 2020, Vol. 40
2020, Vol. 40文章信息
- 王佳敏, 宋海燕, 陈金艺, 张静, 李素慧, 陶建平, 刘锦春
- WANG Jiamin, SONG Haiyan, CHEN Jinyi, ZHANG Jing, LI Suhui, TAO Jianping, LIU Jinchun
- 多年生黑麦草对干旱胁迫下喀斯特异质生境的生长响应策略
- Response strategies of Lolium perenne L. to karst heterogeneous habitats under drought stress
- 生态学报. 2020, 40(13): 4566-4572
- Acta Ecologica Sinica. 2020, 40(13): 4566-4572
- http://dx.doi.org/10.5846/stxb201911162471
-
文章历史
- 收稿日期: 2019-11-16
- 修订日期: 2020-04-28

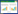




由于特殊的地质环境背景和长期的岩溶作用等, 喀斯特地区地形破碎, 基岩裸露, 岩石与少量土壤呈镶嵌分布, 植物生长的小生境具有强烈的异质性[1-2]。如在基岩裸露突出的地方形成石土面、土坑小生境[3], 其土层十分浅薄, 极易造成岩溶干旱和养分缺失, 且植物根系的纵向拓展空间十分有限[4], 而在岩体周围形成落水洞、漏斗、石缝等小生境, 土层分布较厚, 该生境单位时间蒸发量较少, 保水能力相对较强, 养分相对充足, 且为根系下扎提供充足的空间, 有利于植物的生长[2], 但由于喀斯特黄色石灰土的土壤透气性和通气性差、易板结, 深土区也可能造成水分胁迫, 不利于植物根系的呼吸和营养的获取[4]。另一方面, 随着大气CO2浓度的升高和气温持续上升, 全球背景下的降雨格局正在发生变化, 中高纬度和热带地区的降雨量会增加, 而我国喀斯特所处的亚热带地区的降雨量会减少[5]。喀斯特脆弱的生态系统对于外界环境具有高度的依赖性和敏感性[6], 降雨格局的变化可能会加重“岩溶干旱”发生的频率和强度, 也将进一步加重该地区土层空间的异质性, 从而对植物的生存和发展产生更大的影响[7-8]。因此, 喀斯特地区的植物往往面临着非常严峻的生存考验:在浅土区挨饿受渴?还是在深土区耗费体力下扎觅食?亦或通过自身的智慧充分利用浅土与深土的“资源”, 提高对这种严酷异质生境的适应?
在长期进化过程中, 喀斯特地区植物通常也会形成一系列适应异质性生境的生长和生理策略[9]。如林艳华等[10]的研究发现复羽叶栾树幼苗可以通过提高水分利用效率和降低对根系资源的投资来提高净光合速率以适应喀斯特水分异质生境。黄小辉等[11]的研究表明, 桑树可以通过向水性和向养性运动适应喀斯特地区的异质生境, 但是向养性必须是在水分充足条件下发生, 如果水分缺乏, 即使周边有高养分斑块, 桑树也不会主动向高养斑块生长。
目前研究极大地关注了喀斯特地区植物对岩溶干旱的响应[12], 也有研究逐渐关注该地区土层厚度对植物的影响[2], 但很少关注该地区异质性生境对植物的影响, 尤其是综合考虑土壤水分含量、植物生存空间和土壤透气性等状况的共同影响。因此, 本研究以多年生黑麦草为研究材料, 通过模拟同质和异质生境, 探究异质生境下植物是否会主动寻求高水分、厚土壤、或者更宽阔的生长空间来提高其对异质性生境的适应。
1 材料与方法 1.1 实验材料供试土壤为重庆市具有喀斯特地貌的中梁山黄色石灰土。土壤基本理化性状为:pH为7.4±0.14, 有机质为(0.34±0.02)%, 全氮为(0.28±0.03)g/kg, 全磷为(0.39±0.02)g/kg, 全钾为(23.7±3.22)g/kg, 田间持水量为(39.8±2.23)%, 土壤容重约为0.013 g/cm3。
对于水分及其他营养物质要求较高的植物难以在喀斯特地区生存, 相反对于耐旱、喜钙且善于在异质性石沟、石缝中汲取营养的植物更适合作为喀斯特地区植被恢复的属种[13]。黑麦草(Lolium perenne L.)是禾本科多年生草本, 由于其根系可塑性高, 适应性较强, 可以用于喀斯特地区土壤改良和水土保持[14], 因此本研究选取其为实验材料。
