早期更新的影响. 林业科学, 2014, 50(1): 49-54. |
| [30] |
Masaki T, Nakashizuka T. Seedling demography of swida controversa:effect of light and distance to conspecifics. Ecology, 2002, 83(12): 3497-3507. DOI:10.1890/0012-9658(2002)083[3497:SDOSCE]2.0.CO;2 |
| [31] |
孟令君, 姚杰, 秦江环, 范春雨, 张春雨, 赵秀海. 吉林蛟河针阔混交林乔木幼苗组成及其密度格局影响因素. 植物生态学报, 2018, 42(6): 653-662. |
| [32] |
韩有志, 王政权, 谷加存. 林分光照空间异质性对水曲柳更新的影响. 植物生态学报, 2004, 28(4): 468-475. DOI:10.3321/j.issn:1005-264X.2004.04.004 |
| [33] |
曾德慧, 尤文忠, 范志平, 刘明国. 樟子松人工固沙林天然更新障碍因子分析. 应用生态学报, 2002, 13(3): 257-261. DOI:10.3321/j.issn:1001-9332.2002.03.001 |
| [34] |
Record S, Kobe R K, Vriesendorp C F, Finley A O. Seedling survival responses to conspecific density, soil nutrients, and irradiance vary with age in a tropical forest. Ecology, 2016, 97(9): 2406-2415. DOI:10.1002/ecy.1458 |
| [35] |
陈贝贝, 赵秀海, 倪瑞强, 黄珍. 长白山阔叶红松林乔木树种幼苗组成与空间分布. 东北林业大学学报, 2012, 40(10): 39-42. DOI:10.3969/j.issn.1000-5382.2012.10.010 |
| [36] |
Granda E, Escudero A, Valladares F. More than just drought:complexity of recruitment patterns in Mediterranean forests. Oecologia, 2014, 176(4): 997-1007. DOI:10.1007/s00442-014-3064-x |
| [37] |
Van Huong P, 陈昌雄, 张巧巧, Van Tung H, 范先明, Duy N H. 灌草对胖大海种群更新幼树幼苗密度的影响. 西南林业大学学报, 2016, 36(4): 1-8. |
| [38] |
Tsvuura Z, Lawes M J. Light availability drives tree seedling success in a subtropical coastal dune forest in South Africa. South African Journal of Botany, 2016, 104: 91-97. DOI:10.1016/j.sajb.2015.11.009 |