1.2 实验方法本研究采用土壤深度和水分处理的双因素随机区组试验, 设置3种降水量处理和两种生境处理。生境处理:将异质生境S-D作为实验组, 通过组合深土容器D(长×宽×高:5 cm×10 cm×50 cm)和浅土容器S(长×宽×高:50 cm×10 cm×5 cm)来实现(图 1), 将深土容器置于事先挖好的沟槽中, 使其开口和浅土容器的开口处于同一水平面上, 并用聚酯捆扎带固定组合, 用于模拟喀斯特包括石沟、石缝、石土面等的异质性小生境;保持土壤总体积与异质生境的总体积相同, 将中部设有挡板的同质容器S-S(长×宽×高:100 cm×10 cm×5 cm)作为对照组, 用于模拟喀斯特单一石土面小生境。

|
| 图 1 生境设置 Fig. 1 habitats treatments S表示浅土容器;D表示深土容器 |
水分处理根据重庆市近20年来气象局统计的降雨频率和降雨量计算得来, 降雨频率设置为每3天一次, 降雨量设置3个梯度:对照W0(正常降水)、中度干旱W1(减水50%)、重度干旱W2(减水70%)(表 1)。
| 施水量处理 Treatment of irrigation |
干旱程度 Degree of drought/% |
占降雨量百分比 Precipitation percentage |
同质生境 uniform habitat /(mL/3d) |
异质生境S区 region-S of heterogeneous habitat/(mL/3d) |
异质生境D区 region-D of heterogeneous habitat/(mL/3d) |
| W0 | 对照 | 100 | 1600 | 860 | 90 |
| W1 | 中度干旱 | 50 | 860 | 430 | 45 |
| W2 | 重度干旱 | 30 | 520 | 260 | 30 |
| W0:对照水分Control group;W1:中度干旱Moderate drought;W2:重度干旱Severe drought | |||||
2018年2月6日开始育苗, 5月5日选取生长状态良好、一致的幼苗进行移栽, 异质生境S-D处理需将黑麦草根系均匀的分布在两个规格的容器中, 同质生境S-S处理需将黑麦草根系均匀分布在挡板两侧, 5月25日开始进行灌溉处理, 实验共设7个重复, 处理60天。
1.3 指标测定对地上部分和地下部分进行收获后, 使用数字化扫描仪(STD1600Epson USA)扫描植物仔细清洗过的根系, 用win.Rhzo (Version410B)根系分析系统软件(Refent Instmment Inc, Canada)对植物根系表面积、根系长度、根体积等进行测量, 扫描后将根系及地上部分全部于80℃烘干至恒重, 称取各个分区及地上部分的干重。
1.4 数据统计数据分析采用SPSS 22.0, 用单因素方差分析(One-way ANOVA)方法, 分析相同水分条件下株高、分蘖数、生物量、地上质量分数和根质量分数在同质和异质生境的差异;同时, 用配对样本T检验的方法, 分析异质生境中相同水分条件下生物量、生物量占比、比根长、比根面积和根组织密度等指标在浅土和深土区的差异;用多重比较(LSD)分析生物量、生物量占比、比根长、比根面积和根组织密度在不同水分条件下的差异。本研究使用Origin 8.6软件进行作图。
2 研究结果 2.1 株高和分蘖数在对照和中度干旱条件下, 异质和同质生境中黑麦草的株高和分蘖数均没有显著性差异, 但在重度干旱条件下, 异质性生境中的黑麦草的株高和分蘖数均显著大于同质生境:相较于同质性生境, 异质生境中黑麦草的株高极显著增加(P < 0.01), 其增加幅度为37.86%;分蘖数极显著增加(P < 0.01), 其增加幅度为122.22%(图 2)。

|
| 图 2 不同水分处理下异质和同质生境对黑麦草株高及分蘖数的影响(平均值±标准误) Fig. 2 Effect of different water treatments on biomass height and tiller number of L.perenne in heterogeneous and uniform habitats(M±SE) **极显著水平(P < 0.01), ns未达到显著水平(P>0.05);S-S:同质生境Shallow-shallow soil;S-D:异质生境Shallow-deep soil;W0:对照水分Control group;W1:中度干旱Moderate drought;W2:重度干旱Severe drought |
在对照和中度干旱条件下, 异质生境和同质生境中黑麦草地上、地下部分的生物量和总生物量均无显著性差异;在重度干旱条件下, 异质生境处理下的地上、地下和总生物量均显著大于同质生境(P < 0.05):相较于同质生境, 重度干旱条件下异质生境的地上生物量极显著增加(P < 0.01), 其增加幅度为129.21%, 地下部分生物量显著增加了65.72%(P < 0.05), 总生物量显著增加了107.44%(P < 0.05)(图 3)。

|
| 图 3 不同水分处理下异质和同质生境对黑麦草生物量积累及分配的影响(平均值±标准误) Fig. 3 Effect of different water treatments on biomass accumulation and distribution of L. perenne in heterogeneous and uniform habitats(Mean ± SE) **极显著水平(P < 0.01), *显著水平(P < 0.05), ns未达到显著水平(P>0.05) |
在对照和中度干旱条件下, 异质生境和同质生境中黑麦草的地上质量分数和根质量分数无显著差异;重度干旱条件下, 异质生境的地上质量分数较同质生境显著增加了13.04%(P < 0.05), 根质量分数较同质生境显著减小了23.64%(P < 0.05)(图 3)。
2.3 分区根系生长黑麦草的根系生物量在3种水分条件下, 同质生境处理的挡板两测均无显著性差异, 均保持在整个根系的50%左右;但异质生境深土区生物量均显著高于浅土区生物量(P<0.05)。随着干旱程度的加剧, 异质性生境处理的黑麦草在浅土区的根系生物量呈逐渐减小的趋势, 在深土区的生物量呈先增大后减小的趋势;浅土区总根系生物量之比呈先减小后增加的趋势, 分别占整个植株的17.92%(W0)、12.28%(W1)、15.60%(W2), 而深土区占总根系生物量之比呈先增加后减小的趋势(图 4)。

|
| 图 4 不同水分处理下同质生境和异质生境中黑麦草分区根系生物量(平均值±标准误) Fig. 4 Effect of different water treatments on root biomass accumulation and distribution of L. perenne.in heterogeneous and uniform habitats(M±SE) 不同小写字母表示异质生境中浅土区在不同水分条件下的差异显著性(P < 0.05);不同大写字母表示异质生境中深土区在不同水分条件下的差异显著性(P < 0.05) |
同质生境处理中, 挡板左右两侧的比根长、比根面积、根组织密度在3种水分条件下均无显著性差异。异质生境处理中, 浅土区的比根长和比根面积在对照和中度干旱条件下均显著大于深土区(P<0.05), 随着施水量的减少, 浅土区的比根长和比根面积无明显变化, 而深土区逐渐增大, 浅土区和深土区的差异逐渐减小。浅土区的根组织密度在对照和中度干旱条件下均显著小于深土区(P<0.05), 随着干旱程度的加剧, 深土区的根组织密度逐渐减小, 且与浅土区的差异逐渐减小(图 5)。

|
| 图 5 不同水分处理下同质生境和异质生境中分区黑麦草比根长、比根面积及根组织密度(平均值±标准误) Fig. 5 Effect of different water treatments on biomass specific root length、specific root area and root tissue density of L.perenne in heterogeneous and uniform habitats(M±SE) 不同小写字母表示异质生境中浅土区在不同水分条件下的差异显著性(P < 0.05);不同大写字母表示异质生境中深土区在不同水分条件下的差异显著性(P < 0.05) |
干旱胁迫往往对植物的生物量积累、株高和分蘖数产生很大影响[15]。本研究表明, 与同质生境相比, 异质生境可以缓解降水量减少对黑麦草生物量积累、株高和分蘖数的抑制作用。在土层较浅的同质生境中, 植物根系均遭受干旱和下扎空间少的双重胁迫, 植物在应对这两种胁迫时, 往往引发脱落酸、茉莉酸等的含量上升、生长素的含量减小[16], 这会促使气孔关闭, 光合速率减慢[17], 导致植物生长速率减慢, 减少地上、地下和总生物量的积累[18], 这虽然可以减弱水分不足对黑麦草生理活动造成的压力、一定程度地抵抗水淹和干旱胁迫、维持植物的基本生长, 但这也抑制了黑麦草株高的增长[19], 抑制分蘖芽的形成和生长, 延迟分蘖发育[20];而在异质生境中, 深土区的土壤蓄水能力较强, 土壤含水量较高, 可能会促使深土区的根系产生生长素[16], 一定程度上增大叶片含水量和植株的光合速率, 使异质生境中的植物生物量和株高相对增加, 分蘖数相应增多, 表现为一定程度上缓解植物的干旱胁迫。
植物干物质分配被认为是植物适应逆境胁迫的重要策略之一[21], 许多研究表明, 通过调节地上、地下生物量分配可以改善自身对干旱的适应性[22], 本研究中黑麦草表现为类似的规律, 在降水量较小时, 同质性生境更能促进根系生物量的投入、增加根质量分数;而异质生境会相应的减小根系生物量的投入, 减小根质量分数, 这可能是因为异质生境中, 深土区的存在使根系生长在较为湿润的土壤环境中, 根系多余的增生没有相应的水分吸收回报, 所以可能会通过减少根系生物量的方式, 进而减少根系呼吸作用带来的能量消耗[23]。
在异质性生境中, 根系在不同的土壤条件下往往会采取不同的生物量分配和形态应对策略, 使植株获得更多的水分和养分[24]。本研究表明, 异质生境中, 黑麦草倾向于将更多的根系生物量分配到深土区, 而将较少的生物量分配到浅土区, 这与赵娣等[25]对于番茄分根灌溉的结果一致, 可能与植物根系具有觅食反应有关[26]:处于异质性环境中的植物根系将会在水分和养分充足的区域增生, 以获得更多的资源[27]。随着干旱程度的加重, 异质性生境中深土区和浅土区的水分差异更大, 在维持浅土区正常生长的基础上, 黑麦草会尽可能地减少浅土区的生物量, 同时增加深土区的生物量, 但是这两个区域生物量投入的差异是有限度的, 在重度干旱的情况下, 浅土区水分无法维持浅土区根系的生长, 深土区有可能将水分逆运输至浅土区从而维持浅土区根系的正常生长[28], 故而表现为在重度干旱的情况下, 浅土区占整个根系生物量之比又有升高趋势。
深土区在增大生物量投入的同时增大了根组织密度, 增强根系延伸能力, 帮助根系在深土区克服阻力下扎, 使植株获得大量的水分和养分;而浅土区在减小生物量投入的同时, 充分利用较少的资源和生物量, 通过减小根组织密度、增大比根长和比根面积的方式迅速发展根系, 增强根系吸水能力, 提高水分吸收速率和和养分吸收速率, 加强浅土区水分的吸收和利用。随着干旱程度的加深, 植物往往会增大其比根长和比根面积来适应水分的减少[29-30], 本研究表明:当异质性生境中黑麦草面临干旱胁迫时, 由于浅土区比根长、比根面积增加到达了一定限度, 黑麦草会通过逐渐增大深土区的比根长和比根面积、减小根组织密度来应对干旱胁迫。
致谢: 感谢西南大学地理科学学院项欣奕老师对写作的帮助。| [1] |
李周, 赵雅洁, 宋海燕, 张静, 陶建平, 刘锦春. 喀斯特土层厚度异质性对草地群落结构和优势种生长的影响. 草业科学, 2017, 34(10): 2023-2032. DOI:10.11829/j.issn.1001-0629.2017-0296 |
| [2] |
赵雅洁, 张静, 宋海燕, 李周, 李素慧, 陶建平, 刘锦春. 不同土壤厚度、水分和种植方式对喀斯特两种草本凋落物分解质量损失和化学计量特征的影响. 生态学报, 2018, 38(18): 6549-6558. |
| [3] |
廖洪凯, 龙健, 李娟, 杨江江, 冯业强. 喀斯特地区不同植被下小生境土壤矿物组成及有机碳含量空间异质性初步研究. 中国岩溶, 2010, 29(4): 434-439. DOI:10.3969/j.issn.1001-4810.2010.04.014 |
| [4] |
Zhang J, Wang J M, Chen J Y, Song H Y, Li S H, Zhao Y J, Tao J P, Liu J C. Soil Moisture Determines Horizontal and Vertical Root Extension in the Perennial Grass Lolium perenne L. growing in karst soil. Frontiers in Plant Science, 2019, 10: 629. |
| [5] |
张琪, 李跃清. 近48年西南地区降水量和雨日的气候变化特征. 高原气象, 2014, 33(2): 372-383. |
| [6] |
陆清平, 赵宇鸾, 葛玉娟. 基于栅格的喀斯特山区土地系统脆弱性评价:以贵州普定县为例. 环境科学与技术, 2019, 42(9): 221-229. |
| [7] |
杨宇琼, 戴全厚, 李昌兰, 彭旭东, 严友进. 模拟降雨条件下喀斯特坡耕地氮磷元素地下漏失特征. 中国水土保持科学, 2018, 16(3): 59-67. |
| [8] |
贺晋云, 张明军, 王鹏, 王圣杰, 王兴梅. 近50年西南地区极端干旱气候变化特征. 地理学报, 2011, 66(9): 1179-1190. |
| [9] |
盘邹.喀斯特生态系统的生境异质性与植物适应性[D].桂林: 广西师范大学, 2006.
|
| [10] |
林艳华, 梁千慧, 刘锦春. 喀斯特地区适生树种复羽叶栾树幼苗对干旱胁迫下异质生境的生长和光合响应. 西南大学学报:自然科学版, 2019, 41(8): 20-26. |
| [11] |
黄小辉, 冯大兰, 刘芸, 朱恒星, 陈道静, 耿养会. 模拟石漠化异质生境中桑树的生长和叶绿素荧光特性. 北京林业大学学报, 2016, 38(10): 50-58. |
| [12] |
张中峰, 黄玉清, 尤业明, 何成新, 朱访, 黎彦余. 岩溶干旱胁迫下青冈栎水分参数变化. 生态学杂志, 2011, 30(4): 664-669. |
| [13] |
杨明德. 论喀斯特环境的脆弱性. 云南地理环境研究, 1990, 1(1): 21-29. |
| [14] |
莫正海. 黑麦草的饲用价值与生态作用研究. 现代农业科技, 2010(6): 332-333. |
| [15] |
赵雅洁, 李周, 宋海燕, 张静, 梁千慧, 刘锦春. 喀斯特地区土壤厚度降低和水分减少对两种草本植物混种后光合的影响. 草业科学, 2017, 34(7): 1475-1486. |
| [16] |
郭宾会, 戴毅, 宋丽. 干旱下植物激素影响作物根系发育的研究进展. 生物技术通报, 2018, 34(7): 48-56. |
| [17] |
陈晶.生长素类物质对镉胁迫下玉米幼苗生长及生理特性的影响[D].重庆: 西南大学, 2016.
|
| [18] |
张静懿.生长素调控气孔发育的功能和作用分子机理的研究[D].上海: 上海交通大学, 2015.
|
| [19] |
周勇, 张晓宏, 米日古丽·马木提, 马盾, 周小云. 陆地棉半矮秆突变体sd株高对外源激素的敏感性. 新疆农业科学, 2018, 55(3): 405-413. |
| [20] |
王如芳, 张吉旺, 吕鹏, 董树亭, 刘鹏, 赵斌. 不同类型玉米品种分蘖发生过程中内源激素的作用. 中国农业科学, 2012, 45(5): 840-847. DOI:10.3864/j.issn.0578-1752.2012.05.003 |
| [21] |
赵俊芳, 杨晓光, 陈斌, 王化琪, 王志敏, Bouman B A M. 不同灌溉处理对旱稻根系生长及水分利用效率的影响. 中国农业气象, 2004, 25(4): 44-48. DOI:10.3969/j.issn.1000-6362.2004.04.011 |
| [22] |
Markesteijn L. Drought Tolerance of Tropical Tree Species; Functional Traits, Trade-Offs and Species Distribution[D]. the Netherlands: Wageningen University, 2010.
|
| [23] |
武荣, 李援农. 水氮耦合对冬小麦根系分布和根冠比及产量的影响. 南方农业学报, 2013, 44(6): 963-967. DOI:10.3969/j:issn.2095-1191.2013.6.963 |
| [24] |
马闯, 张文辉, 吴敏, 马莉薇, 周建云, 薛瑶芹. 栓皮栎幼苗根系发育对环境异质性的响应. 林业科学, 2013, 49(10): 58-65. |
| [25] |
赵娣, 王振华, 李文昊, 张金珠, 吕德生. 分根区交替灌溉对膜下滴灌加工番茄根系特征及产量的影响. 核农学报, 2018, 32(10): 2069-2079. DOI:10.11869/j.issn.100-8551.2018.10.2069 |
| [26] |
Einsmann J C, Jones R H, Pu M, Mitchell A J. Nutrient foraging traits in 10 co-occurring plant species of contrasting life forms. Journal of Ecology, 1999, 87(4): 609-619. |
| [27] |
李洪波.植物响应养分空间异质性分布的动态过程及调控根系获取养分的策略研究[D].北京: 中国农业大学, 2014.
|
| [28] |
Breazeale J F. Maintenance of Moisture-Equilibrium and Nutrition of Plants at and Below the Wilting Percentage. Arizona: College of Agriculture, University of Arizona, 1930.
|
| [29] |
白国华. 不同干旱胁迫对榆树幼苗根叶形态的影响. 防护林科技, 2016(10): 41-42, 49-49. |
| [30] |
黄小辉.桑树对干旱生境的响应及其适应性[D].重庆: 西南大学, 2014.
|